
植食性昆虫嗅觉识别植物挥发物机制的研究进展
2022-07-14 02:09吕金言孟昭军
中国农学通报 2022年15期
吕金言,孟昭军
(东北林业大学林学院森林生态系统可持续经营教育部重点实验室,哈尔滨 150040)
0 引言
自然界中,植食性昆虫通常利用寄主植物释放的化学物质来确定自己的飞行方向,准确找到寄主,进行交配、产卵、躲避天敌等生理活动[1]。其中,昆虫的嗅觉、触觉、视觉和味觉都起到重要作用[2]。大多数植食性昆虫主要依靠嗅觉来定位和区别寄主及非寄主植物,大量对植物挥发性化学信号高度敏感的嗅觉感器参与昆虫的觅食等行为[3]。植物挥发性有机化合物(volatile organic chemicals,VOCs)是昆虫进行寄主搜索及定位生境的重要化学信息纽带,在植物与昆虫相互依存的关系中发挥关键作用[4-5]。有学者将信息论结合生态学与进化理论,说明VOCs可通过植物与草食动物之间的信息协调形成稳定的信息结构[6]。研究植食性昆虫嗅觉对寄主VOCs的感受机制,有助于揭示昆虫与寄主植物协同进化关系,为筛选植物抗性品种及开发害虫绿色防控技术提供理论依据。
1 植物挥发物种类及其特性
植物是生态系统食物链中为其他各营养级提供能量的生产者,为克服无法移动的限制并满足自身繁殖需要,植物与昆虫经过长期的协同进化形成了复杂紧密的关系,植物为昆虫提供食物及生活场所,昆虫帮助植物进行传粉、防御等[7-8]。据估计,世界上至少有2万种植物和昆虫有着密切的相互依存关系,这些植物在生长发育过程中产生约5万~10万种次生化合物,这些化合物中的大部分对植物正常生长发育影响不大,但研究表明它们在植物与昆虫的相互作用中非常重要[9]。植物挥发性次生物质是一些分子量在100~200,由植物叶、花、芽表面散发的短链碳氢化合物,包括烃类、醇类、醛类、萜类化合物等[10](表1);其组分易随植物的年龄、生理状态、空间分布、外界温度、土壤、光照等因素的影响而改变,从而影响植食性昆虫识别其寄主植物[11]。植物挥发物不仅能显示植物的身份特征,还能反映出植物在不同环境条件下、不同生长发育时期,甚至不同器官的生理状态[12]。
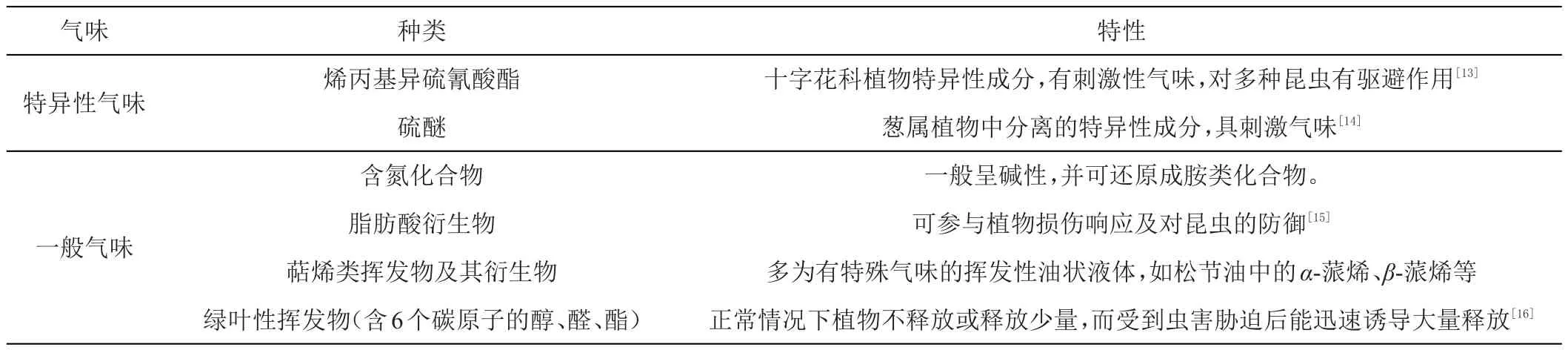
表1 特异性气味和一般气味种类及特性
VOCs能直接影响昆虫求偶、繁殖等行为[17],在植物与昆虫的协同进化中起关键作用,是植物同植食性昆虫建立二级营养关系的信息化合物[18]。绝大多数昆虫,尤其是夜间活动的植食性昆虫通常依靠感知VOCs来发现、定位和选择寄主或规避非寄主[19],昆虫可利用寄主植物释放的化学信号确定飞行方向;而植物也可产生大量的次生代谢物质来抵御昆虫的取食,彼此间形成了多种互相适应和防御机制,昆虫的行为与植物挥发物在相互作用下不断进化[20]。
2 植食性昆虫对植物挥发物的响应
2.1 单食性昆虫与多食性昆虫对植物挥发物的识别
近年来,国内外已有多位学者探究寄主植物挥发物与昆虫行为关系。单食性昆虫可通过嗅觉识别特异性挥发物来确定其寄主,如专食性莲草直胸跳甲(Agasicles hygrophila)[21]对两种特异性寄主植物挥发物D、Z以99:1比例的混合物反应较强,增加挥发物剂量会使雌虫的产卵量显著增大。多食性昆虫则利用挥发物各组分含量的不同区分寄主植物和非寄主植物[22],如(Z)-3-己烯-1-醇能对未交配云斑天牛(Batocera lineolata)雌、雄成虫产生明显的行为引诱作用,而芳樟醇对其有显著的驱避作用,从而完成寄主的选择[23];此外,云杉八齿小蠹(Ips typographus)[24]、东方云杉卷叶蛾(Choristoneura fumiferana)[25]、星 天 牛 (Anoplophora chinensis)[26]等通过触角识别的植物挥发物种类均已被鉴定。对昆虫有生物活性的挥发物鉴定后,常结合室内、外行为选择实验测试、验证其引诱或驱避效果,并确定其最佳配比及施用方法,用于害虫引诱剂或驱避剂的筛选与制备,如对茶小绿叶蝉(Empoasca onukii)成虫有显著驱避作用的二甲基二硫醚(DMDS)、1,8-桉叶素和烯丙基甲基硫醚混合,可制备二元驱避剂[27];而壬醛可能成为对多种天牛有效的引诱剂成分[28-29]。
2.2 不同发育状态下昆虫对植物挥发物的识别
昆虫嗅觉识别能力具有一定的可塑性,即依靠不同的生理状态,如日龄、节律等对相同气味作出不同的反应,从而完成特定的行为[30]。Wu等[31]研究发现,小菜蛾(Plutella xylostella)不同日龄成虫对Z11-16:Ac和三种异硫氰酸盐的EAG反应差异显著,而体型大小对其反应无影响,验证了昆虫的生长发育状态影响其对化学信息物质的识别。Pan等[32]鉴定出丙烯酸丁酯、丙酸丁酯和丁酸丁酯是能引起绿盲蝽(Apolygus lucorum)电生理反应的活性挥发物,并发现其寄主植物枣树和葡萄的活性挥发物排放量9月明显高于8月,棉花的活性挥发物排放量在9月有所下降,证明季节动态下,VOCs释放量的变化可调节绿盲蝽对寄主植物的取食,并引导其在不同寄主植物上的产卵。此外,同种昆虫雌雄虫对同一挥发物的反应可能存在差异,Li[33]利用GC-EAD和GC-MS筛选和鉴定出烟草植物挥发物顺-3-己烯-1-醇对马铃薯块茎蛾(Phthorimaeaoperculella)雌雄蛾的定向行为均有显著的引诱作用;草地贪夜蛾(Spodoptera frugiperda)雌蛾对 1 μg/μL 的氧化芳樟醇具有明显的选择偏好性,而雄蛾则显著排斥10 μg/μL的氧化芳樟醇[34]。
2.3 不同环境条件下昆虫对植物挥发物的识别
通常雌虫感受寄主植物挥发物而确定寄主,雄虫则主要通过雌虫释放的性信息素找到配偶,以完成生殖行为,而环境中VOCs的存在可影响其对性信息素的识别。在寄主植物棉花存在的条件下,海灰翅夜蛾(Spodoptera littoralis)雌蛾的求偶起始时间明显提前,求偶持续时间延长,而非寄主植物云杉推迟了雌蛾求偶行为并缩短其持续时间[35]。因此,田间应用中,一些植物挥发物与昆虫性信息素混合可产生更高效的引诱效果[36]。另外,研究表明与郊区相比,一些城市中心的叶甲类害虫危害更为严重,作为嗅觉线索的植物挥发物受大气中臭氧(O3)等空气污染物影响,VOCs的释放量因O3而减少,意味着昆虫可以检测到的嗅觉信息减少,从而改变昆虫的生命活动[37]。例如,O3大大减弱了十字花科植物挥发物对小菜蛾幼虫的吸引力,推测O3迅速降解了VOCs,且降解程度随O3含量增加而增加,证明大气环境条件干扰昆虫确定寄主的位置,在VOCs介导植物与昆虫的相互作用方面具有生态学意义[38]。
3 植食性昆虫触角嗅觉感器及其神经元在识别植物挥发物上的作用
3.1 植食性昆虫触角嗅觉感器及其功能
昆虫触角表面着生多种嗅觉感器,是其识别寄主植物挥发物的重要工具,可利用扫描电镜对其进行鉴定。不同种昆虫嗅觉感器类型及数量均有较大差异,以毛形感器、锥形感器、刺形感器、腔锥形感器、鳞形感器和栓锥形感器等常见[39],不同类型的嗅觉感器可感受不同的化合物成分,如锥形感器可检测到酯类、醇类和醛类气味,毛形感器常识别昆虫性信息素,腔锥形感器可探测酸类等植物挥发物[40]。单感器记录(SSR)表明,脐橙螟(Amyelois transitella)长毛感器对(Z,Z)-11,13-十六碳二烯醛产生反应[41];烟青虫(Helicoverpa assult)位于3种短锥形感器和4种长锥形感器的神经元可对壬醛和庚醛产生响应[42];麦长管蚜(Sitobion avenae)触角中由板状感器、腔锥形感器组成的第六节原生感觉圈对6-甲基-5-庚烯-2-酮能产生较高的反应,且6-甲基-5-庚烯-2-酮在较低剂量(0.01 μg/L)时就能产生明显的刺激波峰[43]。
3.2 植食性昆虫触角嗅觉受体神经元及其功能
昆虫感器感受环境中的VOCs后,将感受到的化学信号转化为电信号,电信号首先经嗅觉受体神经元(olfactory receptor neurons,ORNs)传递到大脑中的初级嗅觉中枢触角叶(antennal lobe,AL),汇聚来的电信号经初步整合加工后投射至高级神经中枢,最终使昆虫产生相应的行为反应[44]。Suk[45]对小菜蛾雌蛾的毛形感器进行了单感器记录,鉴定出几类ORNs,一类特异性识别十字花科寄主挥发物芳樟醇、非寄主挥发物(E)-β-法呢烯等,而其他几类ORNs对非寄主挥发物倍半萜烯、(E)-β-法呢烯、大根香叶烯等反应[46]。ORNs为双极神经元,其细胞体和树突位于触角嗅觉感器内,其主要功能是将触角感器感受到的气味信息传送到触角叶内[47],在高粱金龟子(Pachnoda interrupta)的盘型感器中,大多数ORNs对异戊酸、乙酰乙酯、4-乙基苯酚等化合物响应[48]。
3.3 植食性昆虫嗅觉感觉神经元及其功能
昆虫触角嗅觉感器内含单个或多个嗅觉感觉神经元(olfactory sensory neurons,OSNs),往往具有不同的反应特征,参与昆虫对化学刺激的反应[49]。神经元轴突延伸到AL,AL被分隔成肾小球,在大多数昆虫中,每个肾小球接收来自单一类型OSN的感觉输入,该OSN表达同源的气味受体[50]。如金雀花蛾(Cydia succedana)两种主要的触角OSNs能从其他挥发物中特异识别寄主植物主要挥发物月桂烯和罗勒烯;同时有一类OSNs对很多非寄主植物挥发物有特异性响应,如(E)-β-环状烯、正壬醇、香叶醇等,由此可以加强其对寄主和非寄主的区分[51]。同种雌雄昆虫OSNs类型及数量也存在一定差异,如胡萝卜木虱(Trioza apicalis)[52]、紫花象甲(Sitona disonideus)[53]、Protapion trifolii[54]等特异性识别寄主或非寄主植物挥发物的OSNs类型均被鉴定。马百伟等[55]利用SSR结合神经元示踪技术,记录到棉铃虫(Helicoverpa armigera)雄成虫触角上长毛形感器对性信息素成分Z11-16:Ald的电生理反应显著,并成功染色标记了该感器内两个OSNs,其轴突通过触角神经分别投射AL内的普通神经纤维球和云状体神经纤维球,该技术有助于鉴定昆虫触角OSNs的形态、功能及投射路径,提供新的研究思路。
4 植食性昆虫触角嗅觉相关蛋白在识别植物挥发物上的作用
昆虫对气味分子的识别过程中,外周嗅觉系统对气味分子进行结合和转运,引起电信号的产生、传递与整合,通过轴突传至昆虫中枢神经系统,进而引发昆虫对外界环境的改变做出应答[56-57]。完整的嗅觉识别过程由多种嗅觉相关蛋白的参与及相互协作完成,这些蛋白包括气味结合蛋白(odorant binding proteins,OBPs)、化学感受蛋白(chemosensory proteins,CSPs)、气味受体(odorant receptors,ORs)和感觉神经元膜蛋白(sensory neuron membrane proteins,SNMPs)等[58]。明确这些嗅觉相关蛋白的结构及其功能,将为进一步研究昆虫嗅觉的编码机制奠定基础。
4.1 OBPs结构及其功能
OBPs是一类存在于嗅觉感器淋巴液内的亲水性小分子蛋白(14k~17 kDa),其特征是氨基酸序列中含有6个保守的半胱氨酸位点,它们之间可形成3对二硫键以维持其蛋白结构的稳定性,其与气味分子的结合是昆虫通过嗅觉系统感知外界环境的基础,在昆虫与外界的信息交流中发挥重要作用[59-60]。随着分子生物学技术的发展,很多昆虫OBP基因已被鉴定,且学者们对其功能开展后续研究,如红脊长蝽(Tropidothorax elegans)触角中有14个OBP基因高表达,均属classic OBPs[61];Li[62]鉴定出桃蛀果蛾(Carposina sasakii)12 个CsasOBP基因,氨基酸序列比对证明6个保守半胱氨酸的存在;免疫印迹结果显示,日本松墨天牛(Monochamus alternatus)MaltOBP1在雌雄虫触角中高表达[63]。
一些昆虫OBPs可以特异性识别植物挥发物、昆虫性信息素成分,如Zhang[64]在大肠杆菌中表达了二点委夜蛾(Athetis lepigone)两种AlepGOBP蛋白,研究发现AlepGOBP2与性信息素(Z7-12:Ac、Z9-14:Ac)及玉米植物挥发物[罗勒烯、(E)-β-法尼烯]高度结合;荧光竞争结合分析表明,柑橘木虱(Diaphorina citri)DcitOBP6与β-石竹烯具有更强的结合能力[65];Jing[66]发现松蛀螟(Conogethes pinicolalis)PBP2与E10-16:Ald的结合最强,GOBP1与Z10-16:Ald的结合最强,并揭示了PBP2和GOBP1中与大多数被测挥发物相互作用的潜在活性位点,证明两者可能在松蛀螟识别性信息素和寄主挥发物方面发挥相似作用。关键结合位点的确定可为筛选昆虫引诱剂提供良好的分子靶标,如β-紫罗酮和β-石竹烯与绿盲蝽气味结合蛋白AlucOBP22高度结合,且AlucOBP22的疏水氨基酸残基参与了配体与萜类化合物的特异性结合[67];苹果小吉丁虫(Agrilus mali)AmalOPB8具有较大的配体结合光谱和与植物挥发物结合的灵活性[68]。此外,OBPs还可保护气味分子不被气味降解酶降解;刺激受体后使气味分子失活,避免嗅觉神经元的过度兴奋;清除有毒物质等[69]。最新研究显示,在昆虫口器、足、中肠、腺体等器官中OBPs同样高表达,暗示其可能具有其他不同的生理功能[70],如中华蜜蜂(Apis cerana)足中特异性高表达的AcerOBP15可能参与其采集花蜜时的味觉识别[71]。
4.2 CSPs结构及其功能
与OBPs相比,CSPs是分子量更小,组织分布更广泛的可溶性蛋白,其序列在不同的昆虫中相似性很高,存在明显的高度保守结构域[72],可结合多种化学物质并传递到嗅觉和味觉受体,其识别植物挥发物的嗅觉功能研究也在不断开展,如美洲牧草盲蝽(Lygus lineolaris)2个CSPs(LlinCSP4和LlinCSP8)在触角中特异性表达[73];斜纹夜蛾(Spodoptera litura)中肠中高表达的SlitCSP3、SlitCSP8和SlitCSP11,随着寄主植物种类的改变,表达量也发生变化,表明它们可能具有结合寄主植物挥发物的功能[74]。CSPs可影响植食性昆虫的觅食、产卵等生理活动,如花绒寄甲(Dastarcus helophoroides)幼虫嗅觉基因表达量最高的是DhelCSP3,推测其嗅觉功能在幼虫觅食的过程中发挥关键性作用[75];在柑橘木虱成虫触角中高度表达的DcitCSP1对ρ-伞花烯、E-β-罗勒烯等萜类化合物具有较高的结合力[76];化学感受蛋白的免疫细胞化学定位结果表明,东亚飞蝗(Locusta migratoria)LmigCSP-II在成虫翅的毛形感器中表达,并参与接触性化学感受的过程[77];Li[78]研究发现,烟粉虱(Bemisia tabaci)CSP2可与硝基苯酚(DMNT)等植物挥发物结合,且DMNT可以阻止烟粉虱寄主选择和产卵。
4.3 ORs结构及其功能
昆虫气味受体包括7个跨膜结构域,其N端和C端分别位于细胞质内和细胞质外[79],具有识别气味结合蛋白传递来的气味分子的功能,并介导气味分子与嗅觉感受器内神经元的特异性结合,将化学信号转化为电信号[80-81]。安兴奎[82]、李彬[83]等发现,绿盲蝽气味受体AlucOR91对(-)-4-萜品醇、1-十一醇、十二醇、1-十五烷醇、异樟醇产生响应,AlucOR57对苯甲醛、氧化石竹烯、庚醛等物质有反应,推测两种OR可能与寄主植物定位和选择产卵场所有关;粘虫[84](Mythimna separata)MsepOR13对低至1×10-9mol/L的植物挥发物丁香酚产生反应,因其高度敏感性推测出MsepOR13在其嗅觉系统中可能发挥重要作用;暗黑鳃金龟(Holotrichia parallela)HparOR27已被鉴定对己醛、月桂酸和十四烷三种寄主植物挥发物有反应[85];草地螟(Loxostege sticticalis)LstiOR5在幼虫的触角神经元中高表达,且通过识别多种植物挥发物参与到寄主选择的过程中[86]。
4.4 SNMPs结构及其功能
SNMPs位于嗅觉神经元细胞的树突膜上,代表性的SNMPs有C端和N端两个跨膜结构域[87-88]。一些研究表明,SNMPs不仅是检测昆虫性信息素必不可少的特异性膜蛋白[89],还可能参与识别植物挥发物的过程,如在黑腹果蝇(Drosophila melanogaster)中,SNMP1与识别法尼醇的OR83c相关[90];沙漠蝗(Schistocerca gregaria)的触角上有33个ORs与SgreSNMP1共表达[91];转录组测序分析表明,在棉铃虫幼虫取食拟南芥、棉花、玉米等不同寄主植物后,HarSNMP2和HarSNMP3的表达水平存在显著差异,表明它们可能参与了棉铃虫的取食行为[92];免疫细胞化学定位实验表明,葱蝇(Delia antiqua)的DantSNMP1仅在毛形感器中表达,表明该蛋白可能参与昆虫嗅觉中信息素的接收[93]。研究表明,鳞翅目昆虫中SNMP1普遍在成虫触角上特异性表达,SNMP2在多种组织中广泛表达,而SNMP3在幼虫中肠中特异性表达;RNA干扰和行为分析表明,家蚕(Bombyx mori)SNMP1在信息素检测中起关键作用[94];而OBPs、SNMPs和OR/Orco复合物在气味检测过程中的蛋白-蛋白相互作用及动态特性等仍需进一步的研究和探索[95]。
5 展望
综上所述,研究昆虫的嗅觉系统有助于阐明昆虫个体间的化学通讯机制,了解昆虫对寄主的识别、选择机制,深入理解昆虫的行为,可应用于许多重要农林害虫的防治[96]。许多学者选择多种已知化合物进行昆虫触角电位反应,筛选出该昆虫触角存在应激反应及反应强度较高的化合物,然后进行行为选择实验,验证其对昆虫生物活性的强弱,用于昆虫引诱剂或驱避剂的研发及害虫防治。而从实际角度出发,实验室环境同自然条件间存在较大差异,会影响实验结果在田间的应用效果,因此,实验室研究应追求更加真实地模拟自然环境,以减少由室内、外差异造成的影响[97]。而最新研究提出VOCs可能是干扰而不是引导昆虫觅食等行为[98],因此植食性昆虫与植物挥发物复杂的相互作用关系还需进一步的发现和探索。
植物挥发物的挥发性较强,其样品从收集到测定易受到多种因素影响,进一步准确鉴定出样品中对害虫有引诱或驱避作用的成分、含量、比例较困难。此外,在自然界中,性信息素通常会和植物挥发物同时存在,VOCs可以作为许多昆虫寻找配偶的额外线索,增加性信息素的吸引力或提高昆虫的选择能力,对昆虫求偶行为有一定的促进作用[99]。如水杨酸甲酯或苯甲醛分别与性信息素5:1混用均可以明显提高对禾谷缢管蚜(Rhopalosiphum padi)的引诱能力[100];而水杨酸甲酯能明显降低粉红天社蛾(Thaumetopoea pityocampa)雌蛾性信息素对雄蛾的引诱效果[101]等。综合利用植物挥发物和性信息素有利于昆虫定位寄主植物,刺激雌虫性信息素的释放,使各种昆虫更容易搜寻到交配对象[102]。将VOCs与性信息素混合使用可成为害虫综合防治的新思路,但昆虫性信息素微量分泌,提取比较困难,人工合成成本高,造成田间应用量较低。因此,需要寻找更科学、实用的信息素采集、测定、提取等方法。
近年来,越来越多的昆虫嗅觉蛋白基因被鉴定,昆虫嗅觉识别分子机制研究不断深入,新的研究方法与技术将不断应用到昆虫化学生态的研究中,研制先进、科学、实用的仪器设备会持续受到关注。如Lu[103]设计了一种嗅觉生物传感器,将桔小实蝇(Bactrocera dorsalis)OBPs固定在叉指电极上以检测信息化学物质;在成功分离纯化后,用特殊设计的聚乙二醇(PEG)固定化OBPs,制备了一种性能良好的传感膜,可以灵敏地检测OBPs与昆虫寄主植物释放的不同信息化学物质如乙酸异戊酯、β-紫罗兰酮和苯甲醛之间的相互作用。
猜你喜欢
家教世界·创新阅读(2020年5期)2020-06-22
故事作文·高年级(2019年6期)2019-06-20
小猕猴学习画刊(2019年4期)2019-05-08
扬子江(2019年1期)2019-03-08
小天使·一年级语数英综合(2017年4期)2017-04-18
Coco薇(2016年8期)2016-10-09
作文评点报·作文素材小学版(2016年9期)2016-03-18
红蜻蜓(2015年11期)2016-02-02
知识窗(2014年10期)2014-05-14
