
冬虫夏草提取液对60Co γ 射线辐照小鼠睾丸组织氧化损伤的保护作用
2022-07-19 09:28魏闪闪王婷彭伟彪汪家春冯旭苏笠储智勇张阵阵
海军医学杂志 2022年4期
魏闪闪,王婷,彭伟彪,汪家春,冯旭,苏笠,储智勇,张阵阵
目前,电离辐照已广泛应用于工业、农业和医学领域,然而电离事故及放射医疗不可避免地对人体造成伤害,引起急性辐照综合征[1]。在电离辐照过程中,放射源在组织内呈指数衰减的形式分布,因此,入射通道的正常细胞及组织均会受到一定程度的损伤,可能引起并发症[2]。水是细胞的主要成分,电离辐照作用于水分子后,产生2 个初级自由基(OH-和H+),进而引起了一系列自由基链式反应,产生多种自由基,特别是辐照后细胞代谢产物活性氧(ROS)作用于线粒体膜电位,促使细胞的生理生化发生改变,诱导生物大分子的破坏、失活,甚至是细胞的凋亡[2-3]。已有研究结果表明,电离辐照导致的DNA 损伤90%是由自由基造成的,因此,清除自由基氧化损伤是防辐照损伤的重要途径之一[3]。
外源性损伤修复药物主要通过直接清除体内自由基来保护机体,进而有效减少辐照损伤[4]。一些中药及其中的活性成分通过减少辐照或光解水产生的OH-自由基,防止自由基对机体产生氧化损伤,从而具有良好的抗氧化功效[5]。因此,开发新的天然辐照防护剂及其活性成分不仅具有广泛的应用前景,而且具有重点开发的科学价值。
作为最早收载冬虫夏草的国家,中国首次于公元8 世纪的《月王药诊》中记载冬虫夏草具有治疗肺部疾病的功效[6]。随后,《千万舍利》《四川通志》《药性考》及《景岳全集》等记载冬虫夏草性温暖,秘精益气,专补命门,能益肾补肾,化痰止咳[7-8]。现代药理学研究表明,冬虫夏草具有抗肿瘤、保护小脑、抗肝纤维化、抗衰老、保护胃和抗糖尿病、壮阳、抗疲劳、免疫调节等多种功效[9-11]。冬虫夏草虽已有上千年的用药历史,但对其功效的研究依然处于初步阶段。本研究测定了辐照小鼠睾丸组织中抗氧化酶活性、脂质过氧化物产物含量、氧化酶相关的基因表达等,初步探讨了冬虫夏草提取液(Cordyceps sinensisexact,CSE)对辐照诱导小鼠睾丸组织氧化损伤的保护作用。
1 材料与方法
1.1 材料
1.1.1 实验药品 野生冬虫夏草产自青海玉树,购买于雷允上药业集团有限公司,由海军军医大学药学院黄宝康教授鉴定为真品。CSE 采用了水提法进行制备[12],由海军特色医学中心(原海军医学研究所)制剂室提供(批号:20161201)。
1.1.2 实验动物 50 只SPF 级C57BL/6 雄性小鼠(体重20~25 g)购于上海西普尔必凯实验动物有限公司,生产许可证号:SCXK(沪)2013-2016。所有小鼠放置温度为22~26 ℃、湿度为44%~65% 的室内动物房中进行饲养,并给予正常饮食、饮水,饲养1 周以适应生长。本次实验已通过海军军医大学伦理委员会审核,且相关操作均遵守实验动物管理规范和适用原则。
1.1.3 动物分组 50 只SPF 级C57BL/6 雄性小鼠,按数字表法随机分为未辐照组(Control 组)、辐照组(Model 组)、低剂量CSE(CSE-L)组(生药剂量312.5 mg/kg)、中剂量CSE(CSE-M)组(生药剂量625.0 mg/kg)和高剂量CSE(CSE-H)组(生药剂量1 250.0 mg/kg),共5 组,每组各10 只。
1.1.4 试剂 生理盐水(批号:160530D13)购于黑龙江福和华星制药集团股份有限公司;丙二醛(malondialdehyde,MDA)检测试剂盒(S0131)、总超氧化物歧化酶(superoxide dismutase,SOD)活性检测试剂盒(S0101)、4%的多聚甲醛(P0099)均购于碧云天生物科技有限公司;髓过氧化物酶(myeloperoxidase,MPO)一抗购买于Cell Signaling Technology;PrimeScriptTMRT Master Mix(Perfect Real v Time)(RR036A)、RNAiso Plus(9109)均购于Takara;SybrGreen qPCR Master Mix(QR-SG-M200)购于Rainbio 公司。
1.1.5 实验仪器 ECLIPSE TI-SR 倒置荧光显微镜(日本NIKON 公司);ECLIPSE C1 正置荧光显微镜(日本NIKON 公司);SpectraMax M5/M5e 多功能酶标仪(美国分子仪器公司);CFX96 Touch 定量PCR 仪(美国Bio-rad 公司)。
1.2 方法
1.2.1 建立电离辐照损伤模型 除Control 组外,其余4 组小鼠均放置于塑料盒中固定,给予单次8 Gy60Co γ 射线全身辐照(辐照率为0.96 Gy/min)。辐照4 h 后,CSE-L、CSE-M 与CSE-H 组灌胃给予相应生药剂量的CSE,Control 及Model 组小鼠给予等体积量的生理盐水。所有小鼠在相同的实验环境下,每日于14:00 进行灌胃给药/生理盐水1 次,共持续28 d。在此次实验过程中,Control、CSE-M 及CSE-H 组小鼠均未出现死亡现象,Model 组小鼠死亡4 只,CSE-L 组小鼠死亡1 只。
1.2.2 标本收集 各组小鼠灌胃给药/生理盐水28 d 后,3% 水合氯醛麻醉小鼠,切开阴囊后,将2 只睾丸取出并用生理盐水洗涤干净,1 只保存于4% 体积分数的多聚甲醛中,另1 只液氮速冻后放于-80 ℃冰箱中保存。
1.2.3 睾丸组织中MDA 水平的检测[13]取(20 ±2)mg 睾丸组织,加入200 μl 蛋白裂解液至细胞粉碎机中研碎,离心,收集上清中的蛋白提取液。聚氰基丙烯酸正丁酯(BCA)蛋白浓度测定试剂盒检测待测样品中的蛋白浓度。根据说明书将蛋白裂解液/标准品/待测样品、MDA 检测工作液加入至0.6 ml 的PCR 管中,加热15 min,冷却,离心半径8.6 cm,离心力(RCF)为1 000×g,离心10 min,取200 μl 各组上清至新的平底96 孔板,放入Spectra-Max M5/M5e 多功能酶标仪中,分别测定各孔溶液在532 nm 处的吸光度。按照该试剂盒说明书画出标准曲线,并分别计算各组样品中MDA 水平。
1.2.4 睾丸组织中SOD 活力的检测[13]取(10 ±2)mg 睾丸组织,加入100 μl SOD 样品制备液至细胞粉碎机中研碎,RCF 为12 000×g(离心半径8.6 cm),离心10 min 后取上清作为待测样品。按照说明书将各组待测样品/SOD 检测缓冲液、对应浓度的WST-8/酶工作液、反应启动工作液加入至96 孔板中。在37ºC 环境下孵育30 min 后,放入Spectra-Max M5/M5e 多功能酶标仪中测定样品管与对照管450 nm 处的吸光度。分别计算各组样品中SOD 活力。结果显示,CSE-H 组小鼠睾丸组织中的SOD 活力显著性高于Model 组(P<0.05),而CSE-L 组及CSE-M 组与Model 组相比差异无统计学意义(P>0.05),因此,本研究后续将进一步采用了苏木精伊红(HE)及TUNEL 染色、qRT-PCR、Western blotting实验对Control、Model、CSE-H 组小鼠的睾丸组织进行了检测。
1.2.5 HE 染色检测小鼠睾丸组织形态[14]取出的睾丸组织多聚甲醛固定后,经TP1020 自动脱水机脱水,进行石蜡包埋,EG 1150C 超薄切片机上做超薄石蜡切片、烤片、切片透明化、脱蜡、HE 染色、封片。将处理好的切片至倒置荧光显微镜,并采集Control 组、Model 组、CSE-H 组小鼠睾丸组织HE 切片图像。
1.2.6 MPO 染色检测小鼠睾丸组织炎性浸润细胞数目[14]将石蜡切片脱蜡至水,抗原修复,阻断内源性过氧化酶,血清封闭,孵育一抗、二抗、二氨基联苯胺(3,3'-diaminobenzidine,DAB)显色,复染细胞核,脱水封片后,放于荧光显微镜下,图像采集分析Control 组、Model 组、CSE-H 组小鼠睾丸组织MPO 图像。
1.2.7 qPCR 检测睾丸组织中凋亡相关基因的表达[11]取(10 ± 2)mg 睾丸组织,放于1 ml 的Trizol中,用细胞粉碎机进行研碎。加入200 μl 氯仿,充分漩涡,静置,离心取上清并加入等量的异丙醇,上下颠倒混匀后静置,离心弃上清。用无酶的75%乙醇溶液洗涤3 次,干燥,加入适量的无酶DEPC 水,混匀。根据RNA 浓度将各组的RNA 进行定量,逆转录形成cDNA 后,于96 孔PCR 板中进行qPCR,计算Control 组、Model 组、CSE-H 组小鼠睾丸组织中谷胱甘肽过氧化物酶(glutathione peroxidase,GSH-Px)、过氧化氢酶(catalase,CAT)及聚ADP-核糖聚合酶1(poly ADP-ribose polymerase1,PARP1)基因的表达。引物序列见表1。
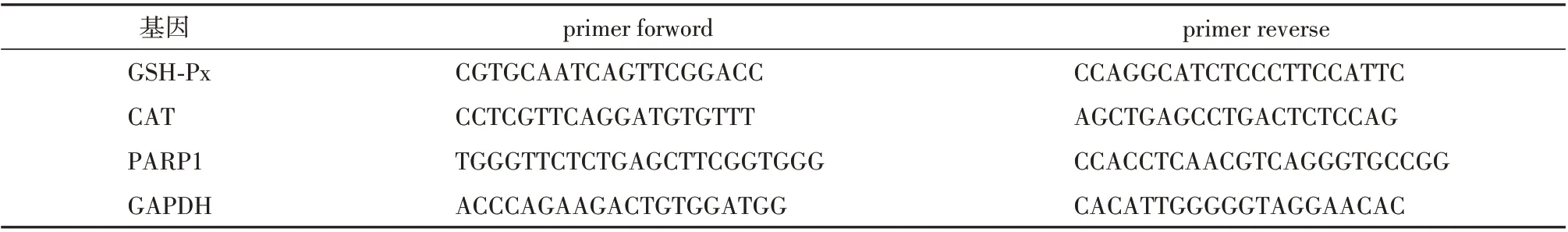
表1 PCR 起始序列(primer sequence)
1.3 统计学处理
采用Graphpad 7.0 软件对实验数据进行分析,组间比较采用单因素方差分析(one-way ANOVA)。P<0.05 表示差异有统计学意义。
2 结果
2.1 CSE 对60Co γ 射线辐照诱导小鼠睾丸组织中MDA 水平及SOD 活力的影响
与Control 组相比,Model 组小鼠睾丸组织MDA水平提高,SOD 活力降低,差异均有统计学意义(P<0.01)。与Model 组相比,CSE-L、CSE-M、CSE-H 治疗组中MDA 水平呈剂量依赖性明显降低(P<0.01);CSE-H 治疗组中SOD 活力升高,差异有统计学意义(P<0.05)。见表2。
表2 CSE 对60Co γ 射线辐照小鼠睾丸组织中MDA 水平及SOD 活力的影响(± s)
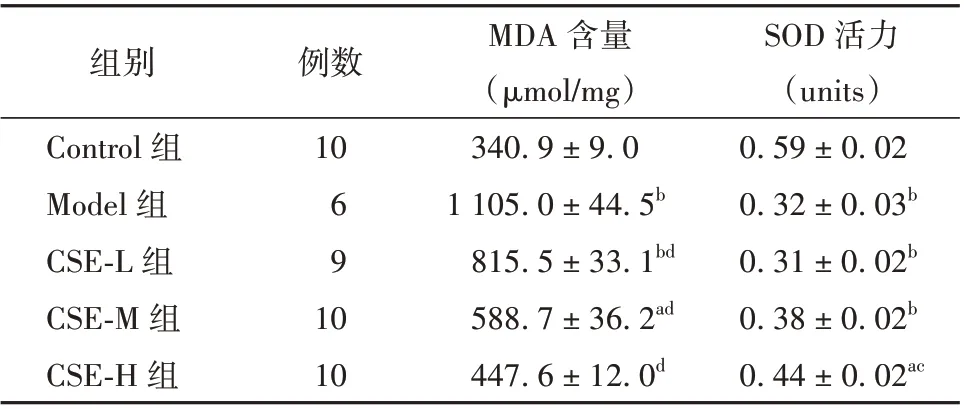
表2 CSE 对60Co γ 射线辐照小鼠睾丸组织中MDA 水平及SOD 活力的影响(± s)
注:与Control 组相比aP<0.05,bP<0.01;与Model 组相比cP<0.05,dP<0.01。CSE 为冬虫夏草提取液,MDA 为丙二醛,SOD 为超氧化物歧化酶;L、M、H 分别代表低、中、高剂量
SOD 活力(units)0.59±0.02 0.32±0.03b 0.31±0.02b 0.38±0.02b 0.44±0.02ac组别Control 组Model 组CSE-L 组CSE-M 组CSE-H 组例数10 69 10 10 MDA 含量(μmol/mg)340.9±9.0 1 105.0±44.5b 815.5±33.1bd 588.7±36.2ad 447.6±12.0d
2.2 CSE 对60Co γ 射线辐照诱导小鼠睾丸组织HE病理学的影响
HE 病理结果显示,Control 组小鼠睾丸组织中生精管膜完整,曲细管、精管间的间质组织结构紧密,排列整齐,层次较清晰,精子较丰富。Model 组小鼠睾丸组织间质结构变疏松,间质细胞不同程度地变性,同时,生精上皮明显变薄,腔内空虚。CSE-H 组睾丸组织的形态完整,结构紧密。见图1。该结果提示CSE可以缓解由辐照引起的睾丸组织结构损伤。
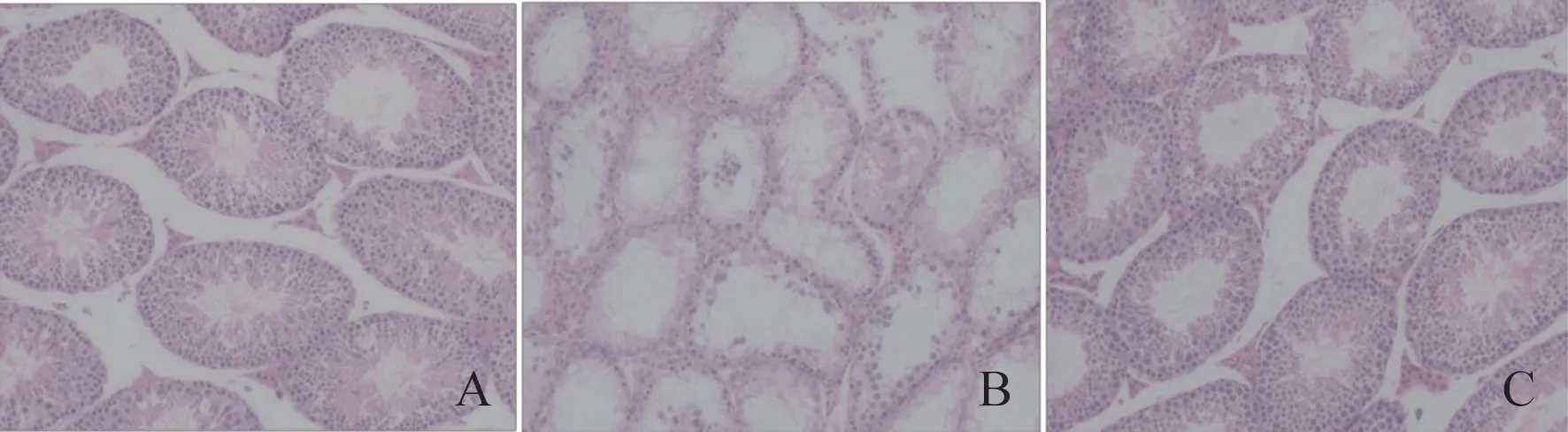
图1 CSE 对60Co γ 射线辐照诱导小鼠睾丸组织病理学的影响(HE×200)
2.3 CSE 对60Co γ 射线辐照诱导小鼠睾丸组织MPO 病理学的影响
MPO 染色结果显示,Control 组、Model 组、CSEH 组小鼠睾丸组织中MPO 阳性细胞数目分别是7.667 ± 1.453、38.000 ± 2.887、18.000 ± 1.732。与Control 组相比,Model 组中小鼠睾丸组织中炎症细胞浸润数目显著增加(P<0.01);与Model 组相比,CSE-H 组中小鼠睾丸组织MPO 阳性细胞数目明显降低(P<0.01)。结果提示高浓度CSE 能有效抑制由辐照引起的睾丸组织中炎性细胞浸润的发生。
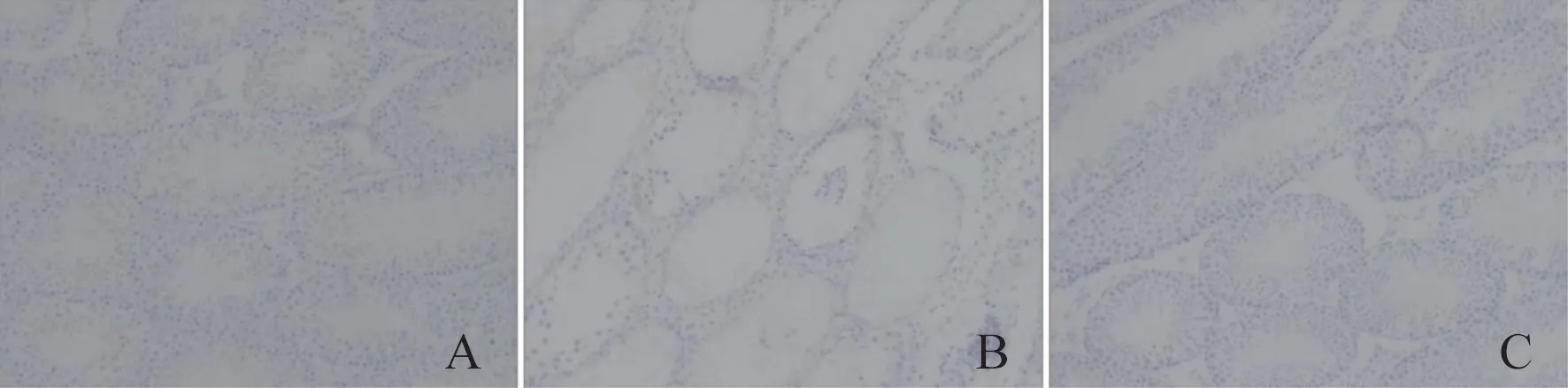
图2 CSE 对60Co γ 射线辐照诱导小鼠睾丸组织病理学的影响(MPO×400)
2.4 CSE 对60Co γ 射线辐照诱导小鼠睾丸组织中PARP1、GSH-Px、CAT 基因mRNA 相对表达量的影响
与Control 组相比,Model 组中小鼠睾丸组织中PARP1 基因mRNA 相对表达量明显上升(P<0.01),GSH-Px、CAT 基因mRNA 相对表达量明显降低(P<0.01)。与Model 组相比,CSE-H 组中GSH-Px 基因mRNA 相对表达量明显上调,PARP1 基因mRNA 相对表达量明显下降,差异均有统计学意义(P<0.01)。见表3。该实验结果表明,CSE 能有效地改善上述与凋亡相关的基因表达。
表3 CSE 对60Co γ 射线辐照诱导小鼠睾丸组织中PARP1、GSH-Px、CAT 基因mRNA 相对表达量的影响(± s)
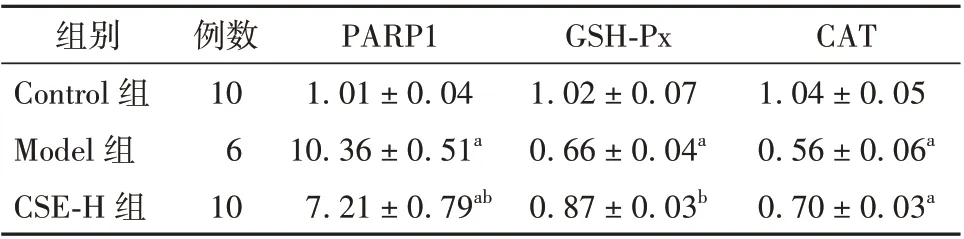
表3 CSE 对60Co γ 射线辐照诱导小鼠睾丸组织中PARP1、GSH-Px、CAT 基因mRNA 相对表达量的影响(± s)
注:与Control 组比较aP<0.01;与Model 组比较bP<0.01。CSE为冬虫夏草提取液,PARP1 为聚ADP-核糖聚合酶1,GSH-Px 为谷胱甘肽过氧化物酶,CAT 为过氧化氢酶,H 代表高剂量
CAT 1.04±0.05 0.56±0.06a 0.70±0.03a组别Control 组Model 组CSE-H 组例数10 6 10 PARP1 1.01±0.04 10.36±0.51a 7.21±0.79ab GSH-Px 1.02±0.07 0.66±0.04a 0.87±0.03b
3 讨论
电离辐照的应用是把双刃剑,在广泛应用于农业、工业、医学、军事等领域的同时,电离事故及放射医疗也给人体的正常组织造成了损伤[1]。对于电离辐照引起的细胞及机体损伤,能利用一些辐照防护剂进行预防及治疗。目前,已有的抗辐射药物虽然疗效明显,但用药量大,且具有很强的不良反应,如恶心、呕吐、头痛等[15]。因此,开发疗效佳、无不良作用、安全可靠的辐照防护剂迫在眉睫。
冬虫夏草作为传统中药,可用于治疗多种疾病[12]。已有报道显示,CSE 可以保护137Cs 辐照小鼠的骨髓造血干细胞及小肠组织损伤,降低UV 电离辐照小鼠皮肤组织中的ROS、H2O2及DNA 损伤水平[16];提高辐照小鼠小肠中的隐窝干细胞活力,降低小肠中凋亡细胞的数目;冬虫夏草多糖(CS-PS)能有效地提高4 Gy60Co γ 小鼠脾脏单核细胞的活力[10,12,17]。然而,CSE 对电离辐照后睾丸组织氧化损伤是否有保护作用尚不清楚。因此,根据已有的研究,本实验进一步研究了CSE 抗辐照作用及其机理,以期深入挖掘应用CSE 进行临床治疗的潜在价值。
生物分子损伤是辐射生物效应的物质基础,电离辐照生物学效应的根本原因主要分为放射线与物质的相互作用导致的生物分子电离和激发的直接作用以及其产生的自由基导致的继发作用。辐照后激发的机体电离反应会造成大量的自由基发生脂质过氧化反应,进而导致机体的氧化损伤,破坏正常生理状态下的抗氧化防御系统的平衡,最终导致机体不能有效地对辐照损伤起到抑制作用。SOD 是抗氧化系统的第一道防线,在体内氧化与抗氧化平衡中起着重要作用,可通过歧化作用将O2-转化成H2O2,清除O2-,降低OH-的含量,并发挥抗氧化作用保护细胞免受损伤[18]。MDA 是脂质过氧化物的分解产物,能够与蛋白质、核酸、磷脂等含有氨基的物质进行交联,促使了大分子化合物的分子交联,进而导致细胞代谢异常及功能破坏[18]。因此,增强SOD 活性和降低MDA 水平可以有效清除辐射引起的自由基,并对辐射损伤起到修复和防护作用。本研究结果显示,60Co γ 射线辐照后,小鼠睾丸组织中的MDA 水平明显上升,SOD 活力显著下降,反映了辐照后小鼠睾丸组织抗氧化能力的降低,而CSE 治疗后,Model 组小鼠睾丸组织中的MDA 水平和SOD 活力均得到了改善性调节,由此可知,CSE可以通过睾丸组织中MDA 水平下调、SOD 活力上调来提高抗氧化能力。该结果与虎杖苷提高辐照后睾丸组织的抗氧化能力的研究结果类似。
Khan 等[19]认为,电离辐照产生较多自由基和DNA 损伤后,将可能进一步导致机体组织出现炎性浸润、充血、增生等病变。为进一步研究CSE 对电离辐照小鼠睾丸组织的保护作用,本研究对Control组、Model 组、CSE-H 组小鼠的睾丸组织进行了HE、MPO 染色,结果显示,Model 组小鼠睾丸组织间质结构变疏松,腔内变空,炎性细胞数目增加。CSE 治疗后电离辐照小鼠睾丸组织变完整,结构紧密,炎性细胞数目减少。本实验结果表明,CSE 保护了小鼠睾丸组织的形态结构,降低了辐照后生精细胞中的炎性细胞数目,其作用机制可能与抗氧化相关。
为进一步验证CSE 的抗氧化功能,本研究采用了qRT-PCR 对各组小鼠的睾丸组织中的抗氧化相关的基因进行了检测。PARP1 是一种细胞核酶,可催化聚二磷酸腺苷(adenosine diphosphate,ADP)核糖基化,不仅在蛋白质的翻译后修饰过程中发挥重要的功能,而且在修复DNA 单链损伤和氧化应激造成的DNA 氧化损伤等响应中占有重要的角色[20]。GSH-Px 广泛分布于细胞质、线粒体中,具有清除生物体内自由基、阻断脂质过氧化链式反应、保护细胞膜的结构及功能的作用,是一种关键的催化氧化酶[21]。CAT 主要是以H2O2为底物,催化相关的电子转移,并将其分解成水、氧气,有效地阻止其对机体有害地羟基自由基的生成,进而清除生物体内的H2O2[11,20]。研究发现,降低PARP1 和提高GSH-Px、CAT mRNA 的表达水平可以有效地抑制脂质过氧化产生的细胞损伤[9,20]。qPCR 实验结果显示,与Control 组相比,Model 组小鼠睾丸组织与抗氧化相关的GSH-Px、CAT 基因mRNA 相对表达量均明显降低,PARP1 基因mRNA 相对表达量明显上升;CSE 治疗后,PARP1 基因mRNA 相对表达量明显下降,GSH-Px 基因mRNA 相对表达量明显上升。该结果表明,CSE 保护睾丸组织可能是通过抑制氧化损伤来起作用。但是关于CSE 对辐照小鼠睾丸组织抗氧化功能影响的具体作用成分因子,以及其作用于辐照损伤反馈信号的分子机制,仍有待进一步的研究分析。
猜你喜欢
中国临床医学影像杂志(2022年5期)2022-07-26
发明与创新·小学生(2021年12期)2021-01-06
军事文摘(2020年22期)2021-01-04
中学生数理化(高中版.高考理化)(2020年10期)2020-10-27
中学教学参考·理科版(2016年3期)2017-05-19
汉语世界(The World of Chinese)(2016年3期)2016-01-10
中学化学(2015年10期)2015-12-14
中学化学(2014年2期)2014-04-23
小学阅读指南·低年级版(2009年4期)2009-05-13
现代家庭(1999年7期)1999-06-14
