
庭疾灶螽中肠及马氏管结构
2022-07-12 02:43张力丹龚旭平彭薪霏石福明常岩林
昆虫学报 2022年6期
张力丹, 龚旭平, 彭薪霏, 石福明, 常岩林
(河北大学生命科学学院, 生命科学与绿色发展研究院, 河北省动物系统学与应用重点实验室, 河北保定 071002)
消化道是消化系统的重要组成部分,其主要功能是提高食物消化与吸收的效率,保护生物体免受有害化学物质、微生物和食物机械损伤的伤害,昆虫消化道的结构和功能反映出对食物的高度适应性(Klowden, 2013; Holtofetal., 2019)。昆虫中肠是消化道结构和功能的中心,中肠前端的胃盲囊是长期进化的结果。中肠上皮行使小分子的有机营养物质、离子和水的运输,是内外环境的分界(Lietal., 2018; Cacciaetal., 2019)。目前,国内外对蝗虫消化系统的研究报道较多,包括消化道的基本结构与中肠细胞吸收、代谢的功能等(Chauvin, 1938; Droste and Zebe, 1974; Maranaetal., 1997; 甘雅玲和郭中伟, 2002; 张小民等, 2006)。对蟋蟀科消化系统的形态、中肠细胞的结构及分泌机制等方面也有研究结果,如蟋蟀科的消化道中肠具有两个胃盲囊,中肠主要由柱状细胞和再生细胞组成,柱状细胞中同时存在局部分泌和顶浆分泌两种分泌机制;中肠细胞的超微结构表明它们参与液体的分泌(从细胞到内腔)和吸收(从内腔到细胞)等(Woodring and Lorenz, 2007; Biagioetal., 2009; Çakici and Ergen, 2012)。此外,Li等(2018)对螽斯科优雅蝈螽Gampsocleisgratiosa的中肠结构、中肠细胞类型及分泌途径等也有报道。但是,关于驼螽科消化系统的研究,却罕见报道。
当食物分解,营养被吸收后,剩余的残留物与代谢产物必须排出体外或隔离,以防止影响其他新陈代谢途径。昆虫的排泄系统主要是马氏管,它的作用是调节渗透压、清除新陈代谢最后产物,并将有毒化合物储存隔离或转移到后肠,最后经肛门排出体外(Klowden, 2013; Amutkan-Mutlu and Suludere, 2020)。 Polat等(2017)发现蟋蟀科的Grylluscampestris马氏管的管腔侧有紧密排列的微绒毛结构,但微绒毛中没有线粒体;微绒毛端部膨大呈圆球状,此结构包含的囊泡被证实是顶浆分泌物;Amutkan-Mutlu等(2021)利用光学显微镜、扫描电子显微镜和透射电子显微镜观察了螽斯科的Leptophyesalbovittata马氏管上皮细胞的结构,依据细胞质电子密度的差异认为马氏管细胞中有2种细胞类型,此外发现马氏管细胞中的球晶富含碳、磷和锰化学成分。
庭疾灶螽Tachycinesasynamorus隶属于直翅目(Orthoptera)驼螽科(Rhaphidophoridae),体型中等偏小,分布广泛。驼螽科昆虫完全无翅,体侧扁,体背隆突;头顶具纵沟;足较细长,缺跗垫,前足基节背面通常具小刺,前足胫节无听器(Adelung, 1902);通常群居生活,栖息于石缝、枯枝落叶层、洞穴等较阴暗潮湿的环境,是直翅目仅有的腐生性类群。Novak和Kustor(1983)发现驼螽科的Troglophiluscavicola和Troglophilusneglectus的生活史为2年,并以成虫越冬;Ander(1939)对螽亚目进行系统发育分析认为驼螽科是最原始的螽斯类昆虫。本研究以庭疾灶螽为材料,应用光学显微技术、激光共聚焦显微技术和透射电镜技术对其消化道的形态结构、中肠和马氏管的组织结构进行研究,旨在为驼螽科物种的内部结构研究提供基础资料,探索消化系统和排泄系统在结构上与食性的适应关系。
1 材料与方法
1.1 试虫来源
庭疾灶螽成虫,于2020年9月及2021年8月采集于河北省保定市莲池区河北大学校园。
1.2 解剖学观察
选取雌、雄成虫各5头,在LEICA M250A体视镜下,用眼科剪沿虫体侧膜剪开后去掉体壁,去除气管、脂肪组织后,露出完整的消化道。将解剖分离的消化道投放到盛有PBS缓冲液的干净培养皿中进行拍照,所用单反相机型号是Canon E0S 7D。
1.3 显微结构观察
1.3.1苏木精-伊红(hematoxylin-eosin, H.E.)染色:将中肠及马氏管置于Bouin氏固定液中固定,经梯度酒精脱水,二甲苯透明,石蜡渗透包埋后,石蜡切片机切片。室温下进行常规的苏木精-伊红染色,Olympus BX51光学显微镜观察拍照。
1.3.2高碘酸-Schiff试剂(periodic acid Schiff, PAS)染色:固定、脱水等制作石蜡切片步骤同1.3.1节。PAS染色时先经过二甲苯脱蜡,之后经过梯度酒精直至到蒸馏水,之后在4%的高碘酸水溶液中处理5 min, PAS中染色15 min。再经过3次亚硫酸水溶液处理,苏木精复染细胞核,后续过程经梯度酒精脱水和二甲苯透明后,中性树胶封片,Olympus BX51光学显微镜观察拍照。
1.3.3微丝、微管荧光染色:选取消化道的中肠部分置于4%多聚甲醛溶液中,4℃下固定24 h。然后在0.5 mol/L蔗糖中脱水,-20℃下用O.C.T.包埋。用Leica CM1860冷冻切片机将包埋块切成5 μm厚的冰冻切片,并粘附于载玻片上。
将10余根马氏管置于滴有4%多聚甲醛溶液的多聚赖氨酸载玻片上,固定10 min。然后用蒸馏水冲洗干净,用吸水纸吸去多余水分后晾干。
在室温下,用1% Triton X-100将中肠切片及马氏管装片处理30 min。然后用5% BSA封闭处理1 h;接着用异硫氰酸标记的抗α-微管蛋白鼠单克隆抗体(anti-α-tubulin-FITC, mouse monoclonal, Sigma-Aldrich)1∶100稀释,室温下孵育1 h,PBS漂洗3次。再将组织用鬼笔环肽-罗丹明(phalloidin-tetramethylrhodamine B isothiocyanate, Sigma-Aldrich)染色45 min,PBS漂洗。接着,DAPI染色15 min,PBS漂洗。之后在FV3000激光共聚焦显微镜下观察,拍照。
1.4 超微结构观察
将切割成约1 mm3的新鲜中肠及马氏管置于2.5%戊二醛溶液中前固定。后转至四氧化锇溶液中进行后固定。梯度酒精脱水后,分别在丙酮、丙酮/环氧树脂Epon812(1∶1, v/v)和纯环氧树脂Epon812中渗透。最后于纯Epon812中进行室温下聚合12 h,60℃聚合48 h。用Reichert-Jung超薄切片机切成厚度70 nm的超薄切片,用柠檬酸铅和醋酸铀双染后,在日立HT7800透射电子显微镜下观察(加速电压80 kV)并拍照。
2 结果
2.1 消化道形态
庭疾灶螽的消化道由前肠、中肠和后肠3部分构成。前肠包括咽、食道、嗉囊和前胃;中肠包括前端的胃盲囊和中肠;后肠包括回肠、结肠、直肠和肛门(图1: C)。咽与食道较短,前端囊状的为接纳和临时贮存食物的嗉囊,嗉囊是消化道中除中肠以外最长的部位,整体呈不规则形,前端细,后端则明显膨胀,背面呈圆弧形,腹面弧度较小(图1: C)。嗉囊后端缩细与前胃相连。在组织学结构上,前胃是消化道最为骨化的部分(图2: A-B)。从胃盲囊延伸到马氏管着生处的管道为中肠。细长的马氏管在中、后肠交界处着生。马氏管着生处之后的肠道为后肠,后肠的前部分粗细均匀,属于回肠;中部细而长,属于结肠;后端有所膨大,为直肠,直肠通向肛门。

图1 庭疾灶螽生态照片及雄性消化道解剖结构Fig. 1 Ecological image and anatomy of male digestive tract of Tachycines asynamorusA: 雌成虫Female adult; B: 雄成虫Male adult; C: 雄成虫消化道解剖结构Anatomical structure of digestive tract of male adult. Co: 结肠Colon; Cr: 嗉囊Crop; Gc: 胃盲囊Gastric caeca; Il: 回肠Ileum; Mt: 马氏管Malpighian tubules; Pv: 前胃Proventriculus; Re: 直肠Rectum.
2.2 胃盲囊显微结构
胃盲囊是中肠前部3个向前延伸的球状结构(图2: A)。3个胃盲囊合抱前胃,是中肠的起始部位(图2: A-B)。胃盲囊内无几丁质内膜。胃盲囊上皮细胞排列有一定规律性,可见2类上皮细胞交替排列。一类为靠近基部的再生细胞,另一类是端部覆盖有微绒毛的柱状上皮细胞。上皮细胞外侧依次为环肌和纵肌(图2: C)。胃盲囊上皮细胞基底膜处PAS染色为红色(图2: D),推测基底膜具有复合糖成分。

图2 庭疾灶螽胃盲囊Fig. 2 Gastric caeca of Tachycines asynamorusA: 胃盲囊解剖结构Dissected gastric caeca; B: 胃盲囊整体横切石蜡切片H.E.染色 Paraffin transection of gastric caeca stained with hematoxylin-eosin; C: 胃盲囊部分横切石蜡切片H.E.染色Part of gastric caeca transection stained with hematoxylin-eosin; D: 胃盲囊部分横切石蜡切片PAS-苏木精染色Part of gastric caeca transection stained with PAS-hematoxylin. Bl: 基膜Basal lamina; Cc: 柱状细胞Columnar cell; Cm: 环肌Circular muscle; Gc: 胃盲囊Gastric caeca; Lm: 纵肌Longitudinal muscle; Mv: 微绒毛Microvilli; Pv: 前胃Proventriculus; Rc: 再生细胞Regenerative cell.
2.3 中肠显微结构
中肠横切面近环形(图3: A-B)。中肠壁结构从外到内可以分为5层:浆膜、环肌、纵肌、基底膜、上皮层。中肠上皮细胞排列整齐,由柱状上皮细胞、再生细胞和内分泌细胞组成,其中柱状上皮细胞与内分泌细胞由再生细胞分化而来。柱状上皮细胞形态大小均一,紧密相贴,在端部特化成整齐排布的微绒毛,被鬼笔环肽-罗丹明染成红色;核大呈圆形,被DAPI染成蓝色(图3: B)。
再生细胞位于上皮层基部,聚集形成龛(niche)。再生细胞体积较柱状细胞小,细胞核相对大,包括干细胞和分化中的细胞。有的再生细胞龛中可以观察到正在进行有丝分裂的干细胞(图3: C-D)。内分泌细胞分布于柱状上皮细胞中,数量较少。光学显微镜下,内分泌细胞不易与柱状上皮细胞区分。
中肠肌肉的层次排列为纵肌在内,环肌在外。在中肠多见顶浆分泌,顶浆分泌时,细胞膜顶端多处发生破裂,将细胞质、细胞器及其分泌物一起排入中肠腔中(图3: C)。此外,在整个中肠腔内观察到围食膜包裹食物团,并且在围食膜和上皮之间有间隙(图3: A)。中肠基底膜较厚,经过PAS染成红色(图3: D)。
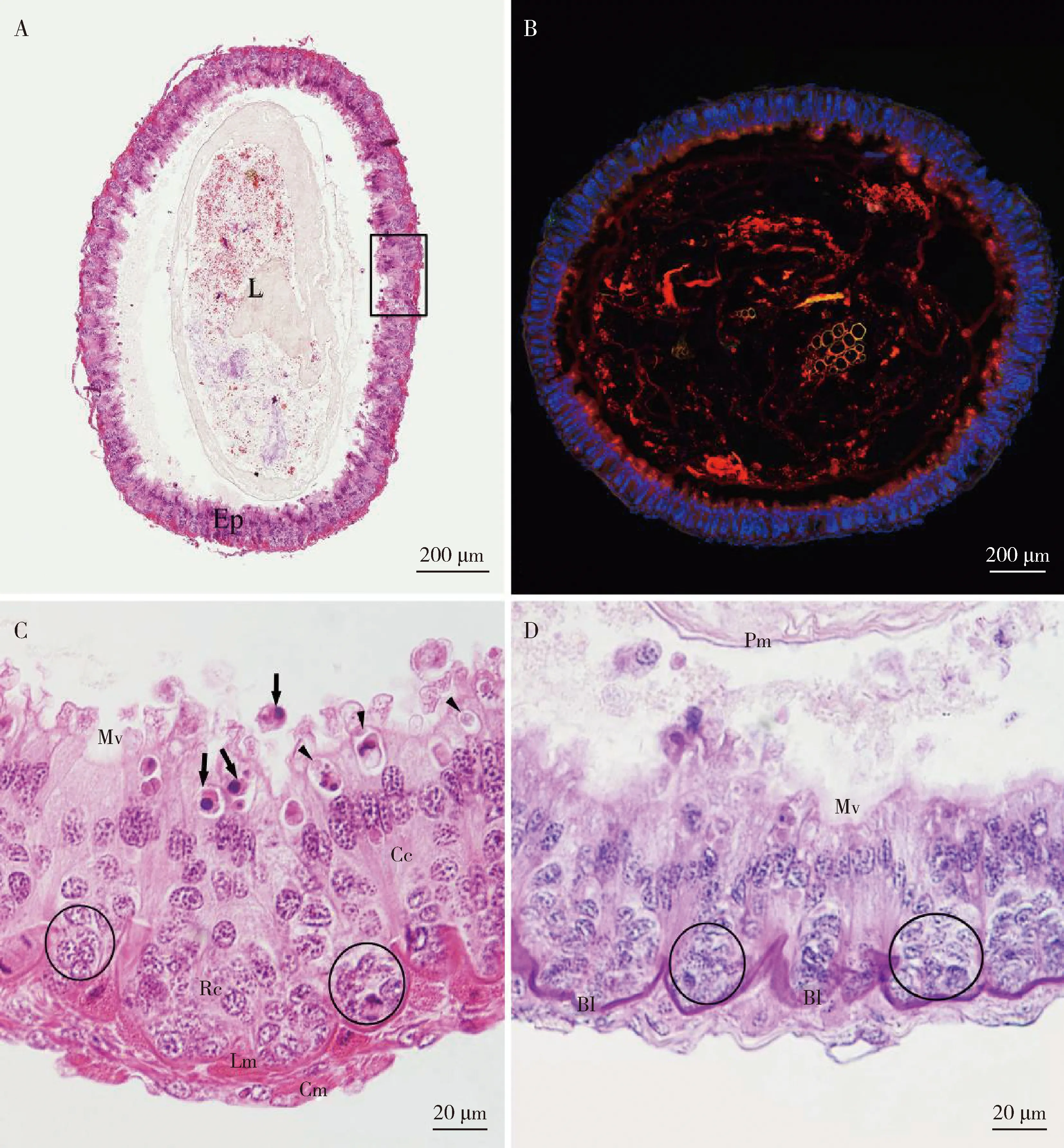
图3 庭疾灶螽中肠Fig. 3 Midgut of Tachycines asynamorusA: 中肠整体横切石蜡切片H.E.染色,方框内局部结构为图C所示Paraffin transection of midgut stained with hematoxylin-eosin and the local structure in line frame shown in Fig. C; B: 中肠冰冻切片免疫荧光染色Frozen section of midgut stained with immunofluorescence; C: 中肠横切石蜡切片H.E.染色,圆圈内为龛, 箭头示球状分泌物质和三角形示线团状分泌物质Paraffin transection of midgut stained with hematoxylin-eosin, inside the circle being the niche, the arrows indicating the secretory substances in spherical particle and triangles indicating the secretory substance like linear clump; D: 中肠石蜡切片PAS染色显示基膜呈红色,中肠腔内有明显围食膜Paraffin section of midgut stained with PAS-hematoxylin showing the basal lamina stained in red and a distinct peritrophic matrix in the lumen. Bl: 基膜Basal lamina; Cc: 柱状细胞Columnar cell; Cm: 环肌Circular muscle; Ep: 上皮Epithelium; L: 内腔Lumen; Lm: 纵肌Longitudinal muscle; Mv: 微绒毛Microvilli; Pm: 围食膜Peritrophic matrix; Rc: 再生细胞Regenerative cell.
2.4 中肠上皮细胞超微结构
2.4.1柱状上皮细胞:细胞端部伸到肠腔,细胞的游离面较宽,基部较细(图4: A)。细胞中含有发达的粗面内质网、大量线粒体和电子密度不同、大小不等的分泌泡分布在整个细胞质内(图4: A)。核周区和端部的细胞器类型一致。在细胞端部区域,细胞紧密地贴在一起,较长的间壁连接呈直线型,维持着上皮细胞间的联系(图4: B)。细胞核呈卵圆形,位于细胞的基部区域,包含明显的核仁和随机分布的不规则团块状的异染色质。柱状上皮细胞顶膜特化形成微绒毛来扩大吸收的表面积,在电子显微镜下可以看到微绒毛中轴含有纵行微丝(图4: A-B)。
电子显微镜下可以观察到柱状细胞在靠近微绒毛的细胞质中,有两类被细胞质包裹的分泌物:一种是由电子密度低的物质包被的线团状结构,另一种是电子密度很高的球状结构(图4: C)。柱状上皮细胞微绒毛可以被鬼笔环肽-罗丹明染成红色,同时线团状结构被鬼笔环肽-罗丹明染成鲜亮的红色(图4: G),但线团状结构被异硫氰酸标记的抗α-微管蛋白抗体染的绿色不明显(图4: F)。结合电镜超微结构,推测柱状上皮细胞微绒毛的主要成分是微丝,柱状上皮细胞分泌物的线团状结构主要由大量微丝包裹而成,最后通过顶浆分泌运输到胞外(图4: C-D)。此外,一些电子密度程度不同的分泌物囊泡也通过此类方式移出胞外(图4: E)。

图4 庭疾灶螽中肠的柱状上皮细胞Fig. 4 Columnar cells of Tachycines asynamorusA: 中肠柱状上皮细胞Columnar cells of the midgut; B: 细胞微绒毛端部形成囊泡释放入管腔中,小图示微绒毛横切Apical part of microvilli of columnar cell generates small vesicles and releases into the central lumen, and the inset shows the detail of microvilli transection; C: 细胞近端部分布2种分泌物质,一种呈线团状,一种呈球状颗粒, 小图示包被分泌物质的囊泡横切Two secretory substances are distributed within the cell, one is like linear clump and the other is in spherical particle, and the inset shows the transverse section of vesicles containing secretions; D: 胞外分泌物质Exocrine substances; E: 靠近微绒毛的分泌囊泡Secretory vesicles near microvilli; F: 中肠冰冻切片免疫荧光染色, 荧光显示细胞质基质被染成绿色(微管蛋白) Frozen section of midgut stained with immunofluorescence, cytoplasmic matrix being strained in green (tubulin); G: 中肠冰冻切片免疫荧光染色, 荧光染色显示柱状上皮细胞的分泌物质(三角形指示)和微绒毛被染成红色Frozen section of midgut stained with immunofluorescence, secretory substances (indicated by triangle) and microvilli of columnar cells being stained in red; H: 中肠冰冻切片荧光染色叠加图Merge of immunofluorescent image of frozen section of midgut. 星号示基底膜。The asterisk indicates the basal lamina. M: 线粒体Mitochondrion; Mi: 微丝Microfilaments; Mv: 微绒毛Microvilli; Nu: 细胞核Nucleus; RER: 粗面内质网Rough endoplasmic reticulum; Sj: 间壁连接Septate junction; V: 囊泡Vesicle.
2.4.2再生细胞:细胞位于中肠上皮组织基部,其体积较小、细胞质有限,通常为多个细胞聚集在一起形成龛(图3: C-D)。细胞内细胞核大,近椭圆形,占据细胞大部分空间(图5: A-B)。细胞核仁明显,异染色质团块清晰,细胞质中可见大量球状或长条状线粒体和少量低电子密度囊泡(图5: B)。细胞基部细胞膜与基膜紧密相连。当柱状上皮细胞凋亡或损伤时,再生细胞分化形成柱状上皮细胞,通过细胞逐渐延长至端部到达管腔。
2.4.3内分泌细胞:细胞通常位于再生细胞龛的外围,其核位于细胞端部,比大多数的柱状上皮细胞的卵圆形核小得多(图5: C)。细胞质向下延伸到基膜,向上不到达管腔,内有大量的中等电子密度的分泌颗粒(图5: D-E)。

图5 庭疾灶螽中肠的再生细胞及内分泌细胞Fig. 5 Regenerative cells and endocrine cells of the midgut of Tachycines asynamorusA: 龛中的再生细胞Regenerative cells in the niche; B: 细胞核周围的线粒体Mitochondria around the nucleus; C: 中肠内分泌细胞与再生细胞基部相邻Endocrine cells adjacent to the basal region of the regenerative cells; D: 内分泌细胞基部与发达的基膜相连Basal region of endocrine cell connecting with developed basal lamina; E: 内分泌细胞包含的分泌颗粒Secretory granules in endocrine cells. Bl: 基膜Basal lamina; Lm: 纵肌Longitudinal muscle; M: 线粒体Mitochondrion; Nu: 细胞核Nucleus.
2.5 中肠基底超微结构
无细胞结构的基膜是一类细胞外基质,位于上皮组织与结缔组织之间。庭疾灶螽的基膜厚度约1~2 μm,较发达,与纵肌紧密相连(图6: A-C)。当管腔中未被食物充满时,基膜向上皮组织基部内折。当管腔充满食物时,基膜会展开为食物提供更大的空间。基膜外周由纵肌与环肌内外嵌合而成,为肠道的蠕动提供机械支持(图6: D-E)。在肌肉层周围散布着的气管细胞可帮助肠道进行呼吸交换气体(图6: F)。

图6 庭疾灶螽的中肠基底Fig. 6 Basal region of the midgut of Tachycines asynamorusA: 中肠基膜Basal lamina of the midgut; B: 分布在基膜下的纵肌Distribution of the longitudinal muscle below the basal lamina; C: 中肠基膜的细节图Detailed image of basal lamina of the midgut; D: 中肠环肌肌肉层Circular muscle layer of the midgut; E: 形状不规则的肌细胞核Irregularly-shaped muscle nucleus; F: 中肠浆膜及气管细胞Serosa and trachea cells of midgut. Bl: 基膜Basal lamina; Cm: 环肌Circular muscle; Lm: 纵肌Longitudinal muscle; Nu: 细胞核Nucleus; Tr: 微气管Tracheole.
免疫荧光染色显示,基膜经鬼笔环肽-罗丹明染成红色(图4: G)。由于鬼笔环肽特异性地与细胞中的微丝结合,故推测基膜上有微丝的成分可能对组织起到支撑作用。中肠基膜经过PAS染成红色(图3: D),推测基膜含有复合糖成分。
2.6 马氏管显微结构
庭疾灶螽的马氏管数目220~260根,基部着生于中后肠的交界处,端部封闭游离于血腔中(图1: C)。马氏管的管壁为具有大的细胞核的单层上皮细胞,有3~5个大型细胞围绕管腔。马氏管靠管腔内为一层长而密集的微绒毛(图7: A-B)。荧光染色观察显示, 马氏管管腔内的微绒毛被鬼笔环肽-罗丹明染成红色(图7: C-F)。
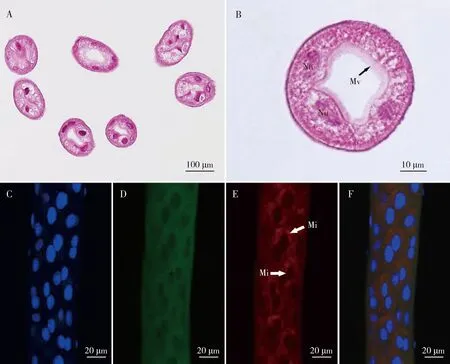
图7 庭疾灶螽马氏管的显微结构Fig. 7 Microstructure of the Malpighian tubules of Tachycines asynamorusA: 马氏管横切石蜡切片H.E.染色Paraffin transection of Malpighian tubules stained with hematoxylin-eosin; B: 马氏管横切石蜡切片细节图Detailed image of paraffin transection of Malpighian tubules; C: 马氏管免疫荧光染色图, 荧光染色显示马氏管细胞核被染成蓝色(DAPI) Immunofluorescent image of Malpighian tubules, the nuclei being stained in blue (DAPI); D: 马氏管免疫荧光染色图, 荧光染色显示细胞质基质被染成绿色(微管蛋白) Immunofluorescent image of Malpighian tubules, cytoplasmic matrix being stained in green (tubulin); E: 马氏管免疫荧光染色图, 荧光染色显示马氏管管腔微绒毛被染成红色(微丝) Immunofluorescent image of Malpighian tubules, the microvilli in the lumen of Malpighian tubules being stained in red (microfilaments); F: 马氏管免疫荧光染色叠加图Merge of immunofluorescent image of Malpighian tubules. Mi: 微丝Microfilaments; Mv: 微绒毛Microvilli; Nu: 细胞核Nucleus.
2.7 马氏管超微结构
透射电镜下观察到马氏管从外向内依次由无细胞结构的基膜、单层细胞结构的上皮组织、中央管腔等组成。微绒毛位于端部细胞膜,排列致密,伸入中央管腔内(图8: A-B)。
单层细胞的细胞核大且明显,位于细胞中央区域,含有异染色质形成的小块(图8: D)。相邻的细胞间存在间壁连接(图8: A)。
细胞基部的细胞膜(基底膜)高度内褶形成膜质迷路,可占细胞近1/3,其中分布大量的形状各异的线粒体与少量的空泡,并垂直于基膜(图8: C)。细胞质内分布着电子致密的同心圆球晶体,同心圆间很有层次感,规则排列。马氏管细胞的外表面由肌肉层和气管环绕(图8: E-F)。
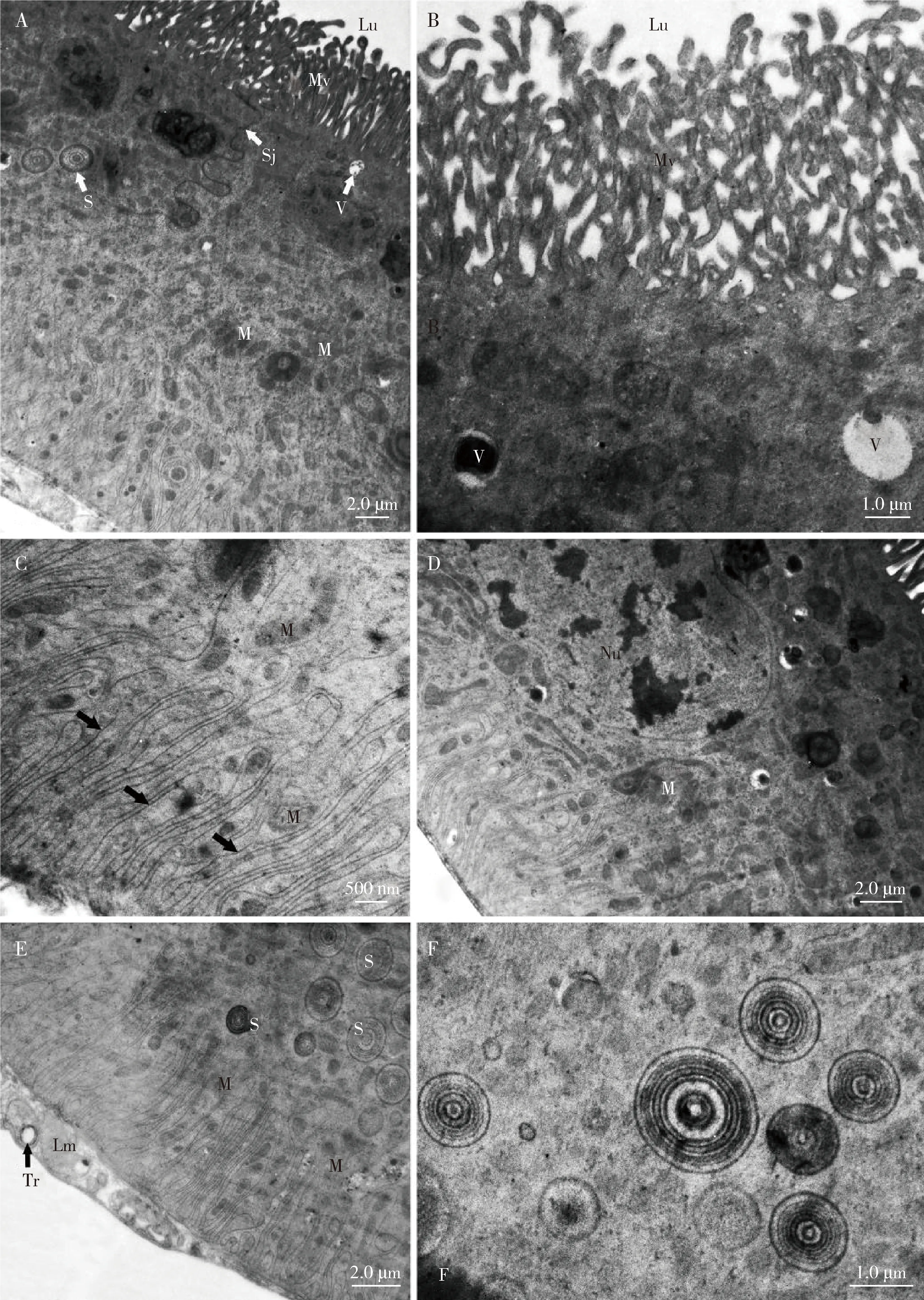
图8 庭疾灶螽马氏管的超微结构Fig. 8 Ultrastructure of the Malpighian tubules of Tachycines asynamorusA: 相邻细胞间存在间壁连接,细胞质顶端部具有纤细的微绒毛Septate junction between two neighboring cells and apical cytoplasm with slender microvilli; B: 分布在细胞质中电子密度不等的囊泡Vesicles with varying electron densities distributed in the cytoplasm; C: 上皮细胞基部细胞膜深度内折形成膜迷路(黑色箭头指示)Basement membrane of epithelial cell with typical multiple infoldings, generating a membrane labyrinth indicated by black arrow; D: 细胞核形状不规则,含有异染色质形成的小块Nucleus in irregular shape, containing several small patches of heterochromatin; E: 细胞的端部和基底部区含有大量的同心圆球晶体Numerous concentrically layered spherites in the apical and basal region of the cell; F: 电子密度不等的同心圆球晶体Concentrically layered spherites with varying electron densities. Lm: 纵肌Longitudinal muscle; Lu: 管腔Lumen; M: 线粒体Mitochondrion; Mv: 微绒毛Microvilli; Nu: 细胞核Nucleus; S: 球晶体Spherite; Sj: 间壁连接Septate junction; Tr: 微气管Tracheole; V: 囊泡Vesicle.
3 讨论
3.1 中肠的结构与功能
昆虫多样化的食性与其消化道的结构和功能相适应(Nation, 2011)。其消化道由单层上皮细胞构成,外围有基底膜和横纹肌。通常认为消化道是从口到肛门的管状结构,其纵贯于血腔中央,具有运送、消化食物和吸收营养物质的作用,是个体完成生长、发育和繁殖的基础(Chapman, 2013)。昆虫前肠和后肠起源于外胚层,前肠在储存食物和消化方面具有相当重要的作用,但在吸收方面作用不显著;而后肠具有回收水和排便的作用(Chapman, 2013)。
中肠是消化道中唯一内胚层起源的部分,中肠胃盲囊的形态、数量、大小和着生位置因昆虫种类不同而异,主要功能是增加中肠的表面积,有利于产生消化酶和吸收营养物质(Chapman, 2013),这可能与昆虫的食性相关。直翅目中不同科的胃盲囊数量具有差异,如蝗科具6个胃盲囊,且每个分成前后2叶;在螽斯科和蟋蟀科的一些种类中,胃盲囊着生于中肠前端,数量仅有2个,其基部较宽,远端部较窄(刘玉素和庐宝廉, 1955; Biagioetal., 2009; Çakici and Ergen, 2012; Lietal., 2018)。本研究发现庭疾灶螽有3个胃盲囊,其表面排布有网状气管(图2: A),表明胃盲囊有很多需氧的代谢活动,如大分子营养物质的吸收、消化酶的分泌等。
中肠细胞积极参与消化酶的产生和分泌以及营养物质的吸收,因此中肠是消化道的功能核心(Chapman, 2013)。中肠主要有两种分化后的细胞类型,即柱状上皮细胞和内分泌细胞。几个再生细胞聚集在一起形成再生细胞龛。此外,中肠细胞中还有一种杯状细胞,在鳞翅目、毛翅目、蜉蝣目及襀翅目的消化道中常见(Chapman, 2013)。本研究结果表明,庭疾灶螽中肠的显微和超微结构观察没有发现杯状细胞,该种中肠细胞共有3种类型:再生细胞、柱状上皮细胞和内分泌细胞。
再生细胞主要补充受损或凋亡的柱状上皮细胞和内分泌细胞(Nation, 2011)。再生细胞位于肠壁细胞基部的再生细胞龛内,在龛中有一个特殊的微环境保持它们的干细胞特征(Illa-Bochaca and Montuenga, 2006)。飞蝗Locustamigratoria的再生细胞聚集在中肠上皮基部的龛内,并沿胃盲囊和中肠分布。蟋蟀Achetadomesticus中肠再生细胞是上皮的干细胞,增殖密集。位于龛内顶端的细胞具有上皮特征并分化为上皮细胞,那些退化的上皮细胞则从基膜分离而消失,中肠细胞的再生是连续的(Rost-Roszkowska, 2008)。膜翅目中Meliponaquadrifasciataanthidioides成虫的再生细胞龛的位置是等距的,若虫细胞龛则不规律分布在柱状上皮细胞间;在变态过程中,幼虫中肠上皮退化,在幼虫的再生细胞分化过程中形成新的成虫中肠上皮(Martinsetal., 2006)。本研究结果表明,庭疾灶螽的再生细胞龛位于中肠上皮基部,数量多且排布较整齐,紧贴于基膜内侧(图3: C-D);通过超微结构发现再生细胞包含数量多、形状不规则的线粒体(图5: B)。再生细胞有丝分裂后,进一步分化出柱状上皮细胞和内分泌细胞。
柱状上皮细胞是所有昆虫中肠的主要细胞类型,承担消化酶的产生和营养的吸收,顶膜上有微绒毛,增加了中肠的表面积。蝗虫Abracrisflavolineata中肠的柱状上皮细胞微绒毛被糖萼覆盖,微绒毛间有深色颗粒,细胞质内大量的高尔基体靠近细胞核,粗面内质网丰富(Maranaetal., 1997)。本研究发现,庭疾灶螽中肠柱状上皮细胞微绒毛密集,有利于与肠腔物质交换(图4: A-B)。胞内大量与分泌相关的细胞器表明柱状上皮细胞的分泌活动强烈。柱状上皮细胞有两种明显的分泌颗粒(图4: C),一种是线团状颗粒,荧光染色与超微结构表明该颗粒由微丝包裹有固定形态的胞内物质而形成,推测该分泌颗粒是为了将细胞内的多余结构排出;另一种是电子密度很高的球状颗粒,可能包含消化酶。对于2种物质的具体组成及其作用,还需进一步验证。
内分泌细胞有两种不同的形态特征:开放型内分泌细胞与中肠腔直接接触,基膜位于中肠基板上;闭合型内分泌细胞不会延伸到整个中肠上皮,因此也不会到达管腔,与基膜接触面小(Cacciaetal., 2019)。 内分泌细胞的分泌特性体现在其细胞质中有许多电子致密颗粒(Bonellietal., 2019)。 Neves等(2003)发现膜翅目的Meliponaquadrifasciataanthidioides中肠细胞中闭合型内分泌细胞体积较小,有不规则的分泌颗粒,电子密度低于开放型内分泌细胞;开放型内分泌细胞的分泌颗粒及线粒体主要位于细胞质基部。 直翅目昆虫中,Illa-Bochaca和Montuenga(2006)应用免疫细胞化学和免疫荧光技术发现飞蝗Locustamigratoria中肠内分泌细胞通常位于细胞龛的外围,增殖速度慢,其细胞核比大多数上皮细胞的核小,通过细长的细胞质延伸到达中肠腔。本研究发现,庭疾灶螽只有一种位于再生细胞龛外围的闭合型内分泌细胞(图5: C),没有膜迷路,在基底细胞区有较大的分泌颗粒聚集,推测它们通过基膜排出内容物到血腔中。内分泌细胞在昆虫中肠中所起的作用尚不清楚,但与脊椎动物一样,这些细胞可能在调节肠道活动方面起作用(Lehaneetal., 1996)。
本研究结果表明,庭疾灶螽的中肠基底层由基膜、肌肉以及其他结缔组织组成。在光学显微镜下观察,基膜位于再生细胞与肌肉层之间,呈线状,结构清晰完整(图3: C-D)。基膜较厚,约1~2 μm,PAS染色呈现明显的红色,表明基膜包含大量的复合糖成分,荧光染色观察到基膜上具有微丝(图4: G)。电子显微镜下,基膜由靠近细胞膜的基板和外围的网板构成,基板的质地均匀且连续,网板的结构类似于网格状,较疏松(图6: C)。致密且收缩性不强的基板为中肠上皮细胞提供了稳定的机械支撑,质地疏松的网板为中肠的收缩运动提供了结构基础。推测基膜的基板与网板的互补结构通过对中肠上皮细胞的支撑作用为肠道的蠕动提供保障。
综上所述,庭疾灶螽中肠符合昆虫中肠的基本结构,中肠壁结构从外到内依次为浆膜、环肌、纵肌、基底膜和上皮层。但胃盲囊的数量与直翅目蝗虫、蟋蟀皆不同,为3个。作为消化道的功能核心,庭疾灶螽中肠细胞类型有再生细胞、柱状上皮细胞和内分泌细胞,没有杯状细胞;中肠3类细胞的着生位置与直翅目其他种相似,同时我们首次观察到了庭疾灶螽中肠内分泌细胞,推测是封闭式中肠内分泌细胞;结合荧光染色,我们认为,庭疾灶螽中肠柱状上皮细胞的线团状颗粒由微丝包裹。另外庭疾灶螽中肠基膜发达,包含微丝蛋白和复合糖成分。
3.2 马氏管的结构与功能
在许多昆虫物种中,马氏管基部连接到中肠和后肠的交界处,远端游离于血腔中,如半翅目长红锥蝽Rhodniusprolixus,双翅目黑腹果蝇Drosophilamelanogaster,蛩蠊目的Grylloblattacompodeiformis等(Jarial, 1990; Martinietal., 2007; Gautam and Tapadia, 2014)。庭疾灶螽的马氏管属于此类,外围丰富的微气管说明马氏管具有需氧的新陈代谢。但是在鳞翅目、脉翅目和鞘翅目的一些昆虫中,马氏管端部伸入直肠,这种马氏管类型被称为隐肾系统(Pachecoetal., 2014; Nocellietal., 2016)。
马氏管的数量、细胞学和组织学结构以及球晶在不同昆虫群体中可能有很大差异,这些差异被认为是重要的,可以用作分类特征。马氏管的数量因昆虫种类不同而异,半翅目有4根马氏管,鳞翅目有6根马氏管,蜻蜓目有50~60根马氏管,直翅目马氏管数目可能超过200根(Savageetal., 1956; Jarial, 1990; Pachecoetal., 2014; Amutkanetal., 2015)。庭疾灶螽的马氏管数目超过200根,说明其代谢速率远高于其他昆虫。
庭疾灶螽马氏管腔内侧的大量微绒毛增加了细胞游离面,提升了物质传递的效率。马氏管细胞基部的基底迷路具有大量线粒体,表明物质从血淋巴到小管上皮的运输过程需要能量,可能与马氏管从周围体液中吸收代谢废物有关,类似动物肾脏的远曲小管。
马氏管作为排泄器官,细胞摄取液体、离子和废物等进入马氏管腔。此外昆虫对废物的另一种策略是将其存储在体内而不排出,因为尿酸是不溶于水的,它可以很容易地存储而不会与其他生理过程起冲突(Klowden, 2013; Nocellietal., 2016)。庭疾灶螽马氏管细胞胞质内可见大量颗粒和同心圆球晶体,主要在靠近基底迷路,在微绒毛侧很少见到,马氏管管腔内也看不到同心圆颗粒的存在,推测这些球状晶体是一种储存排泄物。对直翅目螽斯科的Leptophyesalbovittata的研究发现,马氏管细胞球状晶体的化学成分中含有大量的碳、磷和锰,这些离子是昆虫发育和细胞中正确完成代谢活动所必需的,马氏管在离子的运输和动态平衡的调控中可能起着重要的作用(Amutkan-Mutlu and Suludere, 2020; Amutkan-Mutluetal., 2021)。广布于欧洲的驼螽科穴螽Troglophilusneglectus马氏管细胞球晶体的结构和化学组成随着生命周期而变化,为生命周期的关键过程提供重要矿物质;Troglophiluscavicola在滞育期的不同阶段马氏管细胞的自噬体的丰富度具有差异,且雌性高于雄性(Lipovšeketal., 2009; Lipovšeketal., 2016)。庭疾灶螽马氏管的球晶体中是否包含代谢活动所必需的离子还需要进一步研究。
马氏管外围的数层基膜,其作用可能是将气管和肌肉与马氏管相连。气管可以为马氏管细胞供氧,马氏管外围的肌肉表明马氏管具有一定的伸缩性,促进血淋巴的流动,增强代谢。
综上所述,本研究结果表明,庭疾灶螽的中肠和马氏管的结构与其他直翅目物种基本相似。本研究阐明了庭疾灶螽中肠和马氏管的形态和组织结构,为进一步研究其结构上与生活环境、食性的适应关系奠定了基础,为理解直翅目的进化关系提供了资料。
猜你喜欢
空间电子技术(2022年4期)2022-09-26
科学导报·学术(2020年47期)2020-11-17
中国保健营养(2019年9期)2019-10-20
学习与科普(2019年4期)2019-09-10
智慧少年(2018年9期)2018-10-29
名作欣赏(2017年32期)2017-11-28
人生十六七(2017年6期)2017-06-06
小小说月刊·下半月(2016年7期)2016-05-14
星火(2013年2期)2013-07-18
中华少年(2009年9期)2009-09-14
