
香苏杂交猪THTPA、TPK1 基因组织表达及其与硫胺素含量的关联性分析
2022-07-13 11:22宋林锦许厚强
中国畜牧杂志 2022年7期
陈 晨,李 永,宋林锦,许厚强*
(1.贵州大学高原山地动物遗传育种与繁殖教育部重点实验室,贵州省动物遗传育种与繁殖重点实验室,贵州贵阳 550025;2.贵州大学动物科学学院,贵州贵阳 550025;3.贵州大学生命科学学院,贵州贵阳 550025)
硫胺素(Thiamine)通过热降解途径分解成含硫化合物以增强肉风味,提升肉类感官评价。硫胺素作为体内的必需物质,不仅可以维持动物机体功能,还可以适当减轻动物应激反应。动物体内不能自主合成硫胺素,在食物摄取不足时,会造成硫胺素缺乏。猪缺乏硫胺素的临床表现为神经症状,最直观的就是会影响猪的食欲,影响增重,严重者还会出现呕吐现象。近年来,国内外对不同物种的、基因进行了大量研究。THTPAse(硫胺素三磷酸酶)是催化硫胺素三磷酸产生硫胺素焦磷酸的重要酶类。而硫胺素焦磷酸作为硫胺素在体内的生物活性物质,其含量多寡直接影响了硫胺素的沉积。属于CYTH 蛋白超家族,CYTH蛋白超家族存在一定的反应条件,需要二价金属阳离子加入才能有活性。Song 等对小鼠的THTPAse 活性进行了探究,分析了THTPAse 的特异性机制与水分子在酶催化中的作用。Rusina 等对哺乳动物线粒体中的THTPAse 活性进行探究,发现这是一种可溶性酶,存在于膜间和基质中。以上结果都说明了THTPAse 以多种形式存在于哺乳动物细胞中,行使不同功能。Piotr等对进行克隆,成功克隆了一个高度特异的序列,其分子量为25 kDa。(硫胺焦磷酸激酶)基因是催化硫胺素生成硫胺素二磷酸和硫胺素二磷酸生成硫胺素三磷酸的关键酶的编码基因。Matthias 等研究发现,基因缺陷会造成硫胺素代谢功能紊乱综合症,引起细胞内硫胺素缺乏,引发相关疾病,在对婴儿缺乏症进行治疗中,口服硫胺素和生物素后,患者临床症状趋于稳定。Sechi 等研究发现,有缺陷病的患者血液与肌肉中的硫胺素焦磷酸的含量会下降,而通过补充适当的硫胺素,可使患者临床症状趋近稳定。王海等对优质肉鸡硫胺素含量以及基因的3' UTR 多态性进行分析,发现突变位点与硫胺素含量差异不显著。王姣等对雏鹅饲喂硫胺素后,发现对基因在肝脏中的表达量有影响。、在硫胺素代谢过程中可能发挥着重要的调控作用,目前对、的研究主要聚焦于基因缺陷、酶活性方面,但香苏杂交猪、表达与硫胺素含量相关性尚不清楚。本实验以香苏杂交猪为研究对象,旨在探究在香苏杂交猪中、相对表达量的变化,并结合硫胺素含量进行相关性分析,探究香苏杂交猪在不同组织中、相对表达量与硫胺素含量的关系,为以香苏杂交猪为基础群体的良种分子选育工作提供参考。
1 材料与方法
1.1 材料
1.1.1 主要试剂 Trizol 试剂(购自上海英骏生物技术有限公司,Gib-co,美国),RevertAid First Strand cDNA Synthesis Kit 逆转录试剂盒(Thermo 公司,货号:K1622),pMD19-T Vector(购自擎科生物,货号:SD4005),硫胺素(维生素B)含量检测试剂盒(购自Solarbio,货号:BC4195),2×Taq PCR StarMix with Loa ding Dye(购自Genstar,货号:A112-05),SYBR Green qPCR Master Mix(购自BIO-RAD,货号:1725271),大肠杆菌()DH5由高原山地动物遗传育种与繁殖教育部重点实验室保存并活化。
1.1.2 样本采集 实验动物采用贵州大学香猪场选育的香苏杂交猪(从江香猪与苏太猪杂交选育,含有75%从江香猪血统的香苏杂交猪)。选择饲养环境相同的180 日龄香苏杂交去势公猪,设计空腹期3 h、12 h、24 h 后进行屠宰(每个处理各3 头)。采取心、肝、脾、肺、肾、大肠(结肠)、小肠(十二指肠)、背最长肌8 个组织,一部分经生理盐水(0.9%氯化钠注射液)、DEPC 水(含0.1% 焦碳酸二乙酯)处理之后,保存于液氮中用于RNA 提取。另一部分经生理盐水处理后,进行硫胺素的测定。
1.2 方法
1.2.1、基因CDS 区片段获取 提取香苏杂交猪各组织总RNA 并逆转录成cDNA。根据NCBI(https://www.ncbi.nlm.nih.gov)中野猪()(XM_001928723)、(XM_013979874.2)的序列,运用Primer Premier 5 软件进行引物设计,通过NCBI 在线预测软件进行引物特异性分析,送至擎科(重庆)生物技术有限公司合成。引物信息见表1。PCR 反应体系为:cDNA 1 μL、正反向引物各1.5 μL(10 μmol/L)、2×Es Taq Master Mix 10 μL,加ddHO补足至20 μL。扩增程序:95℃预变性5 min;95℃ 30 s,54℃/60℃(/)30 s,72℃ 60 s 为一个循环,进行35 个循环;72℃延伸10 min。采用1.5% 的琼脂糖凝胶电泳对PCR 产物进行分离、纯化。

表1 引物序列信息
1.2.2 T 克隆载体的构建 将纯化后的目的片段与pMD19-T 载体进行连接反应,将连接产物转化至大肠杆菌DH5感受态细胞中,划线培养,挑取单个菌落培养,进行菌液PCR 鉴定。选择条带明亮的菌液,送至擎科(重庆)生物技术有限公司进行测序。
1.2.3、基因生物信息学分析 利用生物信息学在线软件ProtParam(http://us.expasy.org/tools/protparam.html/)、SOPMA(http://npsapbil.ibcp.fr/cgi-bin/nps a_automat.pl?page=npsa_sopma.html)和SWISS-MODEL(http://swissmodel.ex-pasy.org/)分别对、基因编码的蛋白质进行理化性质、二级结构和三级结构分析;使用MegAlign 软件进行同源性分析,MEGA 6.0 软件构建遗传进化树。运用DTU 在线软 件(http://www.cbs.dtu.dk/services/)与BUSCA(http://busca.biocomp.unibo.it/)对磷酸化位点、跨膜结构、蛋白信号、亚细胞定位进行分析。通过SMART(http://smart.embl-heidelberg.de/smart/set_mode.cgi?NORMAL=1)进行保守结构功能域分析。
1.2.4、基因表达分析 运用RT-qPCR 法检测香苏杂交猪、基因在不同组织的表达,反应体系(10 μL):cDNA 1 μL、正反向引物各0.5 μL(10 μmol/L)、2×SYBR Green qPCR Master Mix 5 μL,ddHO 补足至10 μL。扩增程序:95℃预变性2 min;95℃ 5 s,58℃/60℃(/)5 s,为一个循环,进行39 个循环,数据采用2值法计算,并用SPSS 18.0 软件进行单因素方差分析。
1.2.5 硫胺素测定 利用硫胺素含量检测试剂盒,测定宰前空腹3、12、24 h 香苏杂交猪8 个组织中的硫胺素含量。采用SPSS 18.0 软件进行数据统计分析。
1.2.6 硫胺素含量与mRNA 相对表达的关系 利用SPSS 18.0 软件对香苏杂交猪8 个组织中和基因相对表达量与硫胺素含量进行相关性分析。
2 结果与分析
2.1、基因CDS 区片段扩增 扩增出、基因CDS 区片段(图1),片段大小均与预期结果相符。

图1 THTPA 与TPK1 基因CDS 区扩增条带琼脂糖凝胶电泳图
2.2 T 克隆载体的构建 将纯化后片段与T 载体连接,转化后进行菌液鉴定(图2),送至擎科(重庆)生物技术有限公司进行测序。
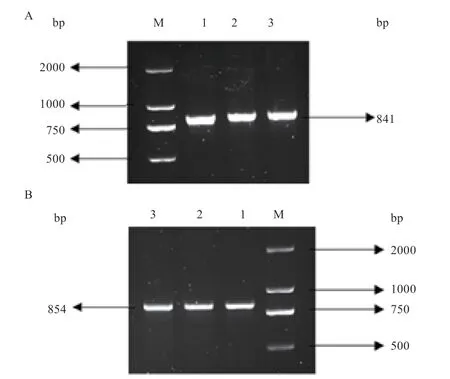
图2 THTPA、TPK1 基因菌液PCR 产物琼脂糖凝胶电泳
2.3、基因生物信息学分析 经过测序发现香苏杂交猪、的CDS 区与NCBI 中的野猪序列(:XM_001928723 和:XM_01397 9874.2)完全一致,不存在突变位点。THTPA、TPK1蛋白质特性如表3 所示。基因编码蛋白不稳定系数为53.19,大于40,属于不稳定蛋白,而基因编码蛋白不稳定系数为34.95,小于40,为稳定蛋白。由亲疏水性可知,两基因所编码蛋白均为亲水蛋白(图3)。THTPA 蛋白主要由3 种结构组成,其中40.87% 为-螺旋,延伸链区域占13.91%,无规则卷曲占45.22%。TPK1 主要由3 种结构组成,其中37.97% 为-螺旋,延伸链区域占15.61%,无规则卷曲占46.41%。蛋白质三级结构预测结果表明,香苏杂交猪THTPA 和TPK1 蛋白的三级结构与二级结构相符(图4),主要由无规则卷曲和-螺旋构成。THTPA有60.9% 定位于细胞质,位于细胞核、线粒体、分泌囊泡、细胞骨架的比例分别为17.4%、8.7%、8.7% 和4.3%。TPK1 主要定位于细胞质,占65.2%,线粒体和细胞核占8.7%,液泡、细胞骨架、分泌囊泡、高尔基体均占4.3%。进行相关性分析发现THTPA 与TPK1 存在一定程度上的相互关系(图3C)。遗传进化树分析可知(图5),香苏杂交猪、基因和野猪完全同源,在哺乳动物中香苏杂交猪、基因遗传距离都较斑马鱼近;香苏杂交猪基因与鸡的遗传距离最远,基因与狗的遗传距离最远,符合物种遗传进化论。采用MegAlign 对13 种物种的和基因核苷酸序列进行同源性分析,结果(图6A)所示,香苏杂交猪基因核苷酸同源性与野猪一致,同源性为100%,与斑马鱼相似性最低,同源性为42.2%。除野猪外基因与水牛的相似性最高(87%)。香苏杂交猪基因核苷酸同源性(图6B)与野猪一致(100%),其次是山羊(90.4%),与斑马鱼相似性最低(57.6%)。基因在所对比的哺乳动物中,同源性都高于80%,表明该基因在哺乳动物中可能会有相似的功能。
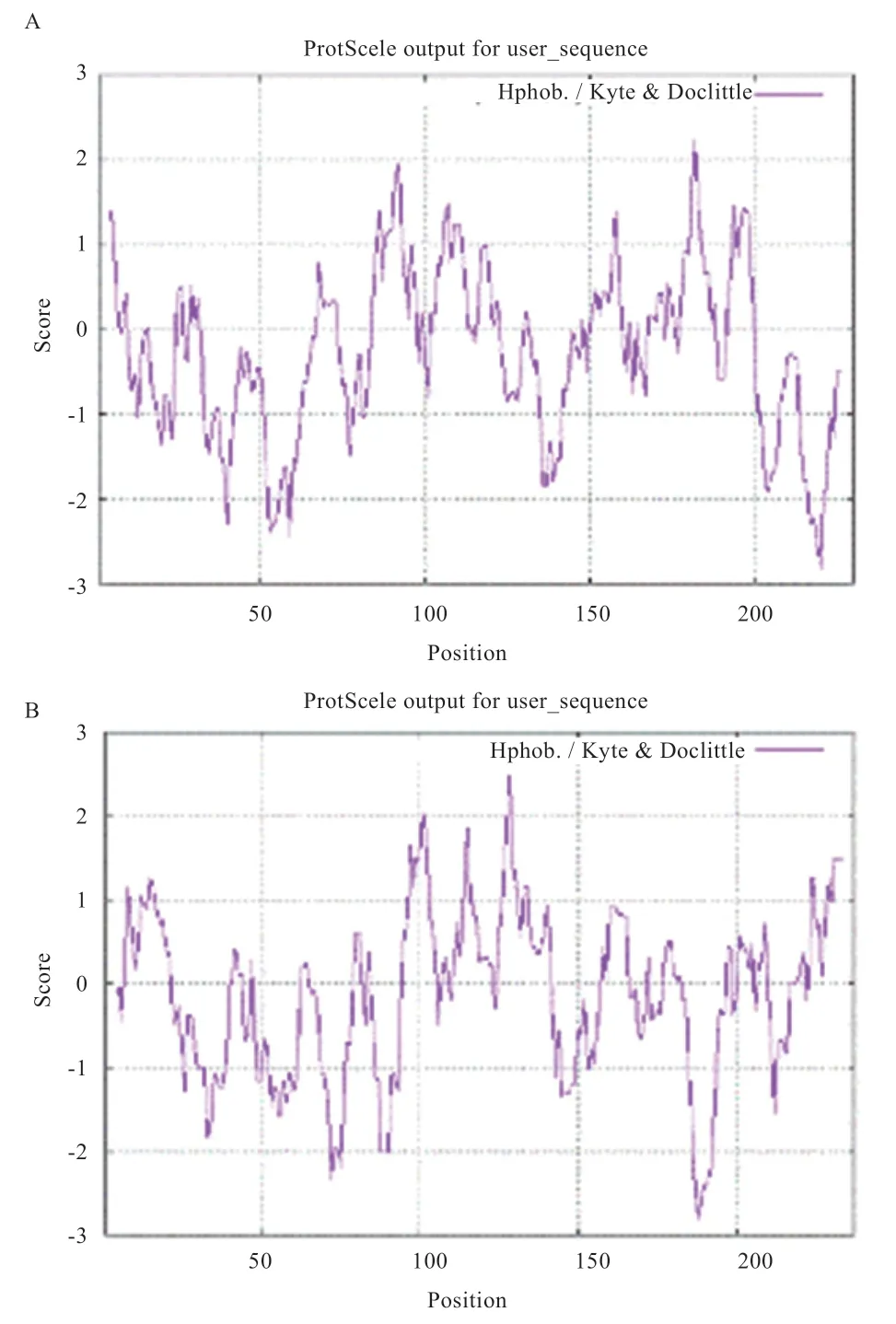
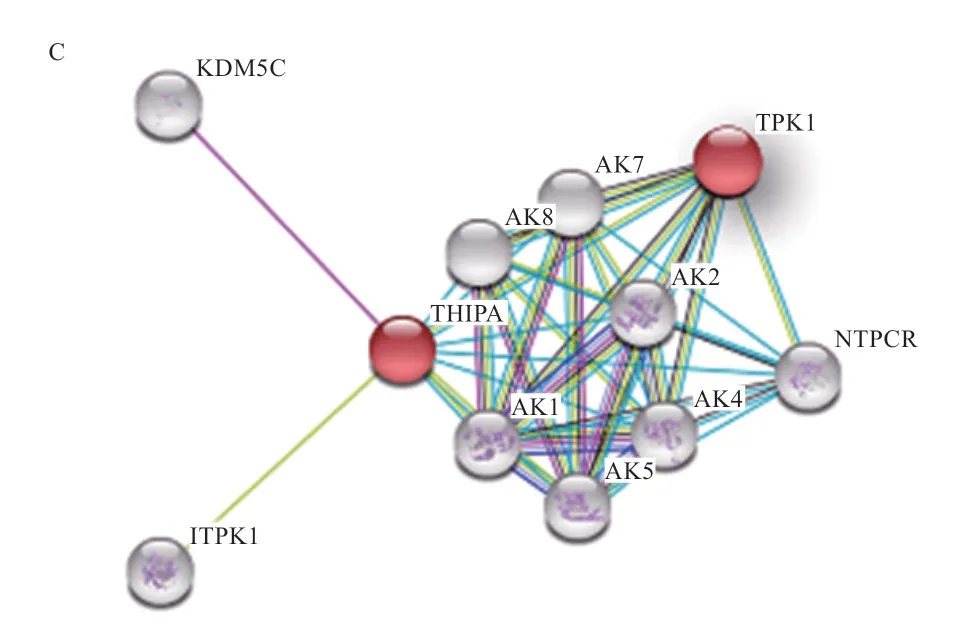
图3 THTPA 与TPK1 亲疏水性及相互关系

图4 THTPA、TPK1 二级、三级蛋白结构分析图
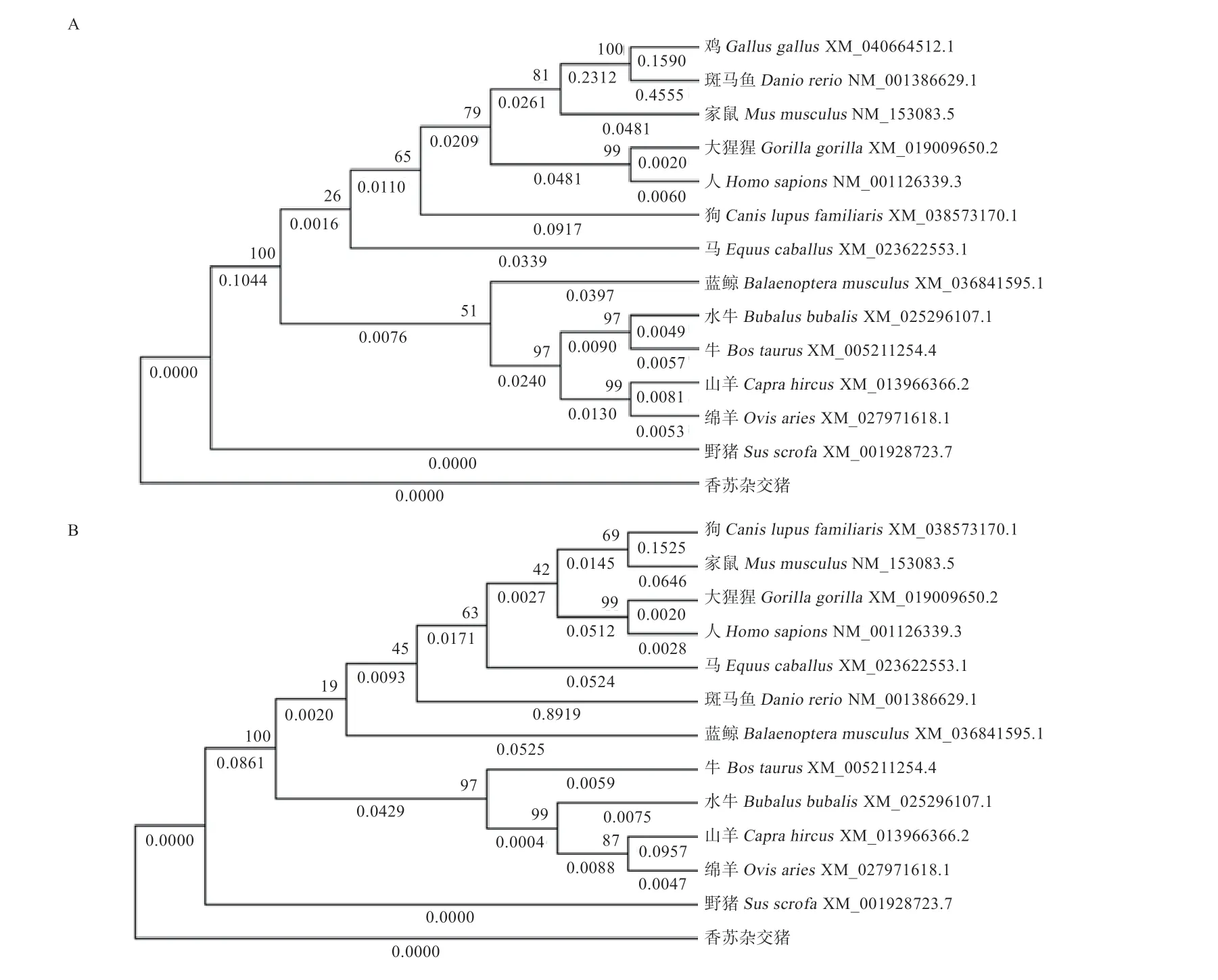
图5 香苏杂交猪THTPA、TPK1 基因遗传进化分析
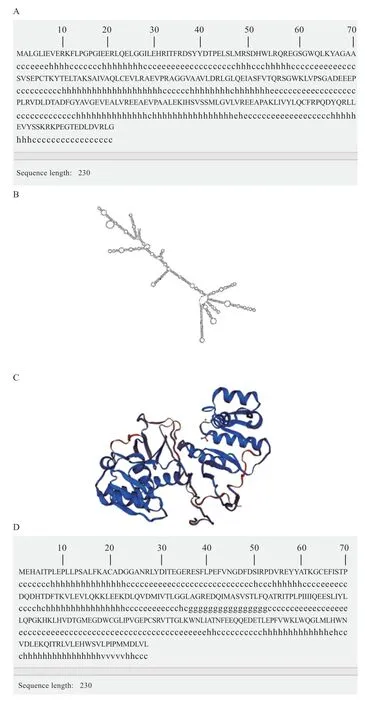

图6 不同物种THTPA 基因(A)和TPK1 基因(B)核苷酸序列同源性及其遗传距离分析结果
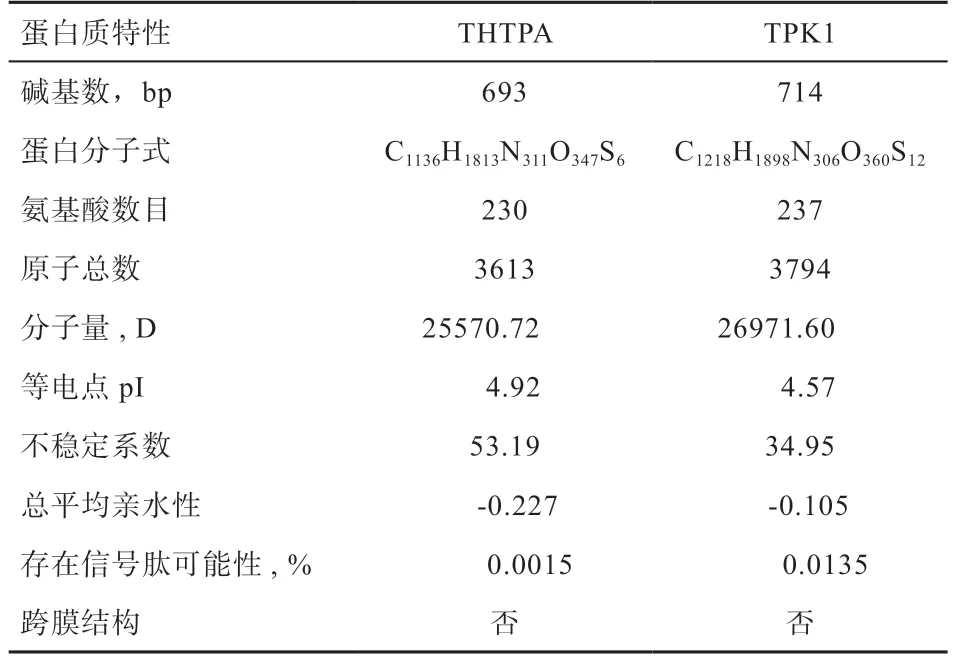
表3 THTPA、TPK1 蛋白基本性质
2.4、基因在香苏猪不同组织中的表达分析 用不同组织不同空腹处理时间cDNA 为模板进行荧光定量PCR 检测,以探究、基因在香苏杂交猪不同屠宰空腹时间各组织中的表达规律。结果(图7)表明,、在香苏杂交猪的各个组织中均有表达,且不同空腹屠宰处理下,、基因表达量存在显著差异。其中在对基因相对表达量检测时,处理3 h 时肝脏中的相对表达量最高,12、24 h 为脾脏中的相对表达量最高。基因在处理3、12 h 时肝脏中的相对表达量最高,24 h 时大肠(结肠)表达量最高。
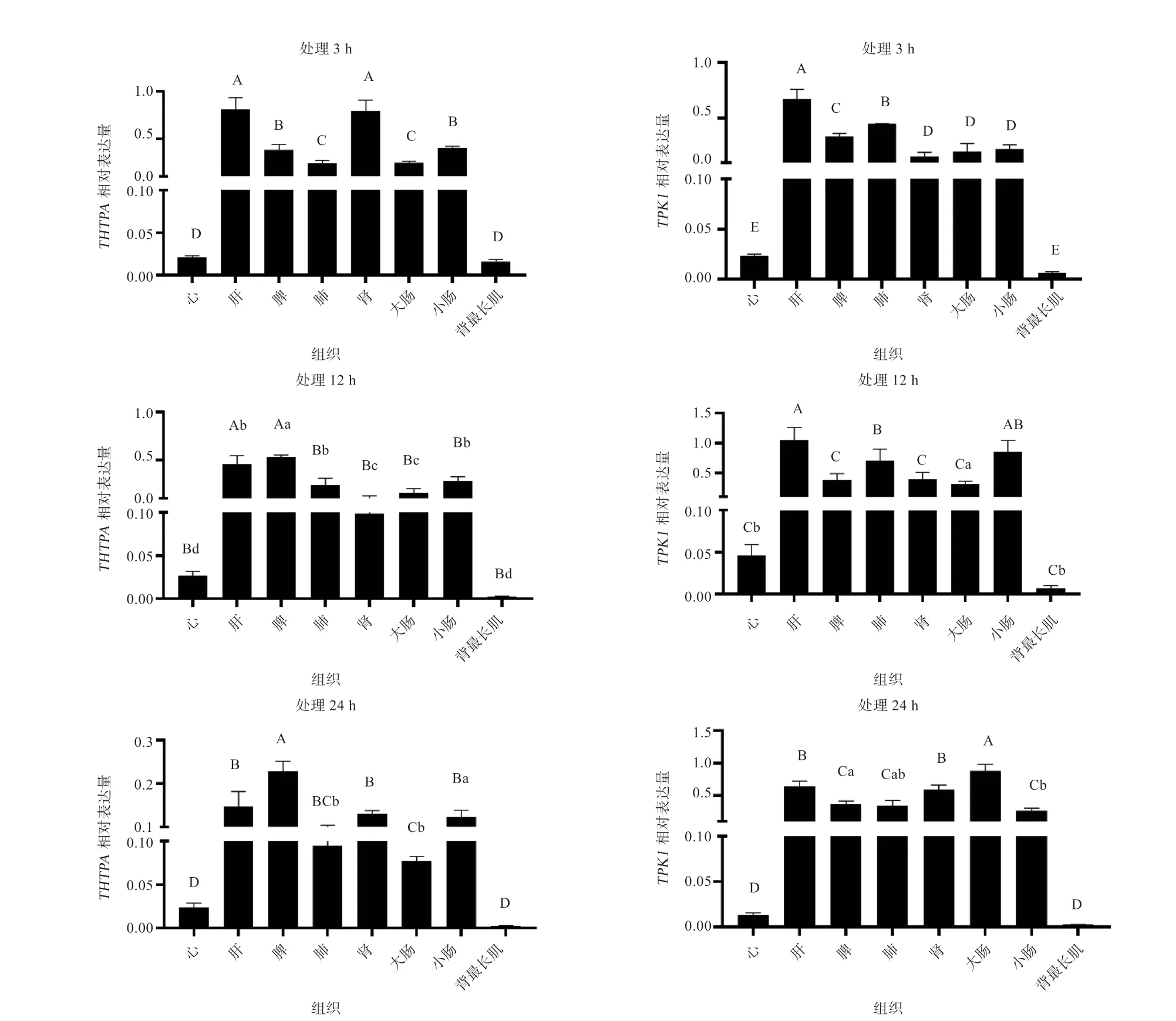
图7 THTPA、TPK1 基因不同处理时间不同组织相对表达量
2.5 硫胺素测定 测定香苏杂交猪8 个组织不同处理时间硫胺素含量(表4)。发现背最长肌中各个阶段硫胺素含量无差异。在处理3 h 时小肠、肾脏中硫胺素含量均上升,此时硫胺素含量最高为肝脏(20.937±2.850 mg/g),其次为小肠(17.514±1.717 mg/g)。在处理12 h 时,肝脏中硫胺素含量依旧最高(21.314±0.342 mg/g),且比处理3 h 有所上升。3 h 时,小肠中硫胺素含量急剧下降(6.902±0.384 mg/g),脾脏中硫胺素含量上升(19.881±0.334mg/g),在24 h 处理下,除肝脏、心脏、背最长肌外各个组织硫胺素含量均有下降。
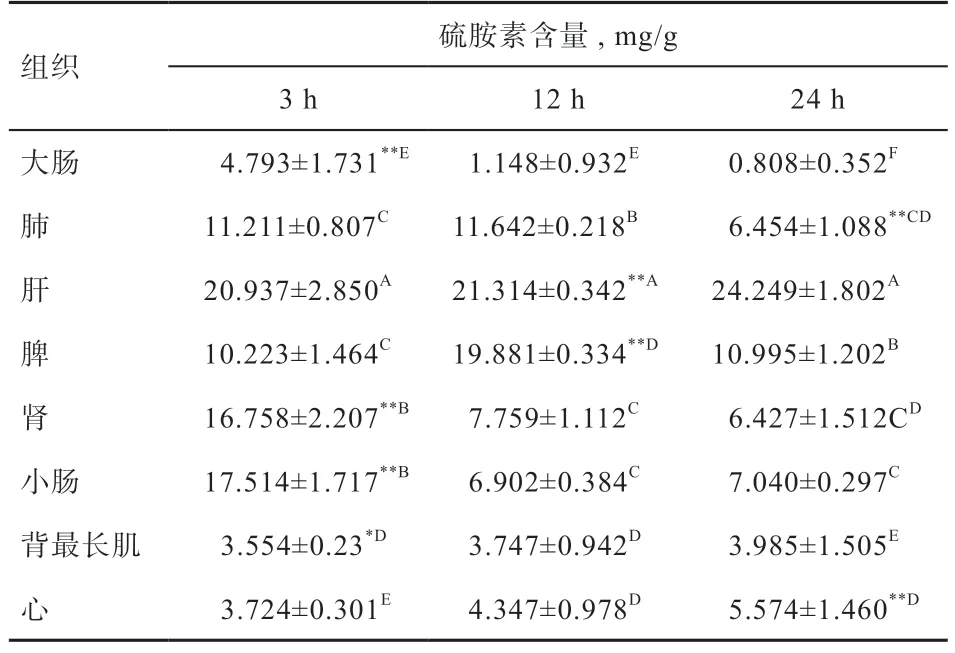
表4 不同组织与不同处理中硫胺素含量
2.6 硫胺素含量与mRNA 相对表达量的关系 对香苏杂交猪、基因在不同阶段各组织中的表达规律进行分析(图8)。基因在脾脏、心脏和背最长肌无明显变化趋势,在肺脏、小肠(十二指肠)、肝脏中先上升后下降。肾脏、大肠(结肠)相对表达量趋势随着空腹时间增加而上升。基因不同组织中的相对表达量在24 h 时都趋于稳定。背最长肌与心脏呈现直线趋势,并无明显变化。其余组织随着时间增加,表达量下降,最后均趋于稳定。背最长肌中各个阶段硫胺素含量无差异。在处理3 h 时小肠、肾脏中硫胺素含量均上升,在处理12 h 时,肝脏中硫胺素含量最高,比处理3 h 有所上升。小肠(十二指肠)中硫胺素含量急剧下降。脾脏中硫胺素含量上升,在24 h 处理下,除肝脏、心脏、背最长肌外各个组织硫胺素含量均有下降,通过SPSS.18.0 软件对心、肝、脾、肺、肾、大肠(结肠)、小肠(十二指肠)、背最长肌的、基因相对表达量和硫胺素含量进行相关性分析(表5)。、基因相对表达量极显著相关(0.483,<0.01),基因相对表达量与硫胺素含量极显著相关(0.657,<0.01),基因相对表达量与硫胺素含量极显著相关(0.378,<0.01)。

表5 THTPA、TPK1 基因相对表达量和硫胺素含量相关性分析

图8 香苏杂交猪THTPA、TPK1 基因与硫胺素在不同空腹阶段各组织的变化趋势
3 讨论
研究表明,硫胺素作为外来风味前体物质,可以降低美拉德反应的褐变作用,提高肉类感官评价。基因在各个组织均有表达,但是酶活性却极低,说明这种酶类除了可以分解硫胺素以外,还存在其他功能。Bernard 等对基因在啮齿动物中的表达量进行探究后发现在睾丸中基因相对表达最高,肝中的酶活性最高,这说明基因相对表达量与THTPAse 酶活性关联性不显著。本实验使用qRTPCR,检测了香苏杂交猪心、肝、脾、肺、肾、大肠(结肠)、小肠(十二指肠)、背最长肌8 个组织中、基因相对表达量。24 h 空腹处理时,基因各组织相对表达量排序为:脾>肝>肾>小肠(十二指肠)>肺>大肠(结肠)>心>背最长肌。生物信息学分析发现基因编码蛋白分子质量为25 kDa,与人和小鼠的分子量25 kDa 相同,与牛24 kDa 存在一定的差异。通过亲缘性对比,与牛的亲缘性最高(86.7%)。Marjorie 等发现THTPAse 在人类中所占的比例高于啮齿类动物,该酶的表达和活性与细胞分化程度有关,即与物种的进化分化相关。有60.9% 定位于细胞质。亚细胞定位最大可能于细胞质,占65.2%。Czerniecki 等对啮齿动物的进行定位发现其存在于神经细胞胞质内。Rusina 等对牛THTPAse 活性进行探究,也发现酶类存在于细胞质中。这与香苏杂交猪的亚细胞定位预测的结果一致,说明可能主要定位于细胞质中且在不同物种中可能都存在于细胞质中。对背最长肌与肝脏中硫胺素含量与基因相对表达量进行分析,肝脏中的表达量与硫胺素含量最高。
基因各组织相对表达量排序为:大肠(结肠)>肝>肾>脾>肺>小肠(十二指肠)>心>背最长肌,硫胺素的含量与基因相对表达量呈现正相关。Anna 等研究发现,基因缺陷病通过补充适当的硫胺素,可使患者临床症状趋近稳定。王娇等对雏鹅饲喂硫胺素后,发现对基因在肝脏中的表达量有影响,与本实验结果一致,说明体内硫胺素变化会影响基因表达量改变,硫胺素与基因在肝脏中的表达有一定影响。对不同空腹处理的香苏杂交猪的肝脏和背最长肌进行硫胺素的测定,结果显示香苏杂交猪背最长肌硫胺素的含量为3.536±0.253 mg/g。李翔等测定不同品种选育猪背最长肌硫胺素的含量,CH 猪硫胺素含量为3.989 mg/g,S05 猪肉中硫胺素含量为2.737 mg/g,不同猪种中背最长肌中硫胺素含量存在差异,这可能是地方猪种的特性,其机理还有待探究。不同组织中的硫胺素含量不同,且随着空腹时间的增加在大肠(结肠)、小肠(十二指肠)、肺、肾这些组织中硫胺素含量下降,在肝、心这2 个组织中硫胺素含量上升,这是由于不同组织的代谢能力差异而普遍存在。在背最长肌中硫胺素含量与基因相对表达量稳定,这就说明,基因在背最长肌中的功能并不主要为硫胺素代谢。
本研究利用qRT-PCR 检测、基因在香苏杂交猪不同空腹时间屠宰后的表达规律,结果发现,、基因在香苏杂交猪不同阶段的心脏、肝脏、脾脏、肺脏、大肠(结肠)、小肠(十二指肠)、背最长肌中均不同程度表达,其中,内脏组织的相对表达量均高于肌肉组织,说明、基因属于广谱表达基因,且在肌肉组织中表达暗示着在香苏杂交猪的、基因可能存在硫胺素代谢以外的功能活性。、基因相对表达量受屠宰空腹时间影响,也就是受到硫胺素吸收代谢的影响,且存在一定的显著性差异。基因在脾脏、心脏和背最长肌中表达量在不同空腹屠宰处理时间无明显变化趋势;在肺脏、小肠(十二指肠)、肝脏中趋势为“A”型模式,先上升后下降;肾脏、大肠(结肠)相对表达量趋势随着空腹时间增加而上升。基因在不同组织中的表达趋势,除肺脏以外,表达量随着空腹时间增加均下降。这说明、基因表达量与硫胺素吸收代谢有一定关系。此外,、基因在香苏杂交猪肝脏中的相对表达量较高,暗示、基因有可能参与了硫胺素代谢等调控功能。在背最长肌中硫胺素含量与、基因表达量都趋近于稳定,这就说明即使不参与硫胺素代谢,可能也参与了其余的生物学功能。
4 结论
本研究成功克隆出香苏杂交猪、基因,并进行生物信息学、组织表达特征分析,、基因在各个组织中都有表达,且在不同处理条件下,肝脏都有较高的表达,在背最长肌中表达量最低。、基因相对表达量相关性极显著,基因相对表达量与硫胺素含量极显著相关,基因相对表达量与硫胺素含量极显著相关。本研究结果可为硫胺素沉积研究提供分子方面的理论依据,为后期硫胺素胁迫研究提供理论支撑。
猜你喜欢
农家参谋(2021年2期)2021-04-23
新传奇(2021年3期)2021-02-26
华声(2020年9期)2020-10-20
福建基础教育研究(2019年2期)2019-09-10
福建基础教育研究(2019年2期)2019-05-28
故事会(2019年10期)2019-05-27
食品与健康(2018年3期)2018-03-29
家庭医药(2017年9期)2017-09-14
小学生导刊(低年级)(2016年8期)2016-09-24
饮食科学(2016年3期)2016-07-04
