
松材线虫入侵对马尾松林群落物种多样性的影响
2022-07-16 07:27龚笑飞李振飞李大标韦博良胡广刘金亮吴初平江波于明坚余水生
浙江林业科技 2022年4期
龚笑飞,李振飞,李大标,韦博良,胡广,刘金亮,吴初平,江波,于明坚,余水生
(1.浙江省遂昌县生态林业发展中心,浙江 遂昌 323300;2.浙江大学 生命科学学院,浙江 杭州 310058;3.浙江理工大学 建筑工程学院,浙江 杭州 310018;4.温州大学生命与环境科学学院,浙江 温州 325035;5.浙江省林业科学研究院,浙江 杭州 310023)
松材线虫Bursaphelenchus xylophilus源自北美,可寄生于多种松属Pinus植物体内引发松树萎蔫病,导致其迅速死亡[1]。1982 年,我国在南京中山陵首次发现松材线虫病,30 多年来已有18 个省份发生松材线虫病,感染松林面积达649 333 hm2,造成了上千亿元的经济损失[2]。松材线虫入侵马尾松Pinus massoniana林后,群落内的优势种马尾松逐渐染病死亡,释放出大量的空间与资源,进而影响群落的物种组成和演替方向[3]。我国亚热带地区以马尾松为优势种的先锋群落经典的演替序列为马尾松林(针叶林)—马尾松针阔叶混交林—常绿阔叶林[4]。松林感染松材线虫病之后,恢复阶段的演替方向会受到染病前松林的物种组成和群落结构以及干扰程度的影响[5]。物种多样性低、立地条件差的松林染病后,其生态系统功能被迅速破坏,群落向顶级群落的演替速度更加缓慢[6];物种多样性高、立地条件好的松林在染病之后,能够维持一定的功能,并在功能发生改变后较快恢复到原有水平[7]。通过对染病马尾松林的研究发现,马尾松林朝着常绿阔叶林和针阔混交林2 个方向发展[8-11]。
物种多样性是对一个群落的结构和功能复杂性的度量,代表着物种演化的空间范围和对特定环境的生态适应,对其进行研究可以更好地认识群落的组成、变化和发展趋势[12]。生长型是植物长期进化与演变过程中对环境适应的结果,不同生长型植物的环境适应策略不同[13]。龙翠玲等[14]通过对不同年龄阶段林隙的植物物种多样性研究发现,不同生长型植物在不同林隙阶段中的优势程度不同。区余端等[15]通过对冰雪灾害后常绿阔叶林中地被植物连续三年的监测发现,不同生长型植物在各年度的组成和分布都存在显著差异。群落中木本植物的总体动态只能揭示出一般趋势,会掩盖某些特殊性,按照生长型分类对群落进行研究,有助于了解不同生长型物种对环境的适应,对研究植物群落的发生、发展以及演替规律都有重要意义[16]。
为了解不同染病阶段马尾松林群落与该地区地带性植被常绿阔叶林结构组成的差异,本研究以浙江省遂昌县松材线虫侵染4 年和8 年的马尾松林群落为研究对象,并与未染病的马尾松林和常绿阔叶林作比较,探讨松材线虫入侵马尾松林后木本植物及不同生长型植物物种的多样性变化和松材线虫入侵后马尾松林的演替趋势,旨在从植被变化角度入手,研究松材线虫入侵马尾松林后引起的群落结构变化和演替特征,为松材线虫入侵马尾松林后生态系统恢复和管理提供基础数据和理论依据。
1 研究区概况和研究方法
1.1 研究区概况
研究区位于浙江省遂昌县,地理坐标为28°13′~ 28°49′ N,118°41′~ 119°30′ E,属中亚热带湿润季风区,年平均气温为16.8℃,无霜期为251 d,年日照时数为1 755 h,年均降水量为1 510 mm。遂昌县森林资源丰富,森林覆盖率达83.47%,其中松林面积有4.3 万hm2,占林地总面积的19.43%[17-18]。松木以马尾松为主,容易感染松材线虫病。2011 年,遂昌县在妙高街道金岸村首次发现松材线虫感染病例,至2017 年,受灾面积已达3 801.5 hm2[18]。
1.2 样地设置与群落调查
根据遂昌县2010 年森林资源二类调查数据、2020 年森林资源监测数据和松材线虫病疫情普查数据,本研究在遂昌县内选取未染病的马尾松林(UN)、染病(本文中指感染松材线虫病)4 年(2016 年染病)马尾松林(4Y)、染病8 年(2012 年染病)马尾松林(8Y)和常绿阔叶林(EV),所选的马尾松林群落在松材线虫入侵之前,其林分结构、林龄和生境相似。在每种群落类型中分别设置3 个30 m×30 m 的固定样地,且每2 块样地之间的直线距离不少于300 m。对样地内所有胸径(DBH)≥1 cm 的木本植物个体进行每木调查,记录每株植物的物种名称,测定胸径、树高等参数,并根据《中国植物志》以及《浙江植物志》将木本植物分为乔木物种和灌木物种[13]。样地的基本情况见表1。
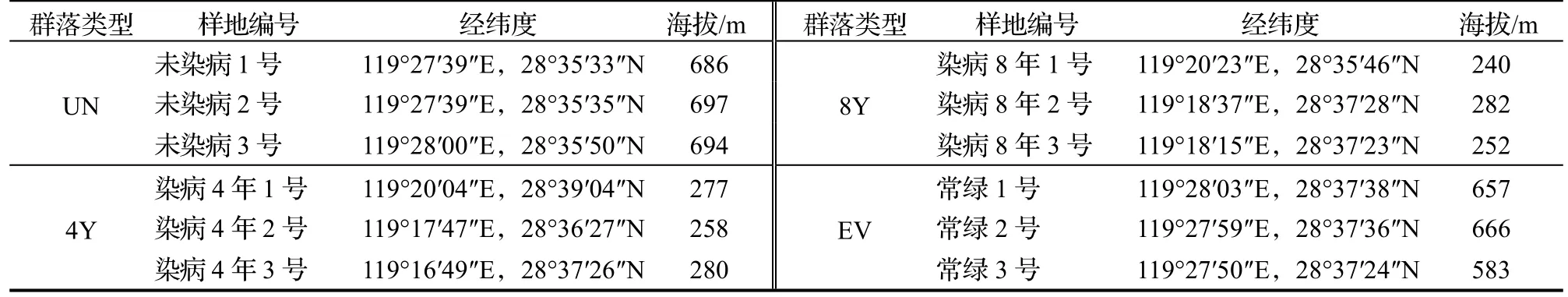
表1 样地的基本情况Table 1 Information of sample plots
1.3 植物群落的径级结构
参考相关径级划分方法[19],并根据调查林分的实际情况,把样地中的林木径级划分为6 个等级:Ⅰ级(1≤DBH<5 cm)、Ⅱ级(5≤DBH<10 cm)、Ⅲ级(10≤DBH<15 cm)、Ⅳ级(15≤DBH<20 cm)、Ⅴ级(20≤DBH<30 cm)、Ⅵ级(DBH≥30 cm),统计各样地中各径级物种的个体多度,计算不同径级植物的多度。
1.4 重要值计算
重要值是用来表示某个种在群落中优势度的综合数量指标,以重要值为测度指标可以避免单纯因植物个体大小差异而导致的某些植物种类在群落中作用的高估[20]。其计算公式为:
重要值=(相对密度+相对频度+相对显著度)/3
相对频度=某一物种的频度/全部物种的频度之和×100%
相对显著度=某一物种的胸高断面积之和/全部物种的胸高断面积之和×100%
1.5 α多样性指数
本研究分别计算了群落内所有木本植物乔木物种和灌木物种的物种α多样性。利用Shannon-Wiener 多样性指数(H′)、Simpson 多样性指数(DS)分别计算物种α多样性,采用Pielou 均匀度指数(J)衡量植被群落物种的分布均匀程度[21-22]。各计算公式如下:
Shannon-Wiener 多样性指数(H′):

Simpson 多样性指数(DS):

Pielou 均匀度指数(J):
软件系统设计也是整个监控系统设计中较为重要的一项系统设计,在其系统的设计中,主要针对系统设计监控成像系统的处理,通过软件设计,将对应的控制程序输入到监控系统设计的内部控制系统中,通过这种系统监督控制处理,能够发挥出整个系统监控工作开展的保障性工作。比如对于监控系统应用界面的设计,以及对应的监控系统启动程序设计等都属于软件系统设计中的构成因素,整个软件系统设计编写中,以国际编程计算机软件信息处理标准为依据。

式中,S为样地内物种的数量;Pi为物种i的重要值。
1.6 β多样性指数
利用Jaccard 指数(qmn)反映群落间的物种组成的相似性,其计算公式[23]为:

式中,a为群落m和n中同时出现的物种数目,b为群落m中出现的物种数目,c为群落n中出现的物种数目。
1.7 数据分析
不同径级物种多度的统计和物种β多样性指数的计算借助Excel 完成,重要值及α多样性指数计算由R 4.0.0的Vegan 软件包完成[24]。为检验不同群落多样性指数的差异,采用R 4.0.0 的Agricolae 软件包进行非参数Kruskal-Wallis 检验,利用Origin 2019 完成有关绘图工作。
2 结果与分析
2.1 不同群落物种丰富度差异
本研究共调查到12 个样地中DBH≥1 cm 的木本植物共有5 179 株,共计116 种,分属41 科、76 属。其中,未染病的马尾松林木本植物共计47 种1 368 株,染病4 年的马尾松林木本植物共计36 种1 231 株,染病8 年马尾松林木本植物共计57 种1 132 株,常绿阔叶林木本植物共计64种1 448 株(表2)。木本植物科、属和种的数量在染病4年马尾松林中最低,在常绿阔叶林中均为最高。

表2 各群落物种组成概况Table 2 Species composition in each community
2.2 不同群落径级结构
所调查的4 个群落的径级结构如图1。由图1 可知,4 个群落的径级分布整体上均呈现为倒“J”形,以小径级植株为主,大径级植株较少,群落处于初级阶段,整体更新良好。在未染病马尾松林、染病4 年马尾松林、染病8 年马尾松林和常绿阔叶林中,径级Ⅰ级的木本植物株数分别占所在林分DBH≥1 cm 的木本植物总株数的66.16%、59.3%、64.13%、58.77%。
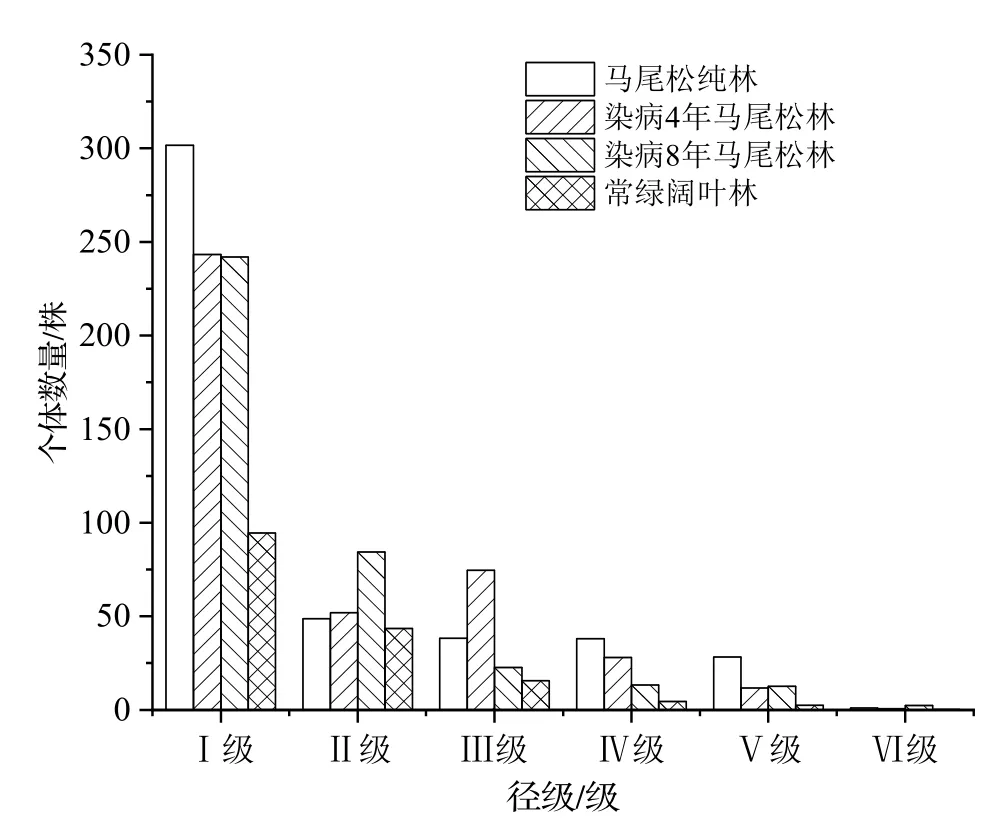
图1 各群落木本植物的径级分布Figure 1 Diameter class distribution of woody plant in each community
各群落中针、阔叶树种的数量分布如表3。由表3可知,在未染病马尾松林、染病4 年马尾松林、染病8 年马尾松林和常绿阔叶林中阔叶树种的占比分别为64.99%、66.61%、90.64%和98.83%,其中在染病8年马尾松林中阔叶树种的比例仅次于在常绿阔叶林的。径级Ⅰ级木本植物数量在各群落木本植物中均占有较大比例,在未染病马尾松林、染病4 年马尾松林、染病8 年马尾松林和常绿阔叶林中,径级Ⅰ级木本植物中阔叶树种的占比分别为93.15%、98.49%、99.31%和99.76%,在2 种染病马尾松林中的阔叶树种比率均高于未染病马尾松林中的。
2.3 物种α多样性
2.3.1 木本植物的物种α多样性 各群落木本植物物种α多样性如图2。由图2 可知,马尾松林染病之后,从未染病到染病4 年,Shannon-Wiener 指数和Simpson 指数降低,但到染病8 年时,Shannon-Wiener 指数和Simpson指数有所增加,并在常绿阔叶林中达到最高。与其他指数不同,Pielou 指数呈现为持续增加的趋势。方差分析结果表明,常绿阔叶林和染病8 年马尾松林木本植物的Shannon-Wiener 指数、Simpson 指数、Pielou 指数均显著高于染病4 年马尾松林(P<0.05);染病8 年马尾松林的Pielou 指数显著高于未染病马尾松林(P<0.05)。
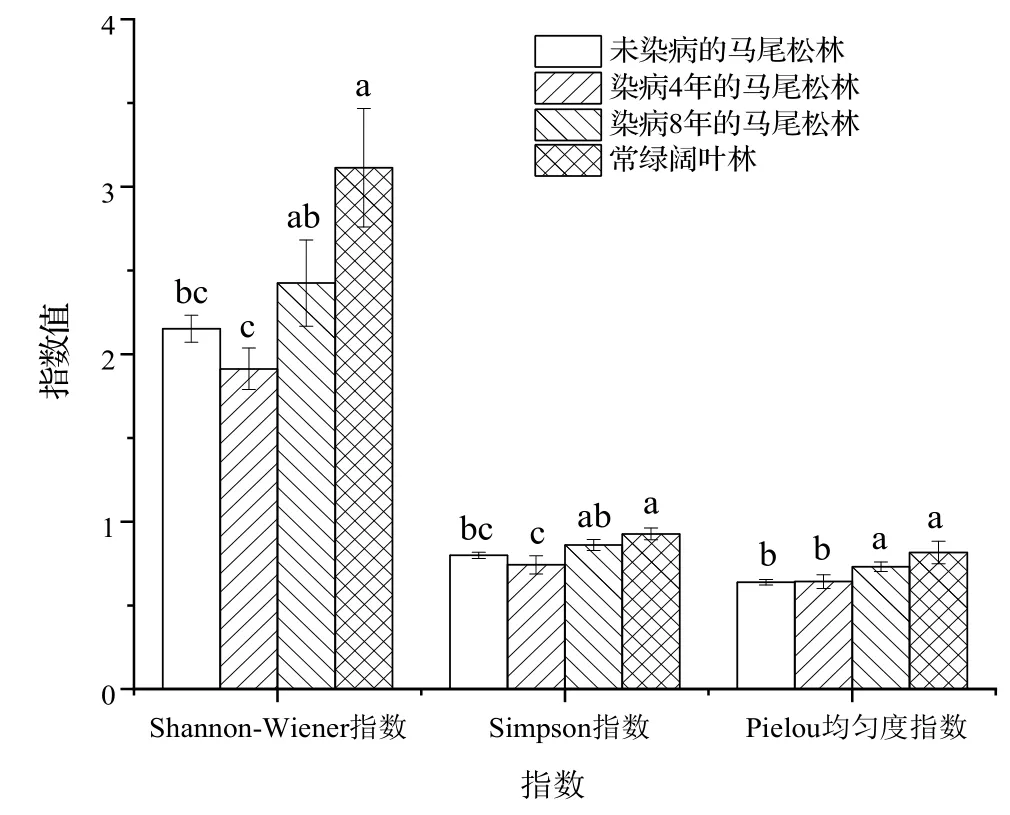
图2 各群落木本植物的物种α多样性Figure 2 α diversity of woody plant species in each community
2.3.2 乔木的物种α多样性 各群落乔木的物种α多样性如图3。由图3 可知,Shannon-Wiener 指数、Simpson指数、Pielou 指数均表现为常绿阔叶林>染病8 年马尾松林>未染病马尾松林>染病4 年马尾松林。方差分析结果表明,常绿阔叶林和染病8 年马尾松林乔木物种的Shannon-Wiener 指数、Simpson 指数、Pielou 指数均显著高于染病4 年马尾松林(P<0.05),常绿阔叶林乔木物种的Shannon-Wiener 指数、Simpson 指数和Pielou 指数均显著高于未染病马尾松林(P<0.05)。

图3 各群落乔木的物种α 多样性Figure 3 α diversity of tree species in each community
2.3.3 灌木的物种α 多样性 各群落灌木的物种α多样性如图 4。由图 4 可知,随着染病时间的增加,Shannon-Wiener 指数和Simpson 指数均呈现为先降低后增加的趋势。Shannon-Wiener 指数和Simpson 指数均在染病8 年时最低。Pielou 指数表现为常绿阔叶林>染病4年马尾松林>未染病马尾松林>染病8 年马尾松林。方差分析结果表明,常绿阔叶林灌木物种的Shannon-Wiener指数和Simpson 指数均显著高于染病4 年和染病8 年马尾松林(P<0.05),常绿阔叶林的Pielou 指数显著高于病8 年马尾松林(P<0.05)。

图4 各群落灌木的物种α多样性Figure 4 α diversity of shrub species in each community
2.4 群落β多样性
不同群落之间的相似性见表4。染病4 年马尾松林与未染病马尾松林的相似性系数最高(0.349),染病4 年马尾松林与染病8 年马尾松林的相似性系数次之(0.348),染病4 年马尾松林与常绿阔叶林的相似性系数最低(0.232)。随着染病时间的增加,不同染病阶段的马尾松林与未染病马尾松林之间的相似性系数逐渐降低,与常绿阔叶林群落之间的相似性系数逐渐增加,染病8 年马尾松林与常绿阔叶林之间的相似性系数(0.263)高于未染病马尾松林与常绿阔叶林的相似性系数(0.258)。
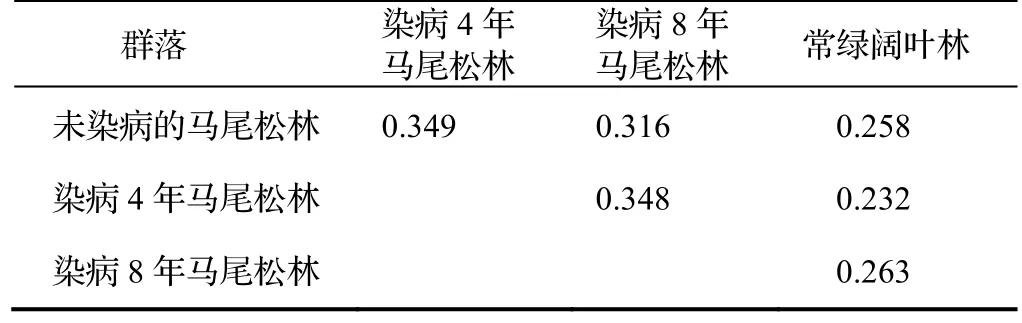
表4 各群落的Jaccard 相似性系数Table 4 Coefficient of Jaccard similarity of each community
3 讨论与结论
3.1 松材线虫入侵对群落结构的影响
4 种群落的径级结构总体均为反“J”形,表明群落内有丰富的幼树储备,能够实现自我更新[25]。在未染病马尾松林中阔叶树种的占比最低,在染病8 年马尾松林中阔叶树种的占比仅次于常绿阔叶林中的,且在染病马尾松林中径级Ⅰ级的小树中阔叶树种的比率均高于未染病马尾松林中的,这表明松材线虫感染导致松树大量死亡后,小径级的物种组成最先发生改变[26],阔叶树种比例有增加的趋势。
3.2 松材线虫入侵对物种α多样性的影响
3.2.1 对群落木本植物物种α 多样性的影响 马尾松林感染松材线虫病后,群落木本植物的多样性指数先降低后增加。在松材线虫病爆发后,染病马尾松死亡形成林窗,生境短时间内发生剧烈变化,不利于一些物种的生存,而且土壤干燥不利于种子的萌发[27],造成多样性的下降,因此,染病4 年马尾松林的木本植物物种多样性降到最低。马尾松的数量下降后,样地环境受松材线虫病的干扰也会减小。林内环境随着林窗的闭合而逐渐改善,因马尾松死亡释放的大量空间与资源也会促进其他物种的发展,使得多样性明显增加[7,28]。染病8 年马尾松林的Simpson 指数已经接近于常绿阔叶林,与未染病马尾松林相比,Simpson 指数升高 说明群落多样性升高,优势种的优势度降低[22];Pielou 指数增加,表明群落的均匀度在增加,即各物种个体数量的差异变小,一定程度上反应群落的优势种(马尾松)的优势度有所减弱。此现象可能符合中度干扰假说,这种程度的干扰既利于新物种的定植和种群扩大,又不会因为过于激烈使得物种无法生存[29]。
3.2.2 对不同生长型植物物种α 多样性的影响 马尾松群落中不同生长型物种的多样性指数对松材线虫入侵的响应不同,乔灌木物种的多样性指数在染病4 年马尾松林中略低于未染病马尾松林。马尾松死亡后,一些不能适应强烈光环境的灌木物种消失,能适应新环境的灌木物种快速占据优势,数量急剧增加[30],使得染病4 年马尾松林群落灌木物种多样性指数降低。林窗有利于林下植物的定居,能适应环境的优势物种生长加快,但是随着优势物种的生长和繁衍占据大量的空间资源,群落的物种数量可能会降低[27]。灌木和草本植物的快速生长繁殖,占据了林窗空间的底层生态位,会抑制乔木物种的定殖和生长[27],极端的生境条件和竞争因素共同导致染病4 年马尾松林群落乔木物种多样性指数和均匀度指数降低。
与染病4 年马尾松林相比,在染病8 年马尾松林中乔木物种的多样性指数均显著增加。马尾松林的林内环境随着林窗的发育逐渐恢复,利于种子的萌发和幼苗更新,乔木生长起来占据了林窗上层空间[31-32],多样性指数已经恢复到高于未染病马尾松林的水平,并且与常绿阔叶林的接近。与染病4 年马尾松林相比,染病8 年马尾松林中灌木物种的多样性指数并无明显变化,这是因为马尾松死亡初期大青Clerodendrum cyrtophyllum、檵木Loropetalum chinense等生长速度较快的灌木物种占据了林窗下层生态空间,随着林内环境的改善,栀子Gardenia jasminoides、冬青Ilex c hinensis等一些耐阴物种虽然已经进入群落,却还未开始大量繁殖,灌木群落处于森林空间的底层生态位,发育过程中受到冠层植物的遮蔽,群落的反应更为复杂[33]。
3.3 松材线虫入侵对群落β多样性的影响
随着染病时间的增加,染病不同阶段马尾松林与未染病马尾松林的Jaccard 相似性系数逐渐降低,而与常绿阔叶林的群落相似性系数增加。这是由于松材线虫侵入马尾松林群落后,林冠层马尾松死亡,造成林窗开放,改变了林内资源和其空间分布,导致阔叶树种进入群落并迅速生长起来,引起森林群落结构变化[28]。本地区的顶级群落是常绿阔叶林。正常情况下,马尾松林会逐渐向针阔叶混交林以及常绿阔叶林演替[8,34]。但马尾松寿命可达170 年,自然演替需要很长的时间[35-36]。而在松材线虫入侵后,短短8 年时间,染病马尾松林与常绿阔叶林的相似性便高于未染病的马尾松林与常绿阔叶林的相似性,这表明干扰在一定程度上加速了演替的进行。
因此,松材线虫入侵或许只是作为原有松林群落自然演替或更新过程中的一个强有力的外来选择因子,虽然其本身对马尾松林造成了破坏,但从演替角度来看,这反而能够促进群落的自然更新、自然演替,向生态效益更高的阔叶林发展。对于染病马尾松林的管理,建议在马尾松死亡后的林窗内补种当地阔叶树种以加快形成多种植物组成、多种层次的顶极群落或稳定性较强的混交次生林。
猜你喜欢
广西林业科学(2022年4期)2022-11-07
林业调查规划(2022年3期)2022-06-13
园艺与种苗(2021年3期)2021-05-26
绿色中国(2019年18期)2020-01-04
农家科技下旬刊(2017年8期)2017-11-13
科学种养(2017年6期)2017-06-13
绿色科技(2016年23期)2017-03-15
科技创新导报(2016年21期)2016-12-17
湖北农业科学(2016年18期)2016-12-08
现代园艺(2016年17期)2016-10-17
