
中国兜兰属宽瓣亚属植物生存现状及保护成效
2022-07-19 01:06:58安明态武建勇杨焱冰
中国环境科学 2022年7期
张 央,安明态,武建勇,杨焱冰,李 志,叶 超
中国兜兰属宽瓣亚属植物生存现状及保护成效
张 央1,2,安明态1,2,武建勇3**,杨焱冰4,李 志1,2,叶 超1,2
(1.贵州大学林学院,贵州 贵阳 550025;2.贵州大学,生物多样性与自然保护研究中心,贵州 贵阳 550025;3.生态环境保护部南京环境科学研究所,江苏 南京 210042;4.贵州省林业科学研究院,贵州 贵阳 550005)
为明确兜兰属()宽瓣亚属(Subgenus.)植物各物种在我国的生存现状与濒危状况,在大量的野外调查和文献、标本数据收集的基础上,对其种群分布、数量、生境特征、市场贸易、受威胁因素、保护现状与保护成效进行统计分析,并依据《IUCN红色名录等级和标准(3.1版)》对该类群的濒危等级进行重新评估.结果表明,野外调查到硬叶兜兰()、麻栗坡兜兰()、杏黄兜兰()、白花兜兰()、巨瓣兜兰()、文山兜兰()、同色兜兰() 7个目标物种, 已知分布于我国50个县(市、区),共计194个自然分布点.宽瓣亚属植物在我国水平方向上主要分布于滇东南、黔西南、黔东北、滇西北、桂北与黔南交界处以及桂西北至桂西南等地区;垂直方向上集中分布于中高海拔段(780~1267m),平均海拔997m;在全国大尺度上,该亚属整体呈零散分布,区域小尺度上个体呈聚集生长;其小生境具有高海拔、透水、透气、喜阴、喜钙的特点.市场调查的贸易率为36.67%,贸易价格高低不等,具有较强的随机性;贸易来源主要为野外采挖,占贸易数量的88.32%,具有较强的地域性;贸易方式为现场和线上网络交易,具有较强的灵活性.该类群主要受到采挖和生境退化的威胁,分别占所有分布点的44.85%、29.83%.宽瓣亚属整体就地保率仅为34.53%,保护成效为一般保护;调查的29个保育地中,有20个保育地对该类群累计保育98次,除文山兜兰和绿叶兜兰外,其他7种迁地保护成效被评估为合适.巨瓣兜兰、文山兜兰由原来的濒危(EN)评估为极危(CR);麻栗坡兜兰、白花兜兰、杏黄兜兰评估等级未发生变化,仍为极危(CR);硬叶兜兰和同色兜兰由原来的易危(VU)评估为濒危(EN);德氏兜兰、绿叶兜兰被评为数据缺乏(DD).
兰科植物;兜兰属宽瓣亚属;生存现状;保护成效;濒危等级;中国
宽瓣亚属(Subgenus.)为兰科(Orchidaceae)兜兰属()的一个亚属,是兰科植物中观赏价值最高、受威胁最大的类群之一.根据有关资料记载[1-3],宽瓣亚属共有12种(不含变种,下同),我国产硬叶兜兰()、同色兜兰()、白花兜兰()、巨瓣兜兰()、文山兜兰()、杏黄兜兰()、德氏兜兰()、绿叶兜兰()、麻栗坡兜兰() 9种,其中硬叶兜兰和杏黄兜兰被合称为“金童玉女”,关注度极高. 宽瓣亚属植物的花形独特,具有极高的观赏价值,各地采挖严重,加上对生存环境要求极为严格,资源日益减少,很多种类已经面临严重威胁,保护迫切,受到国家及社会各界的高度关注,是生物多样性保护的“旗舰”类群.目前该亚属全部物种均列入《濒危野生动植物种国际贸易公约》(CITES)附录I和2021版《国家重点保护野生植物名录》,其中除硬叶兜兰外其他8种均为国家一级保护植物,9种列为《中国生物多样性红色名录—高等植物植物卷》中的受威胁物种(CR、EN、VU),3种列入《全国极小种群野生植物拯救保护工程规划(2011—2015年)》.
兜兰属植物首先起源于中国南部和东南亚,随后分散到东南亚群岛[4],主要分布于亚洲至太平洋岛屿的热带、亚热带地区[5],在我国分布于西南和华南地区,其中大多数分布于海拔1000m以上的较高地区[6],具有一定的区域性.宽瓣亚属植物则分布在滇东南地区与贵州、广西连成一片的岩溶地貌石灰岩地区[6],我国南亚热带地区作为兜兰的生态多样性中心[7],分布着丰富的兜兰属植物资源,其中以云南最多,然而在气候变化、人为采挖、生境退化、生物入侵等威胁下资源逐渐减少,目前在我国的具体分布及资源情况尚缺乏基础资料.兰科植物的贸易活动通常具有传统性、地域性和季节性,贸易种类具有普遍性和随意性[8],贸易活动也成为兰科植物濒危的重要原因之一[9],上世纪兜兰的走私贸易一度成为其濒危的最主要因素,尽管现阶段不存在大规模的贸易走私活动,但小规模的采挖贸易活动仍然威胁着兜兰的生存.我国兜兰属植物大多数种类局限生长在生态环境脆弱的喀斯特地区,且具有个体聚集生长的特点,人为采集时极易将所有个体挖走,并带到市场进行交易,导致局域性灭绝[10].保护植物多样性的有效方式有就地保护、迁地保护和野外回归[11],目前生长在保护区的宽瓣亚属植物到了较好的保护,而在全国大尺度上的保护状况不明;同色兜兰、文山兜兰具有较强的迁地保护适应性,而杏黄兜兰、麻栗坡兜兰迁地保护适应性较差[12],但整个亚属在全国的迁地保护情况不详,保护成效也未见报道.
野外生存现状的调查和评估是中国兜兰属植物保护工作的基础性资料[10],虽然国内外对宽瓣亚属植物的系统发育[13]、离体繁殖[14]、表皮形态[15]等,但从全国范围系统性对宽瓣亚属植物资源本底调查及现状评估的研究未见报道,部分相关研究尚不能完全反映其在我国当前的实际生存现状.因此,本研究基于2019年6月至2020年12月开展的兜兰属宽瓣亚属植物专项调查及多年来积累的野外数据,从该类群的种群分布、数量、生境特征、市场贸易、受威胁因素、保护现状与保护成效现状等方面来阐述中国宽瓣亚属植物的生存现状并对其濒危等级进行重新评估,在此基础上,提出保护建议,以期为该类群的保护管理提供参考依据.
1 材料与方法
1.1 调查方法
1.1.1 野生资源调查 在充分查阅《中国兜兰属植物》、《中国野生兰科植物原色图鉴》、物种2000中国节点(http://www.sp2000.org.cn/)、中国数字植物标本馆(https:/www.evh.ac.cn/)等资料和访问知情人士的基础上,选定云南、贵州、广西三省(自治区)为宽瓣亚属植物主要野外调查范围,并以滇东南、黔南、黔西南、桂北、桂西南等喀斯特地貌集中区为重点调查区域,其他区域补充调查,调查时间集中在2019年6月至2020年12月.采用直数法、样线法、样方法进行野外调查.针对物种分布不丰富、分布范围较散、种群数量较多的区域,设定具有代表性的样线进行调查;针对分布较散,分布面较积小的种类采用直数法,直接统计调查区域内物种的全部个体,实测其分布点的聚集面积;针对分布比较集中的种类,在分布区设立5m×5m的典型样方(至少按最密、中等、最稀疏分布地段各设置1个样方),对样方中的物种进行全面清查.调查过程中记录目的物种的GPS、海拔、地形地貌、生境、是否位于保护区、受威胁因素等基本信息.
1.1.2 迁地保育调查 根据我国大型植物园的分布情况,选择云南、贵州、广西、四川、重庆、江西,湖南,江西、湖北、福建、江苏、浙江、海南、上海、北京等15省(自治区、直辖市)为宽瓣亚属类群的迁地保育调查范围.以植物园、科研院所、花卉企业等为重点,共计29个迁地保育地.调查过程中记录物种名、保育地名称、来源、资源量、生长状况等信息,其中仅景东植物园的迁地保护数据来源于中国迁地保护植物大数据平台(https://espc.cubg.cn/ records/index/index.html).
1.1.3 市场贸易调查 市场贸易主要采用市场追踪调查,根据我国大型花鸟市场的分布及野外调查时访问目标物种的大致去向,再对相应地区进行市场追踪调查,调查范围涉及上述15省(自治区、直辖市).以大型花鸟市场和宽瓣亚属植物原产地的乡镇集市、临时花鸟售卖摊点为重点,共计30个贸易点.调查过程中记录物种名、市场名称、来源、资源量、单价、贸易方式等信息.
1.2 分布面积计算
按《IUCN物种红色名录濒危等级和标准(3.1版)》中的方法计算宽瓣亚属植物的分布区面积(Extent of occurrence, EOO)和占有面积(Area of occupancy, AOO)[16].本研究中宽瓣亚属植物的分布区面积即为宽瓣亚属植物194个分布点最外围点所围成的多边形面积.根据宽瓣亚属植物在野外整体零散,个体聚集的特点,结合李伟伟等[17]对国家重点保护野生植物三蕊草()的评估研究,确定本次占有面积的比例尺为2´2km.具体做法为:在本研究计算的分布区面积范围内绘制若干个2×2km的公里网格,占有面积=出现分布点的公里网格数×单位公里网格面积. 计算过程在ArcGIS 10.6软件中进行.
由于生境破碎化和宽瓣亚属植物在自然的分布往往呈现出多个个体聚集生长的特点,有时候分布区面积和占有面积的大小并不能反映物种的实际分布情况.为反映宽瓣亚属植物在野外的实际分布特征.本文对聚集面积(Aggregation area)作如下界定:宽瓣亚属植物在分布点上由多个个体聚集生长形成了密集的分布单元,该分布单元占地面积的大小即被称为聚集面积.当聚集面积较小时可进行实地测量,较大时则对分布单元最外围的个体进行坐标定位在ArcGIS 10.6软件中绘制多边形进行计算.
1.3 保护现状分析
1.3.1 就地保护现状 就地保护以分布点是否在保护区为标准,在保护区内视为得到保护,反之则无.物种就地保护率为位于保护区的某物种分布点个数/该物种总分布点数.参考秦卫华等[18]、周大庆等[19]的标准来评估宽瓣亚属植物的保护成效,当物种就地保护率大于75%时视为有效保护,50%至75%时视为较好保护,25%至50%时视为一般保护,不超过25%时视为较低保护.
1.3.2 迁地保护现状 迁地保护成效采用杨清等[20]的标准来定性评估宽瓣亚属植物在中国的迁地保护成效,即某物种在5个以上的保育地得到保存则为合适,反之不合适.
1.4 濒危等级评估方法
在覃海宁等[21]评估的《中国高等植物受威胁物种名录》基础上,依据IUCN物种红色名录濒危等级和标准3.1版中设立的(A-E)5项标准[16],结合调查结果分别对9种宽瓣亚属植物进行重新评估.当一分类单元符合A-E任一标准时,该分类单元即被列为相应的濒危等级,如果根据不同标准均能获得一定的等级,那么该分类单元应被置于风险最高的等级.
2 结果
2.1 宽瓣亚属种类组成及资源数量
该亚属在野外调查到硬叶兜兰、麻栗坡兜兰、杏黄兜兰、白花兜兰、巨瓣兜兰、文山兜兰、同色兜兰7个目标物种,共计50个县域分布点,194个地理分布点(表1),德氏兜兰和绿叶兜兰在野外调查未见.7个目标物种的数量均在500株以上,其中硬叶兜兰是该亚属中数量最多的物种(10000~12000株),其次是杏黄兜兰(1000~1500株)和文山兜兰(1000~1200株);同色兜兰、白花兜兰、麻栗坡兜兰、巨瓣兜兰4种数量均在500~800株的范围.由于德氏兜兰和绿叶兜兰在本次野外调查过程中未发现,基础数据缺乏,数量等级不详.

表1 兜兰属宽瓣亚属植物种类及数量等级表
2.2 宽瓣亚属植物空间分布现状
2.2.1 水平分布 由图1可见,宽瓣亚属在我国主要分布于滇东南、黔西南、黔东北、滇西北、桂北与黔南交界处以及桂西北至桂西南.在滇东南、黔西南至黔南、桂西南呈现出明显的集中分布,在滇西北、滇西南、黔中、黔东北、桂东北呈零散分布.其中硬叶兜兰具有分布点多、分布范围广的特点,主要在黔西南至黔南、黔中、黔东北、桂北、滇东南;同色兜兰主要集中分布于广西西南地区一带;麻栗坡兜兰分布较散,滇、黔、桂均有分布,该种是目前发现分布最北端的宽瓣亚属植物,最北端位于29.06°N的道真大沙河国家级自然保护区;白花兜兰分布于贵州黔南与广西北部地区的茂兰国家级自然保护区和木论国家级自然保护区;巨瓣兜兰主要分布在北盘江流域和兴义马岭河峡谷等低热河谷地带;文山兜兰分布于滇东南的文山一带;杏黄兜兰主要分布在我国云南怒江流域.从各省区分布情况来看,云南分布的种类最丰富,有硬叶兜兰、麻栗坡兜兰、巨瓣兜兰、同色兜兰、杏黄兜兰、文山兜兰6种;贵州分布有麻栗坡兜兰、巨瓣兜兰、硬叶兜兰、白花兜兰4种,广西分布有同色兜兰、白花兜兰、硬叶兜兰、麻栗坡兜兰4种.

图1 兜兰属宽瓣亚属植物野外分布示意
2.2.2 分布面积 宽瓣亚属植物194个分布点最外围点所围成的分布区面积(EOO)为586 888.83km2,而占有面积(AOO)仅为460km2,其中硬叶兜兰分布区面积最大达到了162039.11km2,最小的杏黄兜兰仅为435.65km2(表2).从宽瓣亚属植物分布点的聚集面积来看(表中“S”表示分布点的聚集面积,单位为m2),有148个分布点的聚集面积<100m2,占76.29%,分布点聚集面积在100~200m2的分布点有14个,占7.22%,200~300m2的分布点有5个,占2.58%,超过300m2的分布点有27个,占13.92%(表2).三种不同的分布面积表明,宽瓣亚属植物在野外总体表现出具有较大分布区面积的零散分布,在个体分布点上则呈现出小聚集分布的现象,野外调查过程中甚至还发现有的分布点面积仅有1~2m2.

表2 兜兰属宽瓣亚属植物分布面统计表

图2 兜兰属宽瓣亚属植物垂直分布示意
2.2.3 垂直分布 由图2可知,宽瓣亚属在垂直方向上分布海拔整体偏高,最高分布海拔为杏黄兜兰所在的2024m,最低分布海拔为155m,平均海拔997m.宽瓣亚属物种分布最丰富的海拔段为780~ 1267m,有硬叶兜兰、同色兜兰、麻栗坡兜兰、巨瓣兜兰、文山兜兰、白花兜兰6种;各个物种间的分布海拔段不同,其中硬叶兜兰分布海拔范围最大(333~ 1648m),适应海拔变化能力强,而杏黄兜兰分布海拔范围最小(1696~2024m),适应海拔变化能力较弱,生存范围狭窄.平均海拔最高的是杏黄兜兰(1829m),其次是文山兜兰(1398m)、硬叶兜兰(1040m),最低的是同色兜兰(541m).由此可见杏黄兜兰、文山兜兰、硬叶兜兰具有高海拔分布的特点,麻栗坡兜兰分布于中海拔地区,白花兜兰、同色兜兰具有低海拔分布的特点.
2.3 宽瓣亚属植物生境特征
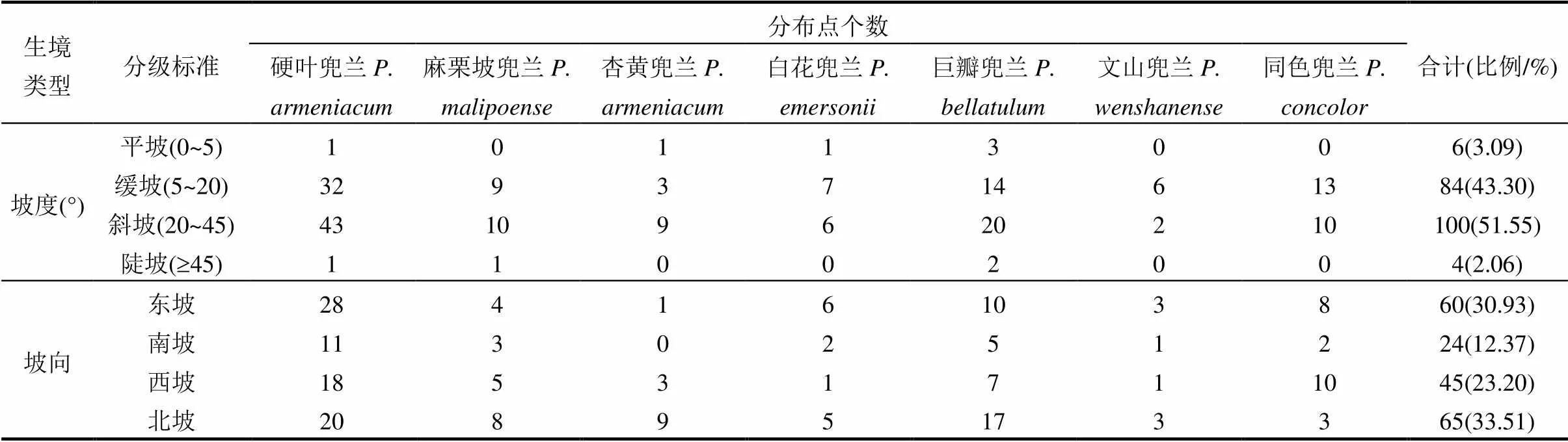
表3 兜兰属宽瓣亚属植物主要生境信息表

续表3
宽瓣亚属植物通常生于排水良好、透光、腐殖质丰富的林下.分布点的坡度在20°~45°有100个,占51.55%;分布点在北坡的有65个,占33.51%;分布点的土壤质地为壤土的有177个,占91.24%;分布点的地貌为喀斯特地貌的有178个,占91.75% (表3).通过对其生境调查统计表明,该类群生境具有高海拔、透水、透气、喜阴、喜钙的特点.
2.4 宽瓣亚属植物贸易现状
调查的30个贸易点中有11个贸易点对宽瓣亚属植物存在贸易活动,贸易率为36.67%,价格从20~120元不等,贸易来源主要为野外采挖,占贸易数量的88.31%,花卉基地仅占11.69%(表4).以贵阳阿栗花卉市场为主的4个城市花鸟市场的贸易来源主要为能提供稳定货源的花卉公司,且多为自主繁育的培育种;以兴义鲁布革乡集市为主的9个宽瓣亚属原产地的贸易来源主要为野外采挖,贸易方式为现场售卖和网络售卖.云南文山恒丰农贸市场贸易数量最多,达到了400株.从贸易物种看,硬叶兜兰是市场贸易频率最高和数量最多的物种,有7个贸易点合计约395株,占贸易总数量的64.12%,远远大于其他同类物种,且大多数为野外采挖所得.
另外,通过与贸易者访谈得知,大多数的宽瓣亚属植物均以零散销售为主,特别是在该属原产地的贸易市场.贸易方式一般在“赶集”时摆设临时摊点出售,或是在线上交易,贸易价格也因株型、花期而异,通常花期价格高于非花期,植株若有变形价格会更高,如“叶艺”、“花艺”.

表4 兜兰属宽瓣亚属植物市场贸易调查统计表
2.5 宽瓣亚属植物受威胁因素
调查结果表明,宽瓣亚属植物主要受到采挖、生境退化、工程建设、自然灾害、病虫害、动物啃食6个方面的威胁(图3).其中生境退化是指人为砍伐、放牧等活动引起分布点生境质量降低,导致宽瓣亚属植物在未来生长繁育过程中受到不良影响.该类群受到采挖威胁的有87个分布点,占44.85%,受生境退化受威胁的有57个分布点,占29.83%,由此可见人为采挖和生境退化是宽瓣亚属受威胁的主要原因.各物种间受威胁因素不一致,巨瓣兜兰、同色兜兰、文山兜、杏黄兜兰主要受采挖威胁;麻栗坡兜兰和硬叶兜兰主要受生境退化威胁;白花兜兰主要分布在保护区,受人为干扰较少,主要受威胁因素为自然灾害和病虫害.

图3 兜兰属宽瓣亚属植物受威胁现状
2.6 宽瓣亚属植物保护现状
2.6.1 就地保护现状 宽瓣亚属194个分布点中有67个位于保护区内,127个分布点位于非保护区,整体保护率为34.53%,就地保护成效为一般保护(表5).7个物种中达到有效保护的仅有白花兜兰1种,保护率为78.57%;达到较好保护也只有麻栗坡兜兰1种,保护率为50.00%;达到一般保护的有巨瓣兜兰、同色兜兰2种,保护率分别为46.15%、34.78%;文山兜兰、硬叶兜兰、杏黄兜兰的就地保护成效为较低保护,保护率均不超过25%.分布最广、数量最多的硬叶兜兰保护率仅为23.38%,远低于宽瓣亚属植物的总体保护水平.总的来说,宽瓣亚属植物就地保护力度偏低,保护成效不足,应着重加强就地保护.

表5 兜兰属宽瓣亚属植物就地保护统计表
2.6.2 迁地保护现状 29个保育地中保育宽瓣亚属植物的有20个,累计受保育98处(表6).保育地之间对物种的保育差异表现为规模大、科研能力强的单位保育物种丰富,如黔西南州绿缘动植物科技开发有限公司、中国科学院昆明植物研究所、上海辰山植物园、深圳市兰科植物保护研究中心4个保育地保育的物种数最多,均达到了8种,而南京中山植物园仅保存硬叶兜兰1种.物种间的保育差异表现为易获得和关注度高的物种保育次数较高,如硬叶兜兰(20处),其次是杏黄兜兰(16处)、同色兜兰(16处)、麻栗坡兜兰(13处),最少的是绿叶兜兰仅为2处.依据杨清等[20]的标准判断,宽瓣亚属植物整体迁地保护成效评估为合适;物种间,白花兜兰、德氏兜兰、巨瓣兜兰、麻栗坡兜兰、同色兜兰、杏黄兜兰、硬叶兜兰的迁地保护成效评估为合适,文山兜兰和绿叶兜兰评估为不合适.

表6 兜兰属宽瓣亚属植物迁地保护统计表

续表6
注:“√”表示该保育地收集保存了该物种;“—”表示该保育地没有收集保存该物种.
2.7 宽瓣亚属濒危等级评估
按照IUCN评估标准对其濒危等级进行重新评估.结果表明(表7),已知的7种均被评估为濒危(EN)以上,其中硬叶兜兰和同色兜兰由原来的易危(VU)评估为现在的濒危(EN),评估标准分别为B2b(ii)c(ii)和B2b(iii)c(ii),在实地调查中发现这两种种群占有面积少于500km2,栖息地生境退化明显,占有面积极度波动;麻栗坡兜兰、白花兜兰、杏黄兜兰评估等级未发生变化,仍然为极危(CR),但评判的标准有所增加,增加的标准分别是A2acd、A2acde、A2acd;巨瓣兜兰、文山兜兰由原来的濒危(EN)提升到极危(CR),评估标准分别为A1acd,A1acd.德氏兜兰和绿叶兜兰在本次评估中被评为数据缺乏(DD),原因是在野外调查过程中两种均未发现,部分保育地有收集保存,据相关资料记载[3],德氏兜兰原产越南,国内有栽培,在广西、云南有分布,绿叶兜兰在云南金屏有分布,经与相关兰科植物专家讨论,这两种在国内的野生分布可能有存疑,现状不明.

表7 兜兰属宽瓣亚属植物濒危等级评估结果
3 讨论
3.1 宽瓣亚属植物生存现状特征
种群数量是反映种群生存力及濒危等级评估的重要指标[22].我国兜兰属植物资源在20世纪八、九十年代已遭受了较为强烈的人为破坏[10],导致数量急剧下降,尽管有学者曾对我国兜兰属植物的生境和野生居群大小进行过调查[23],但这些工作并不全面.通过我们的此次的调查和评估,宽瓣亚属已知的7个种居群数量均在500株以上,最多的硬叶兜兰可达到10000株以上.尽管如此,现阶段的人为采挖与售卖现象仍有发生,特别是在兜兰丰富的滇东南与黔西南地区,若不及时加以制止,今后该类群的数量可能会由于采挖导致下降.另外,我们调查时发现宽瓣亚属植物在原生地能够较好的开花,但结实率偏低,传粉过程不稳定,未来的种群数量也可能波动不定.
在大尺度上物种的空间分布格局主要受到水热条件的影响[24],在小尺度上形成的空间分布格局主要受种群自身生物学特性的影响.我国西南地区喀斯特山地特殊的空间异质性维持了丰富的植物物种多样性[25],滇东南喀斯特地区也被认为是兜兰的起源和演化中心[6].调查结果显示,宽瓣亚属植物在我国主要分布在滇东南、黔西南、黔东北、滇西北、桂北与黔南交界处以及桂西北至桂西南,通过此次的深入调查该类群在我国的县域分布得到明显增加,分布在贵州道真县的麻栗坡兜兰(29.06°E)刷新了兜兰属中分布最北端(接近29°E)的记录.按照IUCN方法计算的宽瓣亚属分布区面积为586888.83km2,约占滇、黔、桂三省(自治区)陆地面积的73.76%,但占有面积仅为460.00km2,有148个分布点的聚集面积不超过100m2,占76.29%,这体现了宽瓣属植物在大尺度上整体呈零散分布,个体在区域小尺度上呈聚集生长的特点.
海拔主要是通过影响降水和热量的分配来影响物种的分布格局[26],甚至还会影响植物的生长发育[27].此次调查宽瓣亚属植物集中分布在780~ 1267m的海拔范围,最低海拔为155m,最高海拔为2024m,平均海拔为997m,各物种的最低海拔均比《中国兜兰属植物》记载的要低,其原因一方面是本次调查范围更宽泛、更深入,增加了一些未记载的分布点,另一方面可能与兜兰属从高海拔→广海拔→低海拔的演化趋势相对应[6].另外,植物所在的地形地貌差异往往导致光、温、水、土、气的差异,进而影响植物生长与分布[28].本次调查发现,宽瓣亚属植物总体主要集中分布在780~1267m的中高海拔、20°~45°的斜坡、北坡、壤土以及喀斯特地貌生境,该类群具有高海拔、透水透气、喜阴、喜钙的特点.因而在今后的迁地保护、野外回归及引种栽培的工作过程中应该选择具有以上特点的区域.宽瓣亚属植物在大尺度上的分布格局与我国西南喀斯特地区得天独厚的气候、水分、地形地貌等环境要素密不可分.兰科植物生活史与菌根真菌有着密不可分的关系,甚至影响其分布[29],宽瓣亚属在小尺度上所呈现的分布格局是否与菌根真菌有关还有待进一步研究.
3.2 宽瓣亚属植物贸易特点
80年代末至90年代中期是我国野生兜兰属植物贸易的高峰,自兜兰属植物全部列入《濒危野生动植物种国际贸易公约》(CITES)附录I以来以及近年来保护力度的加强,该类群大规模的市场贸易得到有效控制.由于兜兰属植物本身极高的观赏价值,使得其备受青睐,这也是兜兰属植物常常活跃于乡镇集市上的主要原因之一.本次调查发现,宽瓣亚属植物除绿叶兜兰外均出现在贸易市场上,贸易活动主要集中在兜兰的原产区,其中云南文山、贵州黔西南、广西崇左较为严重,具有较强的地域性,且来源以野外采挖为主;城市花卉市场对宽瓣亚属植物的贸易较为零散,来源以能够提供稳定进货渠道的花卉基地为主.宽瓣亚属植物贸易数量和贸易价格具有一定的随机性,数量和价格往往在物种的花期较高.贸易方式也较为灵活,除传统的现场贩卖外,网络销售成为新型、快捷的贸易渠道,例如通过组建兰友交流群,在微信、抖音、快手等新型媒体平台公开包邮售卖兜兰及其他兰科植物,销路也是全国各地,大多数兜兰也因此散落民间,生存状况不明.数量多、分布广的物种贸易频率高,例如硬叶兜兰在野外分布较广、数量多且多呈聚集分布,在野外更容易被发现和采挖.
3.3 宽瓣亚属植物致危因素
物种濒危原因主要是由内因和外因造成的.宽瓣亚属植物的传粉方式大多数为欺骗性传粉,特化的花结构,就需要与之匹配的特殊传粉者.兰科植物在种子萌发和原球茎生长阶段需要特定的共生真菌参与[30],宽瓣亚属植物也不例外.因此,特殊的生境需求、传粉媒介、共生真菌可能是导致其濒危的主要内因.但由于兜兰的特殊性,其濒危原因往往是由外因造成的,例如杏黄兜兰的主要濒危原因是直接采挖和生境丧失[31].本此调查也发现,宽瓣亚属植物87个分布点受到采挖威胁,占44.85%,57个分布点受生境退化的受威胁,占29.83%.可见该亚属受威胁的主要原因同样是采挖和生境退化.采挖主要是因其观赏价值极高,倍受社会上爱兰人士的青睐.因此,在有兜兰分布区域的一些村民很容易受到利益的驱使进行采挖兜兰活动.兜兰属个体聚集生长的特点,更容易被采挖[10],而数量较多、易被获得的群体更容易被贸易[32],这一特征常常导致该类群在野外被连片采挖带走流入市场进行贸易,从而导致其濒危.灵活的贸易方式也让这种采挖变得反复循环.生境破坏主要体现在土地用途的改变,建设过程中占用其生境或者由采石取砂引起的水土流失而造成生境破碎.除此之外,工程建设、自然灾害、病虫害、动物啃食也是威胁宽瓣亚属植物生存的重要原因.
3.4 宽瓣亚属植物保护成效
建立自然保护区已然成为保护珍稀濒危植物物种及其生境最有效的方式之一[33].本次野外调查到的7种宽瓣亚属植物除杏黄兜兰外均在保护区有分布,物种就地保护率为85.71%,高于我国1334种兰科植物51.9%的物种保护率[18],但物种分布点的就地保护率仅为34.54%,保护成效为一般保护,即还有大量的宽瓣亚属植物分布在保护区外未得到保护,处于保护空缺状态.例如分布于黔中地区的硬叶兜兰基本上不在保护区内.诚然,物种的自然分布会影响其保护率,如白花兜兰主要只分布在茂兰和木论两个国家级自然保护区内,如此一来保护率可能会偏高,但自从我国建立保护区以来,保护了65%的高等植物群落[34],特别是我国建立唯一的以兰科植物为保护对象的雅长兰科植物自然保护区更是保护了丰富的兰科植物资源.分布在保护区的物种由于保护区的管护力度较大而得到了有效保护,保护区外的物种由于保护力度不足更易容遭到人为破坏,可见采取以保护区(点)为主的保护措施是有效保护物种乃至生物多样性的重要途径.
迁地保护作为一种保护物种的常规手段,在物种多样性保护中具有重要作用[35].据统计,我国160个植物园迁地栽培的本土植物约为20000种,占我国本土高等植物科的91%[35],其中我国本土兰科植物821种,约占我国兰科植物总数的61.5%[36].宽瓣亚属各物种迁地保护率为100%,除文山兜兰和绿叶兜兰外其他物种迁地保护地均达5个以上,迁地保育合理.但有的保育地对物种的保育数量偏低,尚未达到11~21株的最小存活种群个体数[20],如绿叶兜兰,主要原因可能是该种在野外分布极狭域,很难进行引种采集.因此,在迁地保育工作上不仅要注重物种迁地保育次数,还需注重保育物种的数量.
4 结论
4.1 宽瓣亚属植物在野外调查到硬叶兜兰、麻栗坡兜兰、杏黄兜兰、白花兜兰、巨瓣兜兰、文山兜兰、同色兜兰7个目标物种,在我国主要集中分布于滇东南、黔西南、黔东北、滇西北、桂北与黔南交界处以及桂西北至桂西南等地区;垂直方向上集中分布于中高海拔段.该亚属植物具有整体呈零散分布,区域小尺度上个体呈聚集生长的分布特点.
4.2 宽瓣亚属植物主要受到采挖和生境退化的威胁,其市场贸易活动的价格具有随机性,贸易来源具有较强的地域性,贸易方式具有较强的灵活性.
4.3 宽瓣亚属整体就地保率仅为34.53%,保护成效为一般保护;文山兜兰和绿叶兜兰迁地保护成效不足,巨瓣兜兰、德氏兜兰、麻栗坡兜兰、白花兜兰、杏黄兜兰、硬叶兜兰、同色兜兰7种迁地保护成效合适.
4.4 宽瓣亚属植物除德氏兜兰、绿叶兜兰2种被评为数据缺乏(DD)外,其他均评估为濒危及以上等级,其中巨瓣兜兰、文山兜兰、麻栗坡兜兰、白花兜兰、杏黄兜兰5种被评估为极危(CR),硬叶兜兰和同色兜兰2种被评估为濒危(EN).
5 保护建议及对策
5.1 进一步完善法律保护及综合监管体系
宽瓣亚属植物已列入新版《国家重点野生植物保护名录》,得到了切实的法律保护.但目前乡镇集市以及网络平台的交易却未得到有效控制,在互联网快速发展下,网络线上成兰科植物贸易的活跃平台,面对新型网络平台销售模式下,缺乏针对性的法律法规,公开售卖,畅通无阻的快递运输,使得兰花交易更加频繁.因此,在新的国家重点保护名录颁布后还要针对市场、网络平台、快递运输等出台针对性的综合监管措施.
5.2 建立保护小区(点)并开展动态监测体系
为更好的掌握居群动态变化,实现物种长期有效监测,为今后的生态学、生理学、生殖学等综合学科研究提供研究基础.依据现阶段掌握的资料,我们建议每个物种在生境较好、具有一定种群数量、人为干扰较小、便于监测管理的区域设置1~2个保护小区或保护点,用于后期定期监测,同时也可作为野外回归的首选试验地.具体如下:(1)贵州黔中地区硬叶兜兰保护监测点;(2)贵州北盘江巨瓣兜兰保护监测点;(3)云南怒江杏黄兜兰保护监测点;(4)云南元江干热河谷文山兜兰保护监测点;(5)贵州黔西南麻栗坡兜兰保护监测点;(6)贵州黔南白花兜兰保护监测点;(7)广西崇左同色兜兰保护监测点.
5.3 加强种质资源宣传,提高全民保护意识
兜兰属植物分布区地处边远的山区,当地村民生产生活条件艰苦,靠山吃山的现状仍然存在,自然保护意识也不强,一旦某种植物能够产生经济效益,必然有村民进行采挖贩卖,甚至一些老百姓并不知道自己采集的物种是国家保护植物.因此,加强保护植物的宣传教育对其保护尤为重要,要让名录进入大众视野,采取多种灵活、喜闻乐见的形式进行法制教育宣传.例如,通过制作宣传手册、海报等发放给村民或是公示于村委会宣传栏;开发微信“二维码”、“小程序”等让普通大众扫一扫、搜一搜便可图文并茂的了解保护植物有关信息.如此一来便可增强普通大众对保护植物的认识.同时加大对不法分子的打击和惩治力度,使广大人民群众充分认识保护珍稀濒危植物的重要性和违法后果的严重性,从而提高全民自然保护意识.
5.4 引导兜兰属花卉资源扩繁及野外回归
科学合理规划并扩大人工培育力度,满足市场需求是保护兜兰属花卉资源的有效方式之一.兜兰属植物具有较高的花卉观赏价值,随着国民生活水平的提高,人们对兜兰属植物花卉的需求量越来越大,传统的采挖野生资源已对其生存环境和种质资源造成了巨大破坏,有些物种因此而消失.因此,必须对兜兰属植物原产地植物资源进行摸底调查并进行人工选育,利用现代生物技术进行有规模地合理开发利用,建立兜兰属花卉产业基地,以满足人们日益增长的对兜兰属花卉资源的需要,从而减少对野生资源的利用,同时加强野外回归,实现种群扩繁.
[1] 中国科学院中国植物志编辑委员会.中国植物志(17卷) [M]. 北京:科学出版社, 1999:52-62.
Editorial Committee of Flora of China. Flora Reipublicae Popularis Sinicae (Volume 17) [M]. Beijing: Science Press, 1999:52-62.
[2] Flora of China编辑委员会. Flora of China [EB/OL]. http://www. iplant.cn/foc, 2009.
[3] 刘仲健,陈心启,陈利君,等.中国兜兰属植物[M]. 北京:科学出版社, 2009.
Liu Z J, Chen S C, Chen L, Lei S, et al. The genusin China. Beijing: Science Press, 2009.
[4] Tsai C C, Liao P C, Ko Y Z, et al. Phylogeny and historical biogeography ofPfitzer (Orchidaceae) based on nuclear and plastid DNA [J]. Frontiers in Plant Science, 2020,11:1-14.
[5] 杨颖婕,黄家林,胡 虹,等.中国兜兰属植物种质资源保护和利用研究进展 [J]. 西部林业科学, 2021,50(5):108-112,119.
Yang Y J, Huang J L, Hu H, et al. Progress on conservation and utilization ofspecies in China [J].Journal of West China Forestry Science, 2021,50(5):108-112,119.
[6] 王英强.中国兜兰属植物生态地理分布 [J]. 广西植物, 2000,20(4): 289-294.
Wang Y Q. The geography of Chinese species of[J]. Guihaia, 2000,20(4):289-294.
[7] 龙 波,龙春林.兜兰属植物及其研究现状.自然杂志, 2006,28(6): 341-344.
Long B, Long C L. Amazingand its research status [J]. Chinese Journal of Nature, 2006,28(6):341-344.
[8] 宋亚琼,刘芝龙,Sophie Willian,等.西双版纳兰科植物集市贸易特点和保护启示[J]. 生物多样性, 2017,25(5):531-539.
Song Y Q, Liu Z L, Sophie W, Gao J Y, et al. Characteristics of the orchid trade at public markets and implications for conservation in Xishuangbanna, Yunnan, China [J]. Biodiversity Science, 2017,25(5):531-539.
[9] Phelps J. A blooming trade: illegal trade of ornamental orchids in mainland Southeast Asia (Thailand, Lao PDR, Myanmar) [R]. TRAFFIC. Petaling Jaya, Selangor, Malay, 2015.
[10] 罗毅波,贾建生,王春玲.初论中国兜兰属植物的保护策略及其潜在资源优势[J]. 生物多样性, 2003,11(6):491-498.
Luo Y B, Jia J S, Wang C L. Conservations trategy and potential advantages of the Chinese[J]. Biodiversity Science, 2003,11(6):491-498.
[11] 任 海,简曙光,刘红晓,等.珍稀濒危植物的野外回归研究进展 [J]. 中国科学:生命科学, 2014,44(3):230-237.
Ren H, Jian S G, Liu H X, et al. Advances in the reintroduction of rare and endangered rild plant species [J]. Scientia Sinica(Vitae), 2014, 44(3):230-237.
[12] 李秀玲,王晓国,李春牛,等.基于灰色关联分析方法评价13种野生兜兰的迁地保护适应性 [J]. 植物科学学报, 2015,33(3):326-335.
Li X L, Wang X G, Li C N, et al. Adaptability evaluation of ex situ conservation of thirteen wildspecies by Gray- Correlation analysis [J]. Plant Science Journal, 2015,33(3):326-335.
[13] Górniak M, Szlachetko D L, Olędrzyńska N, et al. Species phylogeny versus gene trees: A case study of an incongruent data matrix based onPfitz. (Orchidaceae) [J]. International Journal of Molecular Sciences, 2021,22(21):1-13.
[14] 曾宋君,郭贝怡,孔鑫平,等.兜兰离体快繁技术研究进展 [J]. 热带作物学报, 2020,41(10):2080-2089.
Zeng S J, Guo B Y, Kong X P, et al. Advances in propagation in vitro of[J]. Chinese Journal of Tropical Crops, 2020,41 (10):2080-2089.
[15] 查应琴,关 萍,陈 业,等.中国兜兰属23种植物叶表皮微形态特征初步研究 [J]. 植物科学学报, 2019,37(6):709-718.
Zha Y Q, Guan P, Chen Y, et al.Preliminary study on leaf epidermal micromorphological characteristics of 23ChinesePfitz. [J]. Species Plant Science Journal, 2019,37(6):709-718.
[16] IUCN Species Survival Commission. IUCN Red List Categories and Criteria: Version 3.1 [R]. IUCN Species Survival Commission, IUCN, Gland, Switzerland and Cam-bridge, UK, 2001.
[17] 李伟伟,张 慧,赵莉娜,等.青藏高原特有禾本科植物三蕊草受威胁等级评估 [J]. 生物多样性, 2012,20(6):133-137.
Li W W, Zhang H, Zhao L N, et al. An evaluation of the endangerment status of(Poaceae) endemic to the Qinghai-Tibet Plateau [J]. Biodiversity Science, 2012,20(6):133-137.
[18] 秦卫华,蒋明康,徐网谷,等.中国1,334种兰科植物就地保护状况评价 [J]. 生物多样性, 2012,20(2):177-183.
Qin W H, Jiang M K, Xu W G, et al. Assessment of in situ conservation of 1,334 native orchids in China [J]. Biodiversity Science, 2012,20(2):177-183.
[19] 周大庆,夏 欣,张昊楠,等.中国自然植被就地保护现状评价 [J]. 生态与农村环境学报, 2015,31(6):796-801.
Zhou D Q, Xia X, Zhang H A, et al. Evaluation of the status quo of natural vegetation conservation in China [J]. Journal of Ecology and Rural Environment, 2015,31(6):796-801.
[20] 杨 清,韩 蕾,许再富.中国植物园保护稀有濒危植物的现状和若干对策 [J]. 农村生态环境, 2005,21(1):62-66.
Yang Q, Han L, Xu Z F. Status of and strategy for ex-situ conservation of rare and endangered plants in Chinese botanical gardens [J]. Journal of Ecology and Rural Environment, 2005,21(1):62-66.
[21] 覃海宁,杨 永,董仕勇,等.中国高等植物受威胁物种名录 [J]. 生物多样性, 2017,25(7):696–744.
Qin H N, Yang Y, Dong S Y, et al. Threatened species list of china’s higher plants [J]. Biodiversity Science, 2017,25(7):696–744.
[22] 陈冬东,李镇清.极小种群野生植物生存力分析:方法、问题与展望 [J]. 生物多样性, 2020,28(3):358-366.
Chen D D, Li Z Q. Population viability analysis of wild plant with extremely small populations (WPESP): methods, problems and prospects [J]. Biodiversity Science, 2020,28(3):358-366.
[23] 覃文渊,覃国乐,覃文更,等.白花兜兰的群落结构特征分析 [J]. 北方园艺, 2012,(11):78-80.
Qin W Y, Qin G L, Qin W G, et al. Analysis of community structure of[J]. Northern Horticulture, 2012,(11):78-80.
[24] 朱耿平,刘 强,高玉葆.提高生态位模型转移能力来模拟入侵物种的潜在分布 [J]. 生物多样性, 2014,22(2):223-230.
Zhu G P, Liu Q, Gao Y B. Improving ecological niche model transferability to predict the potential distribution of invasive exotic species [J]. Biodiversity Science, 2014,22(2):223-230.
[25] 林 艳.AM真菌和枯落物对构树与云贵鹅耳枥种内种间养分竞争的影响[D]. 贵阳:贵州大学, 2019.
Lin Y. Effects of AMF and litter on the competition ofintraspecific and interspecific nutrient betweenand[D]. Guizhou University, Guiyang, 2019.
[26] 李小熊,李佶芸,喻 武,等.基于MaxEnt的青藏高原区光核桃适宜分布区预测 [J]. 西部林业科学, 2020,49(5):142-148.
Li X X, Li J Y, Yu W, et al. Simulation of suitable habitat distribution ofin Qinghai-Tibet plateau based on MaxEnt species distribution model [J]. Journal of West China Forestry Science, 2020,49(5):142-148.
[27] 刘旻霞,张娅娅,李全弟,等.甘南高寒草甸植物群落物种多度分布特征 [J]. 中国环境科学, 2021,41(3):1405-1414.
Liu M X, Zhang Y Y, Li Q D, et al. Species abundance distribution characteristics of alpine meadow plant community in Gannan [J]. China Environmental Science, 2021,41(3):1405-1414.
[28] Dordel J, Seely B, Simard S W. Relationships between simulated water stress and mortality and growth rates in underplanted Toona ciliata Roem. in subtropical Argentinean plantations [J]. Ecological Modelling, 2011,222(17):3226-3235.
[29] Mccormick M K, Jacquemyn H. What constrains the distribution of orchid populations? [J] New Phytologist, 2014,202(2):392-400.
[30] Rasmussen H N. Recent developments in the study of orchid mycorrhiza [J]. Plant and Soil, 2002,244:149-163.
[31] 董艳莉.杏黄兜兰的生物生态学特性及迁地栽培试验研究[D]. 北京:中国林业科学研究院, 2006.
Dong Y L. Bio-ecological characters and ex-situ conservation ofS. C. Chen et F. Y. Liu [D]. Beijing: Chinese Academy of Forestry, Beijing, 2006.
[32] 周志华,蒋志刚.野生动植物贸易活动的特点及影响因子研究 [J]. 生物多样性, 2005,13(5):462-471.
Zhou Z H, Jiang Z G. Features and impacting factors on trade of wildlife [J]. Biodiversity Science, 2005,13(5):462-471.
[33] 何友均,李 忠,崔国发,等.濒危物种保护方法研究进展 [J]. 生态学报, 2004,24(2):338-346.
He Y J, Li Z, Cui G F, et al. Advances in conservation methods of endangered species [J]. Acta Ecologica Sinica, 2004,24(2):338-346.
[34] 刘 珉.解读林业发展“十三五”规划——服务国家发展战略与林业现代化 [J]. 林业经济, 2016,38(11):3-8.
Liu M. Introducing the thirteenth five-year plan for forestry development——serving national development strategy and forestry modernization [J]. Forestry Economics, 2016,38(11):3-8.
[35] 黄宏文,张 征.中国植物引种栽培及迁地保护的现状与展望 [J]. 生物多样性, 20(5):559-571.
Huang H W, Zhang Z. Current status and prospects of ex situ cultivation and conservation of plants in China [J]. Biodiversity Science, 2012,20(5):559-571.
[36] 张玲玲,刘子玥,王瑞江.广东兰科植物多样性保育现状 [J]. 生物多样性, 2020,28(7):787-795.
Zhang L L, Liu Z Y, Wang R J. The conservation status of orchids in Guangdong Province [J]. Biodiversity Science, 2020,28(7):787-795.
致谢:感谢以下单位及相关人员在调查过程中提供帮助,他们分别是(按拼音排序):福建省厦门园林植物园、广西桂林植物园、广西木论国家级自然保护区、广西南宁植物园(广西青秀山管委会)、广西雅长兰科植物国家级自然保护区、广西中医药研究所、贵州大学生命科学学院、贵州荔波茂兰国家级自然保护区、贵州省林业科学研究院、贵州省植物园、贵州望谟县林业局、海南省三亚市林业科学研究院、湖北武汉植物园、华南植物园、南京中山植物园、黔西南州绿缘动植物科技开发有限公司、上海辰山植物园、深圳市兰科植物保护研究中心、无锡市鼋头渚风景区、仙湖植物园、云南林业科学研究院、中国科学院昆明植物研究所、中国科学院西双版纳热带植物园、中国科学院植物研究所.
Survival status and protection effectiveness ofSubgenus.in China.
ZHANG Yang1,2, AN Ming- Tai1,2*, WU Jian-Yong3**, YANG Yan-Bing4, LI Zhi1,2, YE Chao1,2
(1.Forestry of College, Guizhou University, Guiyang 550025, China;2.Research Center for Biodiversity and Nature conservation, Guizhou University, Guiyang 550025, China;3.Nanjing Institute of Environmental Sciences, Ministry of Ecology and Environment, Nanjing 210042, China;4.Guizhou Academy of Forestry, Guiyang 550005, China)., 2022,42(7):3461~3472
In order to clarify the survival and endangered status ofsubgenusin China, the population distribution, quantity, habitat characteristics, market trade, threatened factors, protection status and effectiveness of this group were statistically analyzed, based on a large number of field investigations, references and specimens data. And the endangered grade of this group was reassessed according to the IUCN Red List CategoriesandCriteria (version 3.1).The results showed that,,,,,andhad been found in the field investigation. And the seven Subgenera were known to be distributed in 50 counties in China, with a total of 194 natural distribution points. In the horizontal direction of China,Subgenuswere mainly distributed in Southeast and Northwest of Yunnan, Southwest and Northeast of Guizhou, Northwest and Southwest of Guangxi and the junction of Northern Guangxi and Southern Guizhou. In the vertical direction, the Subgenus mainly distributed in the middle-high elevation area (780~1267m), with an average altitude of 997m. On a national scale, the Subgenus were scattered distribution, and the individuals grew in aggregation on the small scale. Their microhabitats were characterized by high altitude, permeable, breathable, shady and high calcium. The market trade rate of Subgenuswas 36.67%, and the trade price had strong randomness; the trade source mainly depended on the extraction of wild resources which accounting for 88.32% of the trade volume, with strong regional characteristics; the trade mode was on-site and online with strong flexibility. This group was mainly threatened by excavation and habitats degradation, accounting for 44.85% and 29.83% of all distribution points respectively. The in-situ protection rate of Subgenuswas only 34.53%, which was general protection effectiveness; The group had been protected 98 times in 20 conservation sites, among the 29conservation sites which were investigated. Except forand, the ex-situ protection effects of the other 7 species were evaluated as appropriate.andwere assessed as critically endangered (CR) from the original endangered (EN); no change was happened in the assessment of,and, they were still belong to critically endangered (CR);andwere assessed as endangered (EN) from the original vulnerable (VU);andwere rated as data deficiency (DD).
Orchidaceae;Subgenus.;survival status;conservation effect;endangered grade;China
X176
A
1000-6923(2022)07-3461-12
张 央(1995-),男,贵州江口县人,侗族,贵州大学林学院硕士研究生,主要从事野生动植物保护与利用研究.发表论文3篇.
2021-12-30
生态环境部“生物多样性调查、观测和评估”项目(8-3-7-20-3);国家自然科学基金资助项目(31960042)
* 责任作者, 正高级实验师, gdanmingtai@126.com; ** 研究员, wujy10@hotmail.com
猜你喜欢
红领巾·萌芽(2024年3期)2024-04-22 04:40:32
红领巾·萌芽(2024年3期)2024-04-22 04:40:32
广西科学院学报(2022年2期)2022-07-14 03:55:52
浙江农业学报(2020年1期)2020-03-07 05:07:12
园林科技(2020年4期)2020-01-18 03:18:54
新农民(2019年29期)2019-02-21 06:35:38
浙江农业科学(2017年8期)2017-08-30 12:48:08
大众健康(2017年2期)2017-04-13 20:07:09
浙江农业科学(2016年5期)2016-12-07 07:03:27
环境与可持续发展(2016年6期)2016-03-13 19:06:03
