
3种珍贵树种幼苗光合特性及日进程研究
2022-07-25 08:10王立冬陈艳艳汤行昊张丽琼梁海燕闫小莉
山地农业生物学报 2022年4期
王立冬,陈艳艳,汤行昊,张丽琼,梁海燕,闫小莉*
(1.福建农林大学 林学院,福建 福州 350002;2.福建省林业科学研究院,福建 福州 350012)
壳斗科 (Fagaceae)植物广泛分布于温带、热带、亚热带森林,是针阔混交林和常绿阔叶林的重要组成成分。壳斗科植物可提供木材,还可作能源植物和木本粮食,具有巨大的开发潜力。大叶青冈((Hand,-Mzt.)Cheng et T.Hong)材质坚重,强度大,收缩性大且耐腐蚀,可用于制作各类木制器械。赤皮青冈((Blume)Oerst)材质硬度高且纹理漂亮,是高档用材树种;同时,赤皮青冈树形高大且为深根性树种,在涵养水源和保持水土方面效果显著,是兼具园林观赏、生态绿化与工业用材等多功能于一体的珍贵乡土树种。红椎(Miq.)是华南地区重要的珍贵乡土造林树种,速生,生态适应性广。大叶青冈、赤皮青冈和红椎是南方地区针阔混交林营造中被广泛应用的阔叶树种,但是,由于长期以来对这3个壳斗科珍贵树种的过量砍伐利用,使其资源受到严重破坏甚至枯竭,已被列为中国Ⅱ级珍稀渐危树种。目前,国内外针对3个珍贵树种的研究多在生理生态、生长发育和养分吸收等方面,如Zhao等研究了不同坡向对红锥人工林土壤种子库数量、质量和幼苗萌发过程的影响;Lian等利用16S rRNA基因高通量测序和PICRUSt预测技术研究了红椎人工林不同土层的土壤细菌群落结构和功能;Xu等研究了青冈的抗冻和抗机械损伤能力,并采用比较蛋白质组学分析方法对青冈发芽率低的原因进行了研究;施肥及不同施肥量对赤皮青冈生长的影响等。总体来说,针对这3个珍贵树种的光合生理特性的相关研究相对较少,特别是针对大叶青冈、赤皮青冈和红椎苗木光合特性和日进程及其与环境因子的关系的研究十分匮乏,仅有吴丽君等研究了不同胁迫条件对赤皮青冈的生长和光合生理的影响。
光合作用是植物体内极为重要的代谢过程,植物通过光合作用合成有机物为自身生长发育提供物质和能量基础。受遗传因子控制,不同植物具有不同的光合生理特性,植物叶片的净光合速率(Pn)、气孔导度(Gs)、蒸腾速率(Tr)、胞间CO浓度(Ci)和水分利用效率(WUE)等主要光合相关生理特性因子综合反映了植物的生长发育、物质生产与水分消耗关系。Pn是反映植物光合能力最直观的光合指标,Pn越大,植物在单位时间内积累的有机物越多,在适宜环境条件下,Pn与叶片叶绿素含量有直接关系,在一定范围内,植物叶片叶绿素含量越高,Pn越大。植物通过调节叶片气孔开合程度来控制体内水分流失和与外界CO气体的交换,气孔导度是影响植物蒸腾作用、呼吸作用以及光合作用的主要因素之一,同时植物胞间CO浓度的高低也与气孔导度有关。光合作用生理过程复杂,不仅与植物自身遗传因子密切相关,还受光照强度、空气温度、空气湿度、空气CO浓度和土壤含水量等外界环境因子影响。
鉴于此,本研究以3个珍贵树种大叶青冈、赤皮青冈和红椎幼苗为研究对象,对3个树种的光合生理特性进行测定,分析比较3个树种的光合日变化和光合日均值,以及与其他光合参数和环境因子间的相关性,以期初步明确3个珍贵壳斗科树种的光合特性及其对环境的适应性差异,从而为3个树种的培育、树种保护和混交造林树种选择提供科学依据。
1 材料与方法
1.1 试验地概况
试验地点设置在福建省林业科学研究院的试验苗圃,地理位置为E 119°30′,N 26°15′,海拔50 m,温暖湿润,阳光充足,雨量充沛,无霜期326 d,属于典型的亚热带季风气候。年平均日照时数为1700~1980 h,年平均降水量为900~2100 mm,年平均气温为20~25 ℃。试验地2021年4月20日日出时刻为5∶34′39″,日落时刻为18∶28′41″,至2021年5月5日日出时刻提前至5∶22′28″,日落时刻推迟至18∶36′28″。
1.2 试验材料
为模拟自然状态下林木幼苗的生长环境,本试验在幼苗种植阶段统一用50%透光率的遮阳网对3个树种幼苗进行遮荫处理,试验选取生长状况良好且长势一致的两年生大叶青冈、赤皮青冈、红椎幼苗各6株作为试验材料进行研究,幼苗苗高采用卷尺进行测量,地径采用游标卡尺进行测量,利用Epson Expression 12000XL扫描仪进行叶片扫描,并用WinFOLIA分析软件对幼苗叶片叶长、叶宽和叶面积进行分析,幼苗植株生长状况和叶片基本数据见表1。
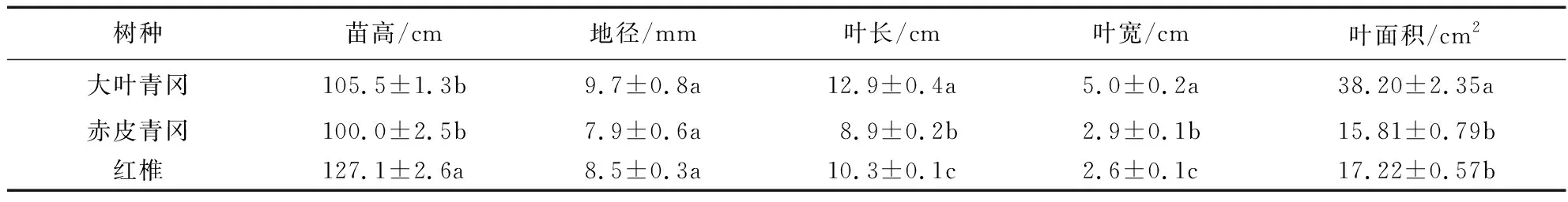
表1 3个壳斗科树种幼苗植株生长状况和叶片基本数据Tab.1 Seedling growth status and leaf basic data of three Fagaceae Species
1.3 试验方法
..光合参数测量
2021年4月下旬至5月上旬选择3天晴朗无云的天气,使用便携式光合仪(LI-6800,LI-COR Biosciences,Lincoln,NE,USA)进行3个树种幼苗的叶片光合参数及其日变化测定,测定时将3×3 cm红蓝光源叶室Light值(光子通量)统一设置为1000 μmol/(m·s),以确保研究树种幼苗叶片获得50%遮荫环境下的最大光子通量,叶室温度设置在30 ℃,CO浓度设置在400 μmol/(m·s),湿度控制在50%~75%。光合作用速率从8∶00到16∶00每间隔2 h测定一次叶片的净光合速率Pn μmol/(m·s)、气孔导度Gs mol/(m·s)、蒸腾速率Tr mol/(m·s)、胞间CO浓度Ci μmol/(m·s)等光合生理指标,以及空气相对湿度(RH)、空气温度(Ta)、空气CO浓度(Ca)和饱和水汽压亏缺(Vpdl)等环境因子,并计算3个树种叶片水分利用效率(WUE)=Pn/Tr。测定时,每个树种选择3棵长势一致的植株,每一个植株在中部向阳的相同高度位置选择3片无病斑损伤的健康成熟叶片进行测定,每片叶片重复测量5次。
..叶绿素含量测定
每株幼苗在树冠中部相同高度位置收集2 g(约4~5片)无病斑损伤的健康成熟叶片,采用丙酮—乙醇(1∶1)浸提法提取叶绿素,在分光光度计下测量645 nm和663 nm波长处的吸光度,用修正过的Arnon公式计算叶绿素含量。
..数据分析
试验数据采用Excel 2019进行处理,采用SPSS 24.0软件进行单因素方差分析和皮尔森相关性分析,用LSD法检验=0.05水平上的差异显著性,采用Origin 2019软件进行绘图。
2 结果与分析
2.1 3个壳斗科树种光合特性日变化
..净光合速率Pn和气孔导度Gs的日变化
图1-a可见,大叶青冈净光合速率Pn日变化呈先降后升的“U”字型,赤皮青冈Pn日变化呈“降—升—降”的倒“S”字型,红椎Pn 8∶00至10∶00下降明显,10∶00之后无显著升降变化。3个树种Pn均在接近正午或正午前逐渐下降且正午之后上升,说明均存在“光合午休”现象。大叶青冈和赤皮青冈Pn最低值出现在12∶00,分别为0.177 μmol/(m·s)和0.991 μmol/(m·s);红椎Pn最低值出现在10∶00,为0.400 μmol/(m·s),大叶青冈和赤皮青冈Pn从8∶00至12∶00分别下降2.21 μmol/(m·s)和0.63 μmol/(m·s),红椎Pn从8∶00至10∶00下降 1.01 μmol/(m·s),大叶青冈Pn降幅最大,赤皮青冈Pn降幅最小。图1-b可知,大叶青冈Gs日变化呈“W”字型,赤皮青冈和红椎Gs日变化均呈“降—升—降”的倒“S”型,大叶青冈Gs最低值出现在14∶00为33.7 mmol/(m·s),赤皮青冈和红椎Gs最低值出现在10∶00分别为18.0 mmol/(m·s)和 14.3 mmol/(m·s)。

图1 3个壳斗科树种叶片净光合速率Pn和气孔导度Gs的日变化Fig.1 Diurnal changes of net photosynthetic rate and stomatal conductance in leaves of three Fagaceae species
..蒸腾速率Tr和胞间CO浓度Ci的日变化
图2-a可见,大叶青冈的蒸腾速率(Tr)8∶00至10∶00下降幅度较大,10∶00之后无显著升降变化,赤皮青冈和红椎Tr日变化均呈“降—升—降”

图2 3个壳斗科树种叶片蒸腾速率Tr和胞间CO2浓度Ci的日变化Fig.2 Diurnal changes of transpiration rate and intercellular carbon dioxide concentration in leaves of three Fagaceae species
的倒“S”型,3个树种均在10∶00出现谷值分别为1431.8 μmol/(m·s)、797.2 μmol/(m·s)和535.6 μmol/(m·s)。 图2-b可见,大叶青冈胞间CO浓度Ci日变化整体呈先上升后下降的趋势,赤皮青冈Ci日变化呈“降—升—降—升”的波状,红椎Ci日变化呈“升—降—升”的倒“Z”型,大叶青冈和红椎Ci最大值出现在10∶00,分别为310.9 μmol/mol和290.6 μmol/mol,赤皮青冈Ci最大值出现在12∶00为284.9 μmol/mol。
..水分利用效率WUE的日变化
由图3可见,大叶青冈、赤皮青冈和红椎水分利用效率(WUE)日变化均呈先下降后上升的趋势,且均在12∶00出现最低值,分别为889.0 μmol/(m·s)、1061.9 μmol/(m·s)和1137.3 μmol/(m·s)。大叶青冈、赤皮青冈和红椎WUE在8∶00-12∶00分别下降56.1%、30.5%和49.8%。

图3 3个壳斗科树种叶片水分利用效率WUE的日变化Fig.3 Diurnal variation of water use efficiency in leaves of three Fagaceae species
2.2 3个壳斗科树种的光合参数日均值差异分析
..净光合速率Pn日均值和气孔导度Gs日均值差异分析
图4-a可见,3个树种Pn日均值差异显著(<0.05),依次为大叶青冈>赤皮青冈>红椎。大叶青冈、赤皮青冈和红椎Pn日均值分别为1.86 μmol/(m·s)、1.50 μmol/(m·s)和0.80 μmol/(m·s),大叶青冈Pn日均值分别比赤皮青冈和红椎高23.8%和133.3%。图4-b可见,3个树种Gs日均值差异显著(<0.05),与Pn均值对应,呈相同顺序规律,依次为大叶青冈>赤皮青冈>红椎。大叶青冈、赤皮青冈和红椎Gs日均值分别为40.9 mmol/(m·s)和25.7 mmol/(m·s)和18.7 mmol/(m·s),大叶青冈Gs日均值分别比赤皮青冈和红椎高59.0%和118.3%。

注:图中不同小写字母代表不同树种间差异显著(P<0.05),下同。图4 3个壳斗科树种叶片净光合速率Pn、气孔导度Gs、蒸腾速率Tr和胞间CO2浓度Ci的日均值Fig.4 Daily average values of net photosynthetic rate,stomatal conductance,transpiration rate and intercellular carbon dioxide concentration in leaves of three Fagaceae species
..蒸腾速率Tr日均值和胞间CO浓度Ci日均值差异分析
图4-c可见,3个树种Tr日均值差异显著(<0.05),依次为大叶青冈>赤皮青冈>红椎。大叶青冈、赤皮青冈和红椎Tr日均值分别为1521.6 μmol/(m·s)、1010.4 μmol/(m·s)和669.0 μmol/(m·s),大叶青冈Tr日均值分别比赤皮青冈和红椎高50.6%和127.4%。图4-d可见,大叶青冈与赤皮青冈和红椎之间Ci日均值差异显著(<0.05),赤皮青冈与红椎之间差异不显著,3个树种Ci日均值依次为大叶青冈>红椎>赤皮青冈。大叶青冈、赤皮青冈和红椎Ci日均值分别为299.1 μmol/mol、275.8 μmol/mol和280.18 μmol/mol。大叶青冈的Ci日均值分别比赤皮青冈和红椎高8.5%和6.8%。
..水分利用效率WUE日均值差异分析
图5可见,大叶青冈与赤皮青冈之间及大叶青冈与红椎之间WUE日均值差异显著(<0.05),赤皮青冈与红椎之间差异不显著。3个树种WUE日均值顺序为赤皮青冈>红椎>大叶青冈,赤皮青冈WUE日均值最高,为1538.4 μmol/(m·s),分别比红椎和大叶青冈高1.5%和18.5%。

图5 3个壳斗科树种叶片水分利用效率WUE的日均值Fig.5 Daily average value of water use efficiency in leaves of three Fagaceae species
2.3 叶绿素含量
由表2可见,3个树种叶片的叶绿素含量存在显著差异(<0.05),叶绿素a含量、叶绿素b含量和总叶绿素含量的高低顺序均为:大叶青冈>赤皮青冈>红椎。大叶青冈叶片的叶绿素a含量分别比赤皮青冈和红椎高4.6%和44.7%,叶绿素b含量分别比赤皮青冈和红椎高0.3%和56.3%,总叶绿素含量分别比赤皮青冈和红椎高3.3%和47.8%。3个树种叶片的叶绿素a含量均大于叶绿素b含量。3个树种叶片叶绿素a与叶绿素b的比值分别为2.5、2.4和2.7。

表2 3个壳斗科树种幼苗叶片的叶绿素含量Tab.2 Chlorophyll content in seedling leaves of three Fagaceae Species
2.4 相关性分析
..净光合速率和蒸腾速率与其他光合指标的相关性
3个树种Pn和Tr与其他光合指标的相关性如表3,大叶青冈和赤皮青冈Pn与Tr呈极显著正相关,红椎Pn与Tr呈显著正相关;大叶青冈和赤皮青冈Pn与Gs呈极显著正相关,红椎Pn与Gs呈显著正相关;3个树种Pn均与Ci呈极显著负相关;3个树种Pn均与WUE呈极显著正相关。3个树种Tr均与Gs呈极显著正相关;大叶青冈和红椎Tr与Ci呈极显著正相关,赤皮青冈Tr与Ci呈不显著正相关;大叶青冈和红椎Tr与WUE呈极显著负相关,赤皮青冈Tr与WUE相关性不显著负相关。

表3 3个壳斗科树种叶片光合参数间的相关性Tab.3 Correlation between photosynthetic parameters in leaves of three Fagaceae species
..净光合速率和蒸腾速率与环境因子的相关性
3个树种Pn和Tr与环境因子的相关性如表4,大叶青冈和红椎Pn与Ta呈极显著负相关,赤皮青冈Pn与Ta呈不显著正相关;3个树种Pn均与RH呈极显著负相关;大叶青冈和红椎Pn与Ca呈不显著正相关,赤皮青冈Pn与Ca呈极显著负相关。大叶青冈Tr与Ta呈显著负相关,赤皮青冈Tr与Ta呈显著正相关,红椎Tr与Ta呈不显著负相关;大叶青冈和红椎Tr与Vpdl呈极显著负相关,赤皮青冈Tr与Vpdl呈不显著负相关。表5可知,3个树种Pn均与Ls呈极显著正相关,Ci均与Ls呈极显著负相关,Ls减小,胞间CO浓度上升,净光合速率下降。说明引起3个树种“光合午休”的主要因素为非气孔限制因素。

表4 3个壳斗科树种叶片光合参数与环境因子间的相关性Tab.4 Correlation between photosynthetic parameters and environmental factors in leaves of three Fagaceae species

表5 3个壳斗科树种叶片光合午休相关因子间的相关性Tab.5 Correlation among Photosynthetic lunch break related factors in leaves of three Fagaceae species
3 结论与讨论
3.1 3个树种光合特性及其日变化差异
不同树种的光合生理存在较大差异,光合作用可作为判断植物生长和抗逆性强弱的指标。植物叶片光合作用日变化的特征能够反映植物对环境的适应性,研究光合日变化是探究植物光合能力和分析环境因子影响植物生长和代谢的重要手段,大多数植物的日变化一般表现为单峰型或双峰型,有时会出现平坦型和不规则型。本试验中,大叶青冈净光合速率日变化呈先降后升的“U”字型,赤皮青冈和红椎净光合速率日变化均呈“降—升—降”的“S”型,3种植物的净光合速率日变化未呈明显的单峰型或双峰型,但净光合速率均有在正午或接近正午前下降且正午之后上升的现象,说明3个树种幼苗存在“光合午休”现象。植物“光合午休”主要表现为正午或午后叶片光合作用减弱,净光合速率降低,净光合速率的降幅反映了植物对正午强光和高温等环境的适应能力,大叶青冈、赤皮青冈和红椎Pn的降幅分别为2.21 μmol/(m·s)、0.63 μmol/(m·s)和1.01 μmol/(m·s),赤皮青冈净光合速率降幅最小,“光合午休”现象最不明显,说明赤皮青冈对强光和高温环境的适应性比大叶青冈和红椎强。3种植物的气孔导度、蒸腾速率日变化规律与净光合速率相似,均在10∶00至14∶00出现最低值或谷值,植物在中午气孔收缩,气孔导度和蒸腾速率随之下降。张斌等研究了3种不同的生境对赤皮青冈光合特性的影响,结果表明不同生境条件下,赤皮青冈的净光合速率与蒸腾速率变化趋势基本一致,且呈双峰曲线,均在10∶00和14∶00出现峰值,13∶00出现谷值,这与本试验结果有偏差,本试验测得的赤皮青冈净光合速率从8∶00开始下降,在12∶00出现最低值,在14∶00出现峰值,原因可能是张斌等以成年大树为试验材料,本试验以两年生幼苗为试验材料,植物在不同生长阶段其光合生理存在差异,也可能是种源不同,本试验地经度比张斌等的试验地高9°13′,不同种源的植物光合生理指标具有明显的地带差异性,植物在不同生境中长期适应会形成不同的生活习性和不同的光照获取策略。
净光合速率决定着植物物质积累能力的高低,在一定程度上也决定着植物生长的快慢。3个树种中大叶青冈净光合速率日均值最高,赤皮青冈次之,红椎最低,3个树种气孔导度和蒸腾速率日均值大小与净光合速率呈相同规律,大叶青冈蒸腾作用最强,不仅有更强的矿质营养运输能力,而且气孔打开程度更大,使更多的CO通过气孔进入细胞间隙供叶片进行光合作用,因此大叶青冈的光合固碳能力最强,赤皮青冈次之,红椎最弱。水分利用效率是评价作物生长适宜度的综合生理生态指标,能反映植物有效利用水资源的能力,其值越大,说明植物对水分的利用效率越高。3个树种中赤皮青冈水分利用效率最高,红椎次之,大叶青冈最低,消耗等质量的水赤皮青冈制造的干物质比大叶青冈和红椎多。植物叶片的叶绿素含量在一定程度上反映了植物的光合能力,通常来说,叶片叶绿素含量越高植物吸收光能制造有机物的能力越强。3个树种叶片在叶绿素含量上差异显著(<0.05),大叶青冈叶绿素含量最高,赤皮青冈次之,红椎最低,这与3个树种的净光合速率日均值大小规律一致。研究表明,叶绿体基因组是绿色植物细胞第二大基因组,仅次于核基因组,在结构上较为保守,但经过长期的生物进化过程,不同树种仍会形成基因结构的差异,进而形成基因表达上的差异,影响植物的叶绿体结构和叶绿素含量,最终影响植物的光合作用能力。
3.2 3个树种光合作用对环境条件的适应性
植物光合作用的能力与环境条件密切相关,空气温度、空气CO浓度和空气相对湿度等环境因子共同作用并影响着植物的光合作用和生长发育。同时,植物也可通过调节自身生理生化反应响应环境的变化。不同植物对环境条件变化的适应性不同。正午或正午之后出现的强光、高温和干旱等许多环境胁迫都能造成植物净光合速率降低,不同的植物其“光合午休”现象产生的原因各异,主要是气孔限制因素和非气孔限制因素。Ls是反映植物叶片对大气CO利用效率的大小的指标,其表达公式为:Ls=1-Ci/Ca,植物叶片胞间CO浓度与大气CO浓度共同决定气孔限制值的大小,根据Farquhar的观点,通过Ls、胞间CO浓度和净光合速率间的变化关系判断净光合速率的下降是否由气孔限制因素造成,如果Ls减小,胞间CO浓度增加,而净光合速率下降,则引起“光合午休”的原因是非气孔限制因素,反之是气孔限制因素。本研究中发现,3个树种净光合速率与气孔导度呈极显著或显著的正相关关系,但与胞间CO浓度均呈极显著负相关,3个树种净光合速率均与气孔限制值呈极显著正相关,胞间CO浓度均与气孔限制值呈极显著负相关,Ls减小,净光合速率下降,胞间CO浓度却上升,说明引起3个树种“光合午休”的主要因素为非气孔限制因素。强光和高温是造成植物“光合午休”最主要的两个生态因素,由于光合作用测量过程中全天光强值设置在1000 μmol/(m·s),无法通过光强变化判断3个树种“光合午休”的原因,只能通过Pn与Ta的变化关系判断3个树种的“光合午休”是否由高温引起。光合作用生理过程复杂,易受环境影响,对温度的变化极为敏感。研究表明,高温使植物PS Ⅱ反应中心闭合程度(V)增加,受体侧损伤,引起电子传递效率降低,进而抑制光合。大叶青冈和红椎Pn与Ta呈极显著负相关,赤皮青冈Pn与Ta呈不显著正相关,说明高温可能是引起大叶青冈和红椎“光合午休”的原因,但不是引起赤皮青冈“光合午休”的原因,引起赤皮青冈“光合午休”的原因可能是强光,过剩的光能会产生活性氧,导致脂质过氧化,细胞膜的破坏,甚至发生光氧化,破坏植物的光合机构。试验结果说明赤皮青冈对高温的适应性比大叶青冈和红椎强。
饱和水汽压亏缺是重要的生态指标,在全球变暖的大环境下,陆地表面温度升高增加了大气的饱和水汽压,但空气实际水汽压没有以相同的速度增加,饱和水汽压亏缺会随着温度的升高而增加。植物光合对饱和水汽压亏缺的响应可以直观地反映植物光合作用对空气温度和水分变化的适应能力。3个树种净光合速率与空气相对湿度均呈极显著负相关,大叶青冈和红椎净光合速率与空气温度和饱和水汽压亏缺均呈极显著负相关,赤皮青冈净光合速率与空气温度和饱和水汽压亏缺呈不显著正相关,温度升高,饱和水汽压亏缺增大,大叶青冈和红椎净光合速率均下降,而赤皮青冈净光合速率不下降,说明赤皮青冈对环境温度和水分变化的耐受性强于大叶青冈和红椎。大叶青冈和红椎的蒸腾速率与空气温度呈显著负相关,赤皮青冈的蒸腾速率与空气温度呈显著正相关,即空气温度升高,赤皮青冈的蒸腾散热能力随之增强,说明3个树种中赤皮青冈对高温环境的适应性较强。这与张斌等的研究结果一致,原因是赤皮青冈具有较强的光合可塑性,可以通过调节气孔开合程度来改变其气体交换参数,从而适应环境的变化。
综上所述,3个壳斗科树种均存在“光合午休”现象,大叶青冈的光合固碳能力最强,用于造林栽培具有较高的经济效益和生态效益;赤皮青冈水分利用效率最高,且对强光和高温环境的适应性比大叶青冈和红椎强;红椎不适合在强光和高温环境下栽种,在培育幼苗过程中应适当遮荫。
猜你喜欢
智富时代(2019年4期)2019-06-01
智富时代(2019年4期)2019-06-01
东坡赤壁诗词(2018年6期)2018-12-22
广西农学报(2017年1期)2018-02-10
发明与创新·中学生(2017年5期)2017-05-12
中学化学(2016年10期)2017-01-07
中学化学(2015年8期)2015-12-29
作文与考试·小学低年级版(2015年22期)2015-12-07
鸭绿江(2009年9期)2009-10-14
中学生数理化·高考版(2008年2期)2008-11-01
