
宫内发育迟缓小鼠妊娠期胰岛lncRNA和mRNA的表达谱分析
2022-07-16 04:59:02袁庆新
南京医科大学学报(自然科学版) 2022年7期
王 莉,袁 逸,唐 艺,孙 璐,袁庆新
南京医科大学第一附属医院内分泌科,江苏 南京 210029
糖尿病受遗传因素和环境因素共同作用,随着健康与疾病发育起源理论的提出,“胎源性疾病”受到越来越多的重视,子宫内环境改变、胚胎宫内发育不良及表观遗传的变化,将影响其成年后代谢性疾病的发生,这些个体成年后怀孕又将出现进一步代谢异常,这种跨代效应导致糖尿病发病率不断增加,从胚胎发育及孕期入手探究糖尿病的发病机制是目前热点之一。
宫内发育迟缓(intrauterine growth retardation,IUGR)是常见的胚胎发育异常形式,可以导致新生期个体的胰岛面积明显减少,胰岛素染色减弱,胰岛较正常少且松散,血清胰岛素含量低于正常,成年后容易发生糖耐量异常[1],这些IUGR雌性个体成年后,面对怀孕这一压力测试,可能出现进一步的代谢失衡、胰岛功能障碍。
许多研究已证实正常妊娠期母体会经历一系列的生理变化,以满足自身和胎儿不断变化的营养需求,包括心血管系统、肾脏系统、血液系统、呼吸系统和内分泌系统的适应性改变。孕期雌激素、孕激素、胎盘催乳素、胎盘生长激素、脂联素、瘦素等水平的变化及碳水化合物代谢的增加,导致生理性胰岛素抵抗,胰岛β细胞团出现代偿性增殖,从而保证妊娠期母体的糖代谢平衡。IUGR 雌性个体成年后妊娠期会出现怎样的代谢及胰岛功能改变,其可能机制如何,目前少有研究。
文献表明,成年期的糖代谢异常可能是其生命早期环境印迹的“程序化”结果,表观遗传学在其中发挥着重要作用[2]。表观遗传主要包括DNA 甲基化、组蛋白修饰、染色质重塑、非编码RNA 等,是指基因的核苷酸序列不发生改变的情况下,基因表达发生可遗传的变化。非编码转录本依据其大小分为两类,具有200个或更少核苷酸的(nucleotide,nt)为小非编码RNA,包括微小RNA(microRNA,miRNA)、与piwi 蛋白作用的RNA(piwi-interacting RNA,piRNA)和小干 扰RNA(small interference RNA,siRNA);超过200 nt 的非编码RNA 为长链非编码RNA(long non-coding RNA,lncRNA),不编码蛋白质,结构上与信使RNA(messenger RNA,mRNA)相似,但缺乏开放阅读框,位于细胞质或细胞核内,核lncRNA 主要参与表观遗传基因调控或维持核结构,而细胞质lncRNA 参与转录后基因调控[3]。在人胰岛和纯化的β细胞[4]以及小鼠胰岛细胞[5]已经发现有1 000 多条lncRNA 表达,部分参与胰岛β细胞的分化成熟过程,且表达受糖浓度调节。lncRNA的表达在时间和空间上都受到严格的调控,因此失调的表达谱是疾病或发育状态改变的重要标志。
本研究首先通过孕期给予母鼠8%的低蛋白饮食获得IUGR 新生小鼠,然后待新生小鼠成年后与同周龄雄鼠配繁,最后获得IUGR孕鼠。对IUGR孕鼠(IUGR pregnancy,IP)和正常孕鼠(normal pregnancy,NP)胰岛RNA 进行高通量测序分析,比较两者lncRNA 和mRNA 表达差异,对 差异lncRNA 的靶mRNA 和差异mRNA 进行功能注释和富集通路分析,为糖尿病的早期预防和诊治提供新思路。
1 材料和方法
1.1 材料
8 周龄C57BL/6 雄性小鼠16 只、雌性小鼠32 只购自上海南方模式生物科技股份有限公司,动物饲养和实验均通过南京医科大学实验动物福利伦理委员会(IACUC)批准。小鼠胰岛β细胞系min6 细胞来自南京医科大学第一附属医院内分泌科实验室。
RNA isolater Total RNA Extraction Reagent、SYBR Green Master Mix(南京Vazyme 公司);Primer-ScriptTMRT Master Mix(TaKaRa 公司,日本);RNA 引物(上海Invitrogen 公司);Histopaque1077(Sigma 公司,美国)。PCR扩增仪(Eppendorf公司,美国);实时定量PCR 仪(Thermo Fisher 公司,美国);Nano Drop超微分光光度仪(Thermo Fisher公司,美国)。
1.2 方法
1.2.1 实验动物造模
8 周龄C57B6/L 小鼠饲养在SPF 级环境。待小鼠适应环境后,傍晚雄性与雌性以1∶2合笼,次日清晨以阴道有精栓且阴道涂片发现精子为确定妊娠。
NP 组:自确定妊娠开始予以标准繁殖饲料(蛋白含量为20%)。IP 组:①IUGR 小鼠造模。孕鼠自确定妊娠开始予以低蛋白饲料(购自北京协同生物有限公司,等热量、8%的蛋白质含量)至新生小鼠出生;IUGR 小鼠造模成功标准:新生鼠体重低于同胎龄同性别小鼠体重的第10 百分位点以下;②IUGR雌小鼠8 周成年后与同周龄雄鼠合笼,自受孕开始饲以标准繁殖饲料(蛋白含量为20%)。
1.2.2 小鼠胰岛的提取
NP、IP 孕鼠各3 只,用1%戊巴比妥钠麻醉后固定,剖开腹腔,分离穿刺胆总管,注入冷胶原酶消化、过细胞筛网,收集胰岛,放入Histopaque1077 中纯化。所有胰岛均在显微镜下手挑计数。
1.2.3 差异RNA检测分析
广州基迪奥生物科技有限公司使用RNA isolater Total RNA Extraction Reagent 裂解胰岛细胞,提取总RNA后使用Nano Drop超微分光光度仪进行RNA质检。
从总RNA 中去除核糖体RNA,以最大限度地保留所有编码RNA(coding RNA)和非编码RNA(non-coding RNA,ncRNA)。得到的RNA 随机打断成为短片段,再以片断化后的RNA 为模板,用六碱基随机引物(random hexamers)合成cDNA 第1 链;接着加入缓冲液、dNTPs(dUTP 代替dTTP)、RNase H和DNApolymeraseⅠ合成cDNA第2链,经过QiaQuick PCR 试剂盒纯化并加EB 缓冲液洗脱经末端修复、加碱基A,加测序接头,然后通过UNG(uracil-Nglycosylase)酶降解第2条链。用琼脂糖凝胶电泳进行片段大小选择,进行PCR扩增。最后建好的测序文库用Illumina-HiSeqTM4000进行测序。
对下机数据进行过滤得到clean data后,将reads用Tophat2(2.1.1)比对到小鼠参考基因组(ensemble 104)上,并利用Cufflinks重构转录本,得到已知转录本与新转录本。利用CPC、CNCI等软件对新转录本进行编码能力预测,得到新预测的lncRNA。然后分别对样本中的mRNA、lncRNA 进行表达量分析,并使用edgeR 包对组间lncRNA 和mRNA 进行差异分析。|log2FC|>1,P<0.05表示有差异。
利用lncRNA 与mRNA 表达量的Pearson 相关系数、RNAplex 等预测lncRNA 的靶mRNA,对靶基因进行基因本体论(gene ontology,GO)功能注释分析和京都基因与基因组百科全书数据库(Kyoto encyclopedia of genes and genomes,KEGG)通路富集分析。
1.2.4 实时荧光定量PCR
逆转录反应按照RT-PCR 试剂盒说明书进行。将AceQ qPCR SYBR Green Master Mix(High ROX premixed)应用于StepOne Plus 系统,进行qRT-PCR反应。反应体系20 μL:模板cDNA 1 μL,SYBR Green Master Mix 10 μL,上下游引物各0.4 μL,灭菌ddH2O 8.2 μL。引物序列如下,β-actin:上游5′-TGAGCTGCGTTTTACACCCT-3′,下游5′-TTTGGGGGATGTTTGCTCCA-3′;Neat1:上游5′-CAAGAAACAGCAACACCAGAAG-3′,下游5′-TAAGGTCCCCATTCAAGTCAGT-3′;FTX:上游5′-AAGATCTCCGCTGCCAGATG-3′,下游5′-CTGCTCCTGTGCCACGAATA-3′。反应条件:预变性95 ℃5 min;95 ℃10 s,60 ℃30 s,40 个循环。根据熔解曲线判断引物的特异性及扩增效率,采用2-ΔΔCt法对基因相对表达量进行计算和分析。
1.2.5 min6细胞的培养及不同糖浓度刺激
小鼠胰岛细胞系min6 细胞,其正常培养基(糖浓度25 mmol/L)的配制:84%的高糖DMEM(含4.5 g/L 葡萄糖),15%胎牛血清(FBS),1%双抗(100 U/mL 青霉素和100 μg/mL 链霉素),0.035%的2.5 mmol/L β-巯基乙醇。细胞置于37 ℃、含5%CO2培养箱中,每天更换新鲜培养基,融合度达80%左右进行传代。将高糖DMEM 替换成低糖DMEM(含葡萄糖1 g/L),余成分不变,此时低糖完全培养基的糖浓度为5.5 mmol/L,然后在10 mL低糖完全培养基中分别加入0.01、0.02、0.05 g 葡萄糖粉,得到11.1、16.7、33.3 mmol/L 不同糖浓度的培养基。将min6细胞接种于6孔培养板中,待细胞贴壁后,加入无糖低血清(0.25%)培养基培养6~8 h后弃去,分别加入不同糖浓度培养基2 mL,标记好糖浓度,放入37 ℃、含5%CO2培养箱中,24 h后收RNA。
1.3 统计学方法
使用Graphpad Prism 8.4.3 软件绘制实验结果图,基因表达量以均值±标准差()表示,两组间比较采用独立样本t检验,多组间比较采用单因素方差分析(one-way ANOVA)。P<0.05 为差异有统计学意义。
2 结果
2.1 lncRNA的组间差异
在NP、IP 中共鉴定出个30 426 个lncRNA 转录本,其中已知的转录本13 404(44%),新的转录本943 个。使用edgeR 软件对组间lncRNA 的表达量进行差异分析,发现在NP 和IP 中有差异表达的lncRNA 1 007个,其中483个上调,524个下调。图1示差异表达lncRNA 的火山图和热图。表1 列举了10个高差异表达的lncRNA。

表1 差异表达的lncRNATable1 Differentially expressed lncRNAs
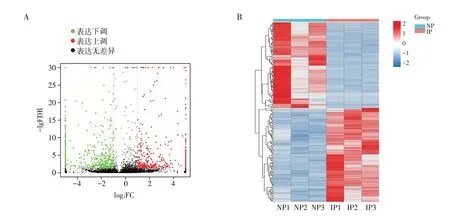
图1 正常孕鼠和IUGR孕鼠差异表达的lncRNAFigure 1 Differentially expressed lncRNAs between normal pregnant and IUGR pregnant mice
2.2 mRNA分析
2.2.1 mRNA的组间差异
在NP、IP 中共鉴定出66 652 个mRNA 转录本,其中已知的转录本43 034个(64.57%),新的转录本10 018个。使用edgeR 软件对组间mRNA 的表达量进行差异分析,发现在NP 和IP 中有差异表达的mRNA 50 个,其中22 个上调,28 个下调。图2 示差异表达的mRNA的火山图和热图。表2列举了10个高差异表达的mRNA。
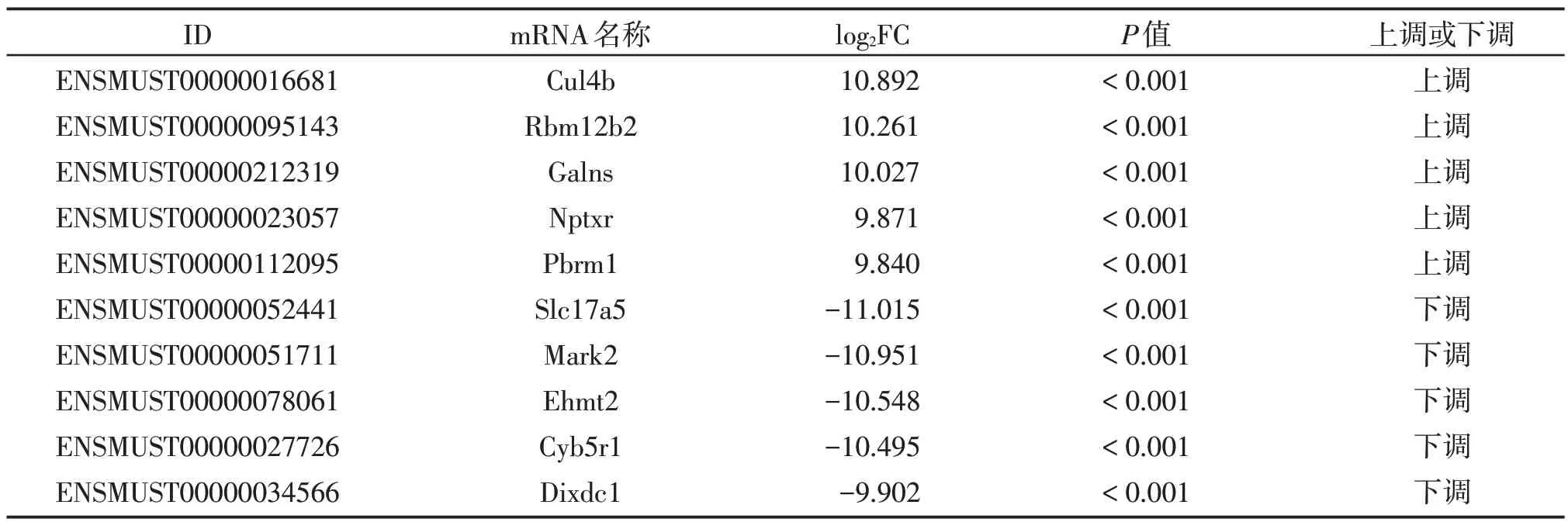
表2 差异表达的mRNATable 2 Differentially expressed mRNAs
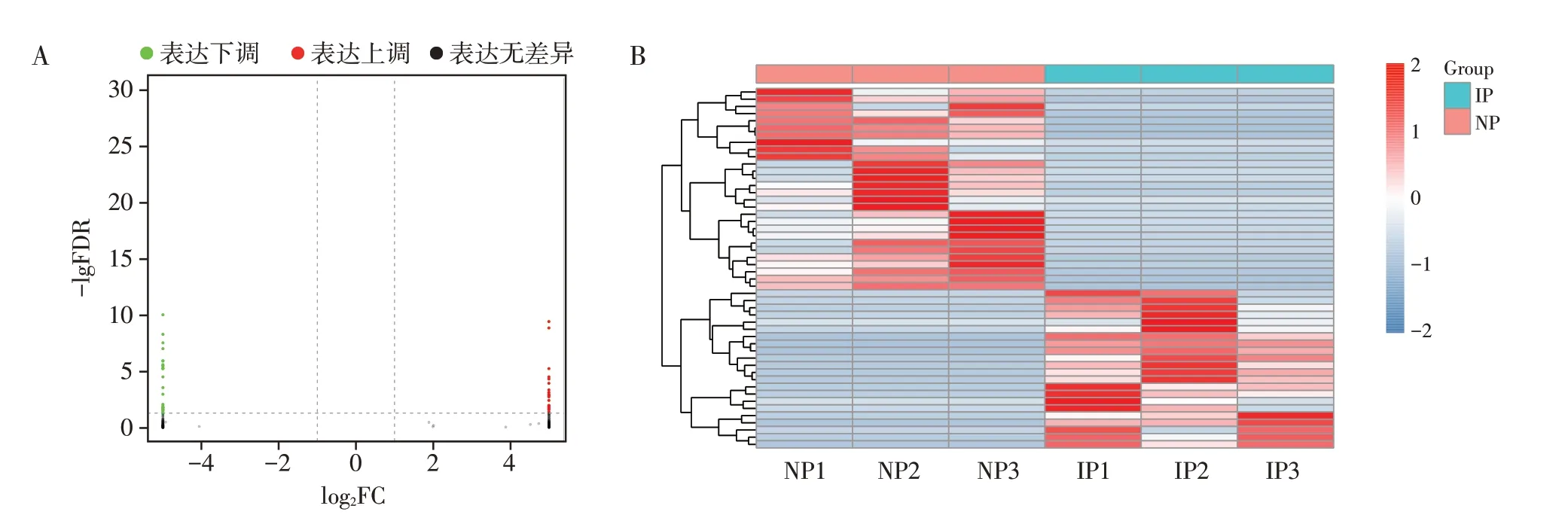
图2 正常孕鼠和IUGR孕鼠差异表达的mRNAFigure 2 Differentially expressed mRNAs between normal pregnant and IUGR pregnant mice
2.2.2 差异mRNA的GO/KEGG通路富集分析
图3显示了差异mRNA 的KEGG 通路富集分析和GO功能注释分析。KEGG信号通路富集分析中,按基因数量从多到少排序,前5 位分别是代谢通路(metabolic pathways)(4个基因)、PI3K-Akt信号通路(PI3K-Akt signaling pathway)(3 个基因)、内质网中的蛋白质加工(protein processing in endoplasmic reticulum)(3 个基因)、集落黏附(focal adhesiom)(3个基因)、AMPK信号通路(AMPK signaling pathway)(2 个基因)(图3A)。GO 功能注释分析中绿色表示下调基因,红色表示上调基因,按照参与GO 富集条目下调基因的数量排序,生物学途径(biological process,BP)中前5 位是细胞进程(cellular process)(18个基因)、单有机体过程(single-origanism process)(16 个基因)、生物调节(biological regulation)(11 个基因)、代谢过程(metabolic process)(11个基因)、刺激反应(response to stimulus)(9 个基因);细胞组分(cellular component,CC)中前5 位 是细胞(cell)(22 个基因)、细胞部分(cell part)(22个基因)、细胞器(organelle)(21 个基因)、细胞器部分(organelle part)(13 个基因)、细胞膜(membrane)(12 个基因);分子功能(molecular function,MF)中前4 位是绑定(binding)(18 个基因)、催化活性(catalytic activity)(9 个基因)、转运活性(transporter activity)(4 个基因)、化学排斥物活性(chemorepellent activity)(1 个基因)(图3B)。

图3 正常孕鼠和IUGR孕鼠差异mRNA的KEGG和GO富集分析图Figure 3 The KEGG and GO enrichment analysis of differentially expressed mRNAs between normal and IUGR pregnant mice
2.3 lncRNA与mRNA的关联分析
lncRNA 参与调控许多转录后进程。对mRNA的调控方式包括:反义(antisense)作用、顺式(cis)作用、反式(trans)作用。
2.3.1 antisense作用
antisense 作用指一部分反义lncRNA 与正义的mRNA 结合而调控基因沉默、转录及mRNA 的稳定性。图4A列举了3个差异lncRNA 与其靶mRNA 的antisense 作用,antisense 作用具有高度的保守性和稳定性[6],其中lncRNA Ppp1ca被报道通过MAPK通路参与肿瘤的发生[7],其与Rad9A 的互作关系未被研究,lncRNA 1010001N08Rik、AC154652.2 未曾有报道。差异lncRNA antisense 作用的靶mRNA 的KEGG、GO 富集分析如图4所示,KEGG 信号通路分析中按富集基因数量排序,前5 位分别是metabolic pathways(511个基因)、癌症通路(pathway in cancer)(204 个基因)、MAPK 信号通路(MAPK signaling pathway)(167 个基因)、次生代谢物的生物合成(biosynthesis of secondary metabolites)(158个基因)、PI3K-Akt signaling pathway(157 个基因)(图4B)。GO 功能注释分析中按基因数目排序,BP 中前5 位是cellular process(4 050 个基因)、single-organism process(3 712个基因)、biological regulation(3 208个基因)、metabolic process(2 184 个基因)、response to stimulus(1 904 个基因);CC 中前5 位分别是cell(4 249 个基因)、cell part(4 224 个基因)、organelle(3 310 个基因)、membrane(2 520 个基因)、organelle part(2 128 个基因);MF 中前5 位分别是binding(4 499 个基因)、catalytic activity(1 943 个基因)、transporter activity(460 个基因)、分子功能调节(molecular function regulator)(437 个基因)、核酸结合转录因子活性(nucleic acid binding transcription factor activity)(377个基因)(图4C)。

图4 差异lncRNA与其靶mRNA的antisense作用及靶mRNA的KEGG和GO富集分析Figure 4 The antisense between differentially expressed lncRNA and its target mRNA,the KEGG and GO enrichment analysis of the target mRNA
2.3.2 cis作用
cis作用指同一染色体上lncRNA的功能与其邻近的蛋白编码基因相关,位于这些基因上游或下游10 kb的lncRNA可能在转录或转录后水平对基因的表达进行调控。图5A列举了3个差异lncRNA与其靶mRNA 的cis作用。cis调控主要依赖顺式作用元件进行,顺式作用元件是指存在于基因旁侧序列中能影响基因表达的序列,包括启动子、增强子、调控序列和可诱导元件等,它们参与核内基因表达的调控,顺式作用元件通常转录为非编码RNA,如lncRNA。图5A 中lncRNA Ppp1ca、1010001N08Rik、AC154652.2 通过cis 分别和Rad9A、Gata6、Slc37a1有互作关系,而图4A是通过antisense作用有互作关系,这是因为lncRNA 和mRNA 链较长,antisense 作用的位置和区域与cis 作用的位置区域有所不同。差异lncRNA cis 作用的靶mRNA 的KEGG、GO 富集分析如图5 所示,KEGG 信号通路富集分析中按照基因数量排序,前5 位分别是metabolic pathways(847 个基因)、pathways in cancer(284 个基因)、biosynthesis of secondary metabolites(254个基因)、胞吞(endocytosis)(229 个基因)、PI3K-Akt signaling pathway(219 个基因)(图5B)。GO 功能注释分析中按基因数目排序,BP 中前5 位是cellular process(6 308 个基因)、single-organism process(5 438 个基因)、biological regulation(4 618 个基因)、metabolic process(3 780个基因)、response to stimulus(2 718个基因);CC 中前5 位分别是cell(6 745 个基因)、cell part(6 430 个基因)、organelle(5 119 个基因)、membrane(3 610 个基因)、organelle part(3 459 个基因);MF 中前5 位分别是binding(6 813 个基因)、catalytic activity(3 102 个基因)、molecular function regulator(601 个基因)、transporter activity(555 个基因)、nucleic acid binding transcription factor activity(515个基因)(图5C)。

图5 差异性lncRNA与其靶mRNA的cis作用,差异性lncRNA cis作用的靶mRNA的KEGG和GO富集分析Figure 5 The cis between differentially expressed lncRNA and its target mRNA,the KEGG and GO enrichment analysis of differentially expressed lncRNA cis target mRNA
2.3.3 trans作用
trans 作用指lncRNA 的功能与编码基因的位置没有关系,而与其共表达的蛋白编码基因相关,可以通过样本间lncRNA 与蛋白编码基因的表达量相关性分析或共表达分析方法来预测其靶mRNA,lncRNA 在其转录或转录后水平对基因的表达进行调控。图6A 列举了3 个差异lncRNA 与其靶mRNA的trans 作用,两者可以位于不同的染色体上,内源性竞争RNA(ceRNA)就属于此类[8],lncRNA Ppp1ca可以竞争性地结合miR-125b从而阻止其诱导的tau磷酸化[9]。差异lncRNA trans 作用的靶mRNA 的KEGG、GO 富集分析如图6所示,KEGG 信号通路富集分析中按基因数量排序,前5 位分别是metabolic pathways(675 个基因)、pathways in cancer(241 个基因)、biosynthesis of secondary metabolites(183 个基因)、MAPK signaling pathway(180个基因)、PI3K-Akt signaling pathway(179 个基因)(图6B)。GO 功能注释分析中按基因数目排序,BP 中前5 位是cellular process(4 692 个基因)、single-organism process(4 239 个基因)、biological regulation(3 550 个基因)、metabolic process(2 473 个基因)、response to stimulus(2 138 个基因);CC 中前5 位分别是cell(4 487 个基因)、cell part(4 437 个基因)、organelle(3 100个基因)、membrane(3 071个基因)、细胞膜部分(membrane part)(2 641 个基因);MF 中前5 位分别是binding(5 144个基因)、catalytic activity(2 200个基因)、分子传感器活性(molecular transducer activity)(578个基因)、transporter activity(543个基因)、信号转导活性(signal transducer activity)(540 个基因)(图6C)。
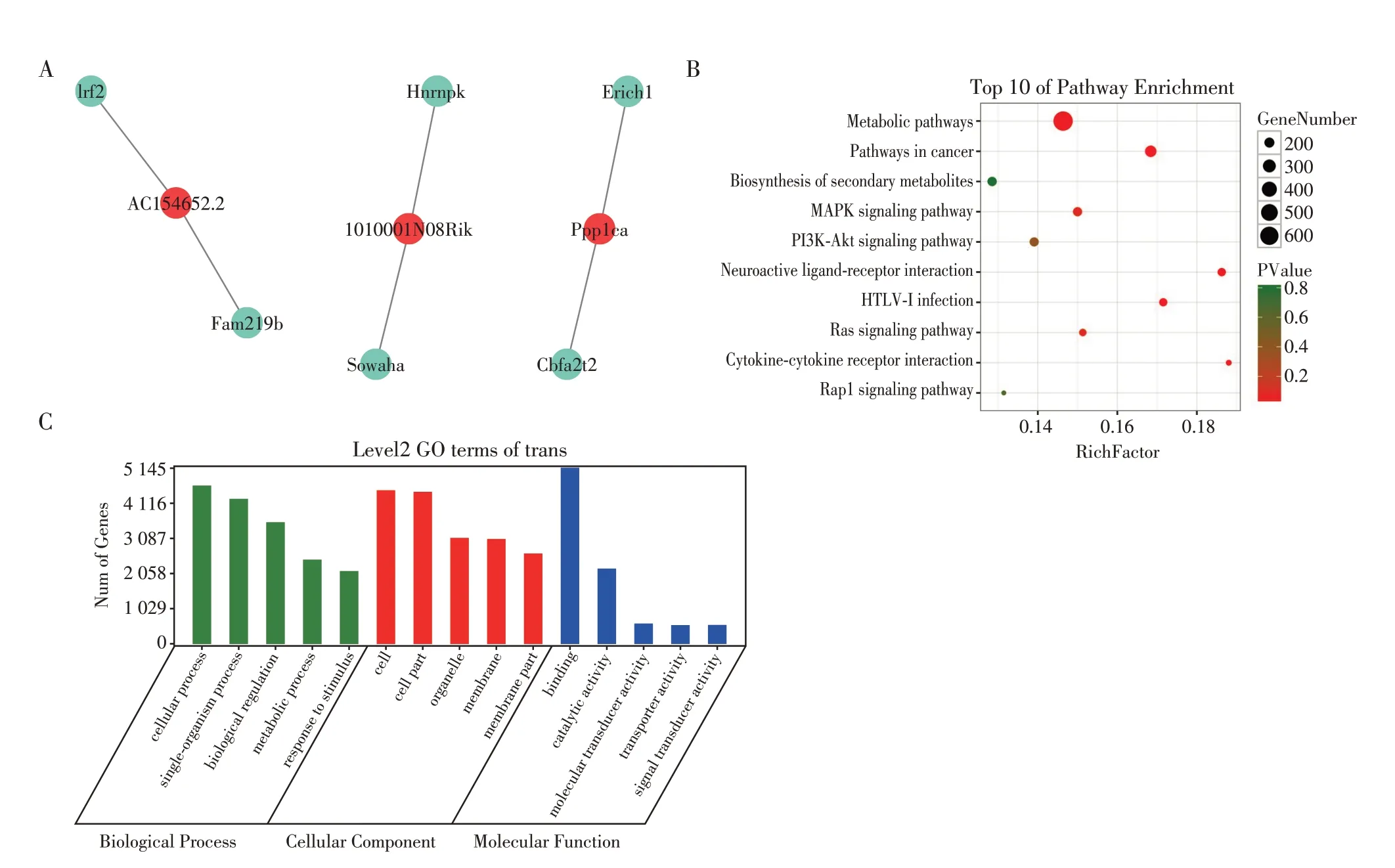
图6 差异性lncRNA与其靶mRNA的trans作用,差异性lncRNA trans作用的靶mRNA的KEGG和GO富集分析Figure 6 The trans between differentially expressed lncRNA and its target mRNA,the KEGG and GO enrichment analysis of differentially expressed lncRNA trans target mRNA
2.4 差异表达lncRNA FTX和Neat1的进一步分析
选取在NP和IP中高差异表达且可能与孕期胰岛发育、细胞增殖相关的lncRNA FTX 和Neat1进一步分析,qRT-PCR 示正常小鼠、IUGR 小鼠怀孕后FTX 的表达量分别较未孕时下降(P<0.001),而Neat1 的表达量分别较未孕时升高(P<0.01,图7)。FTX、Neat1在怀孕后的正常小鼠和IUGR 小鼠之间表达水平有显著差异(P<0.05,图7)。
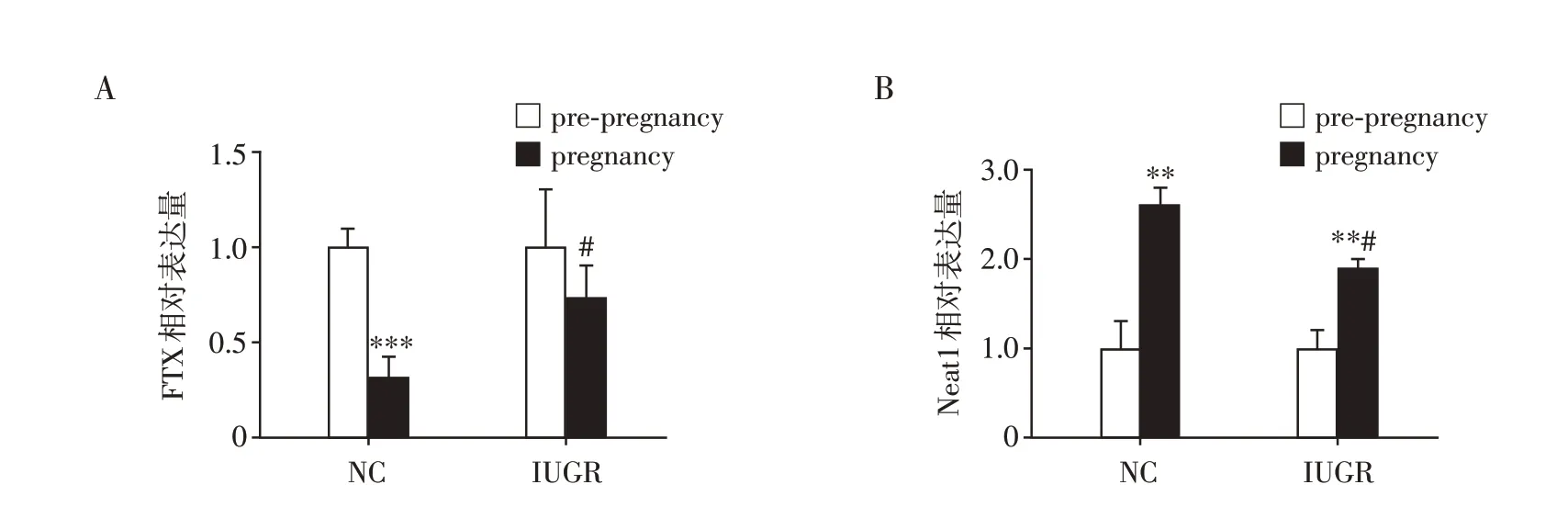
图7 lncRNA FTX(A)、Neat1(B)在正常小鼠、IUGR小鼠的未孕和孕期的表达Figure 7 Expression of lncRNA FTX(A)and Neat1(B)in pre-pregnancy and pregnancy of normal mice and IUGR mice
为明确lncRNA 的表达是否受糖浓度调节,本研究测定了min6 细胞在不同糖浓度刺激下FTX 和Neat1 的表达情况。FTX 在低糖浓度(5.5、11.1、16.7 mmol/L)、高糖浓度(33.3 mmol/L)下表达水平较正常糖浓度上调(P<0.001,图8A),而Neat1的表达相反(P<0.01,图8B)。
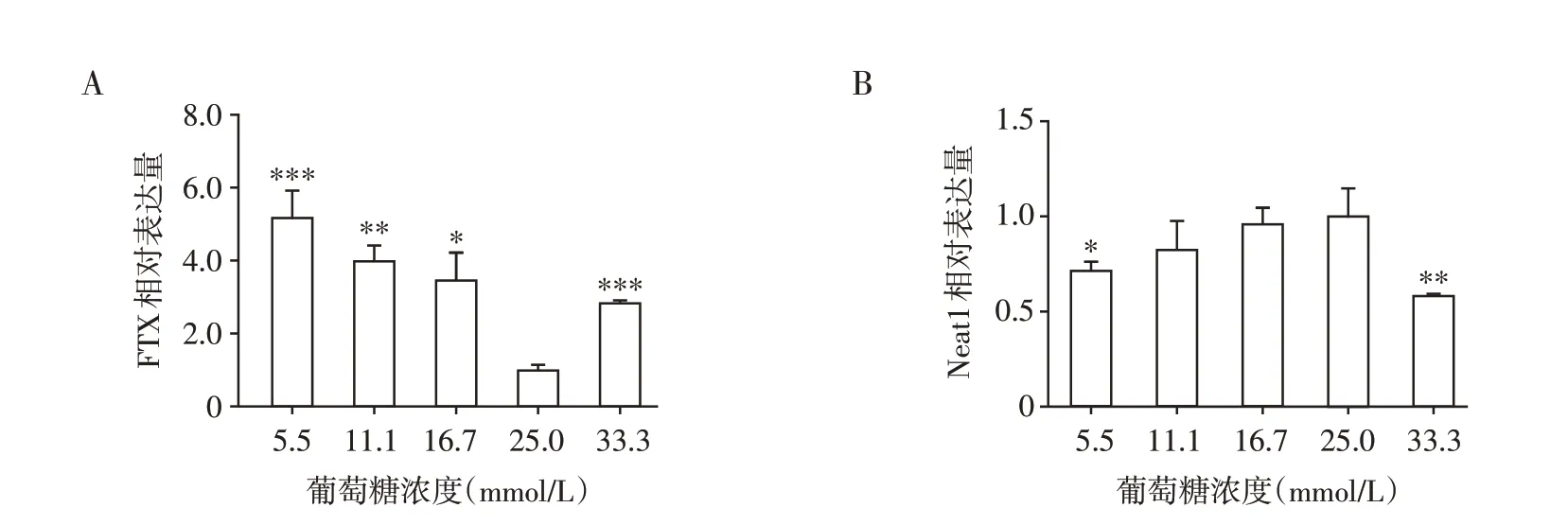
图8 不同糖浓度下min6 细胞中lncRNA FTX(A)、Neat1(B)的表达Figure 8 Expression of lncRNA FTX(A)and Neat1(B)in min6 cells under different glucose concentrations
进一步查找FTX 和Neat1 与其靶mRNA 的互作关系,图9A示FTX与其靶mRNA均为trans作用,图9B 示Neat1 与Frmd8(mRNA)为cis 作用,余为trans作用。
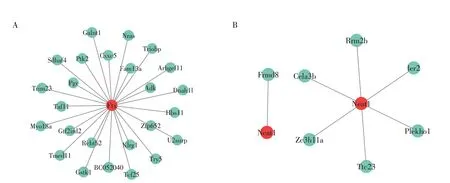
图9 lncRNA FTX(A)与靶mRNA的trans作用,lncRNA Neat1(B)与靶mRNA的cis作用(左)、trans作用(右)Figure 9 Trans-action of lncRNA FTX(A),cis(left)and trans(right)action of lncRNA Neat1(B)
3 讨论
妊娠是一个逐渐发生生理性胰岛素抵抗的过程,正常情况下胰岛β细胞通过代偿性增殖来维持孕期的糖代谢平衡,而IUGR 个体在胚胎期和新生儿早期胰岛发育和功能出现损害,成年后是否能应对妊娠这一代谢压力测试不得而知,目前关于IUGR 孕期的IUGR 小鼠研究未见报道。本研究首次对IUGR 孕鼠胰岛转录组进行高通量测序分析,为深入探究其发病机制提供理论基础。
越来越多的报道显示lncRNA 影响胰岛细胞的发育和功能、糖尿病的发生发展和IUGR 的发病过程。研究发现,lncRNA paupar 通过调控pax6 促进了胰岛α细胞的发育和功能[10];lncRNA hottip 通过调节p38-MAPK 通路改善糖尿病视网膜病变[11];本课题组前期研究表明,lncRNA TUG1 参与了IUGR致2 型糖尿病胰岛功能损伤的过程[1]。此外,lncRNA 在小鼠妊娠期不同阶段动态表达,其中lncRNA Gm16308(lnc03)促进小鼠妊娠期胰岛β细胞增殖[12],而lncRNA 表达谱的改变可诱发妊娠期糖尿病[13],Zhang 等[14]证明了lncRNA MALAT1 在妊娠期糖尿病孕妇组血清中的表达水平高于正常孕妇组。
本课题组前期实验结果显示:小鼠孕期存在胰岛增殖,但IUGR雌鼠孕期较正常小鼠孕期胰岛β细胞增殖能力减弱,胰岛大小及数量增加较少,出现糖耐量异常,为进一步探讨其发病原因,本研究对IUGR 孕鼠胰岛进行全转录组测序及分析,结果提示,差异lncRNA的靶mRNA、差异mRNA的GO分析中BP 集中于细胞及组织过程、生物调节、代谢及应激过程,CC集中在细胞及器官、胞膜等过程,MF集中于整合、催化活性、转运活性、分子功能调节等,这些基因均可对IUGR妊娠期胰岛功能产生影响,如能量代谢、β细胞周期调节及胰岛素胞吐过程等变化。KEGG 富集分析显示差异lncRNA 的靶mRNA、差异mRNA主要集中在代谢通路、癌症通路、次生代谢产物的生物合成、PI3K-AKT通路及MAPK通路。这表明IUGR 孕鼠和正常孕鼠在合成代谢、分解代谢以及癌症的发生发展方面存在差异。
PI3K/AKT、MAPK 通路在维持胰岛β细胞增殖和存活方面起着关键作用。文献报道,催乳素受体和PI3K(AKT、mTOR、p70S6K)通路、MAPK(MEK、ERK1/2)通路参与大鼠妊娠期胰岛质量的增加和葡萄糖敏感性的增强[15]。妊娠期胎盘生长因子可能刺激胰岛内皮细胞释放生长因子,激活β细胞中的PI3K/AKT 信号以增加其增殖[16]。表皮生长因子通过PI3K/AKT 和MAPK 通路激活存活蛋白survivin,抑制β细胞凋亡[17]。这些过程和通路异常可能导致IUGR小鼠孕期发生胰岛功能障碍。
本研究发现lncRNA Neat1、FTX 在小鼠怀孕后表达量改变,并可能分别通过MAPK 途径、PI3K/AKT途径影响小鼠孕期胰岛细胞增殖及功能,提示lncRNA 的表达在妊娠期这个特殊的时间段受到严格的调控。既往研究发现Neat1可以通过调节细胞的增殖及凋亡,导致疾病的发生,如肿瘤、白血病及糖尿病肾病[18]。研究证实,lncRNA Neat1 在宫内发育迟缓模型的绒毛滋养细胞中表达上调[19],提示Neat1 与特发性IUGR 胎儿的胎盘功能障碍有关,而Neat1 在IUGR 孕鼠胰岛发育、胰岛结构及功能维持中的作用及机制尚未见报道。lncRNA FTX参与多种疾病的发生、发展,如lncRNA FTX 通过调节miR-192-5p/EIF5A2 轴促进直肠癌的进展[20],通过海绵化miRNA-22 调节pten/PI3K/AKT 信号通路而减少新生小鼠心肌细胞肥大[21],但FTX 在胰岛中尚未被研究。本研究在IUGR 孕鼠和正常孕鼠胰岛中检测lncRNA Neat1和FTX的表达水平,差异有统计学意义。不同糖浓度刺激min6 细胞后,发现二者的表达受糖浓度影响,lncRNA这一特点可能在IUGR孕鼠发生糖耐量异常过程中发挥作用。测序结果还发现lncRNA FTX 与其靶mRNA 均为trans作用,lncRNA Neat1 与Frmd8(mRNA)为cis 作用,余为trans 作用,结合lncRNA 和mRNA 的互作分析(图4A、5A、6A)可见,同一种lncRNA 可以通过不同的互作方式调控不同mRNA 的表达,而同一种lncRNA 和同一种mRNA 之间可以存在不同的互作方式,这是因为不同的作用方式在lncRNA和mRNA核苷酸链上所在的位置和区域不同。
文献表明,lncRNA 可能通过充当miRNA 海绵并调节mRNA 转录后沉默而起到基因调节剂的作用[3],本课题组正在实验验证测序结果中的互作关系,若干扰或过表达lncRNA 后,其靶mRNA 的表达量随之发生相应的变化,接下来再深入探讨其中是否涉及lncRNA-miRNA-mRNA这一“海绵”作用。
本研究通过对IUGR孕鼠胰岛RNA进行高通量测序,筛选出了IUGR 孕鼠和正常孕鼠差异表达的lncRNA 及mRNA,并对差异lncRNA 的靶mRNA 进行GO、KEGG分析,为研究IUGR小鼠妊娠期发生胰岛功能障碍提供了理论基础。在此基础上,进一步深入探究差异表达的lncRNA 对IUGR 小鼠妊娠期胰岛功能的具体调控作用及其机制,以期早期防控糖尿病,降低糖尿病的发病率。
猜你喜欢
新民周刊(2022年27期)2022-08-01 07:04:49
传染病信息(2021年6期)2021-02-12 01:52:58
生命科学研究(2018年1期)2018-05-29 01:13:01
中国实验动物学报(2016年2期)2016-07-15 01:46:36
生物医学工程学进展(2015年1期)2015-02-28 14:53:42
肝胆胰外科杂志(2015年1期)2015-02-27 11:11:34
化学工业与工程(2015年1期)2015-02-10 03:01:41
中国病理生理杂志(2015年10期)2015-01-26 04:36:31
中国医学科学院学报(2014年6期)2014-03-11 20:26:19
生命科学研究(2012年3期)2012-04-29 00:44:03
