
角倍蚜越冬若蚜的生物学特性
2022-07-12 02:44:10魏洪媛丰国蕊邵淑霞杨子祥
昆虫学报 2022年6期
许 鑫, 魏洪媛, 丰国蕊, 邵淑霞, 杨子祥
(中国林业科学研究院高原林业研究所, 国家林业和草原局资源昆虫培育与利用重点实验室, 昆明 650224)
角倍蚜Schlechtendaliachinensis属于半翅目(Hemiptera)蚜科(Aphididae)绵蚜亚科(Eriosomatinae)五节根蚜族(Fordini),其生活史为异寄主全周期型,以若蚜在侧枝匐灯藓Plagiomniummaximoviczii等藓类植物上越冬,春季羽化为有翅蚜,迁飞到盐肤木Rhuschinensis上取食寄生并形成虫瘿,称为角倍。秋季虫瘿自然爆裂,瘿内有翅蚜形成并迁飞到藓类植物上,开始新的生活史循环(张广学和钟铁森, 1983; Blackman and Eastop, 2007)。角倍是经济价值最高的五倍子种类,其产量占五倍子总量的70%以上,对五倍子生产具有决定性的作用,广泛应用于化工、医药、食品、电子和矿冶等行业(张宗和, 1991; 李志国等, 2003; 杨子祥, 2011)。
国内外对角倍蚜的生活史和各虫态的生物学特性进行过研究,但对越冬若蚜的研究较少。角倍蚜的世代包括春迁蚜、性蚜、干母、干雌、秋迁蚜和越冬若蚜等(唐觉, 1976),其中干母和干雌在瘿内生活和繁殖,封闭的虫瘿为其提供了稳定的营养来源、较高的湿度和较好的保护,种群数量随着虫瘿体积的增加而快速增加,每个虫瘿内的干雌可达5 000~6 000头(Shaoetal., 2013),但秋季虫瘿成熟,第3代干雌发育成秋迁蚜迁飞到藓上后,由于失去了虫瘿的保护,加上秋冬季的低温和降雨等,秋迁蚜及其生殖的若蚜死亡率极高,仅1龄若蚜的死亡率就高达90%以上(赖永祺等, 1990),因此秋迁蚜的数量虽然很大,但春迁蚜数量常常不足,造成角倍产量低且不稳定,严重制约了五倍子生产的发展。赖永祺等(1990)对越冬若蚜的生物学特性及数量变动进行了研究,发现了越冬若蚜的死亡率极高,但未对越冬若蚜的转移取食和形态特征等进行调查。
本实验应用数码显微系统对角倍蚜越冬若蚜的行为习性、形态特征、种群动态和群体龄期结构等进行系统研究,并结合寄主侧枝匐灯藓藓层厚度和覆盖率的变化,探寻越冬若蚜种群数量变动的影响因子,旨在阐明角倍越冬若蚜的生物学习性,为角倍蚜种虫培育技术的改进提供科学依据。
1 材料与方法
1.1 供试材料
1.1.1供试昆虫: 供试昆虫角倍蚜和冬寄主侧枝匐灯藓均采自四川省峨眉山市川主镇荷叶村(29°37′N, 103°24′E,海拔1 010 m)。8月上旬将侧枝匐灯藓定植于湿润的无纺布表面,10月上旬采集成熟并自然爆裂的角倍蚜虫瘿,让秋迁蚜自然迁飞到无纺布藓块上(1 m2藓块平均约4万头),保湿培育。从10月至翌年3月,每月中旬定期采集田间的有蚜藓块,带回实验室,用数码显微系统连续观察和记录越冬若蚜的发育情况。
1.1.2仪器:数码显微系统:Keyence VHX-1000(日本);体式显微镜:Olympus SZX16(日本)。
1.2 角倍蚜生物学特性和形态观察
1.2.1秋迁蚜的行为观察:取秋迁蚜刚迁飞后的藓块,裁剪成10 cm×10 cm大小,置于数码显微系统下,连续观察和记录秋迁蚜在藓枝上的搜寻、爬行和繁殖行为。
1.2.2越冬若蚜的行为观察及形态特征:从越冬若蚜在藓上爬行和寻找藓枝取食开始,连续观察若蚜的搜寻、取食、泌蜡、蜕皮和移动等特性,直到春迁蚜产生。从藓枝上定期挑取各龄若蚜(若蚜龄期根据蜡球内蜕皮的数量判断),利用数码显微系统测量各龄若蚜的体长和体宽等。每个龄期30个重复。蜡球内和蜡球外若蚜数量的统计标准是:蜡球完全成形且从外部看不见虫体的,统计为蜡球内若蚜;没有或者未完全成形,从外部可看见虫体的,统计为蜡球外若蚜。
1.2.3越冬若蚜的种群动态和龄期分布:从10月至翌年3月,每月中旬在藓块上设置10 cm×10 cm的样方,按5点取样法取样,每个样点取4 cm2,统计样点上所有若蚜的龄期和数量,分析龄期组成和数量动态。重复3次。
1.2.4蜡球内若蚜数量调查:从10月至翌年3月,每月中旬在藓块上设置10 cm×10 cm的样方,在每个样方内随机选取20个完整的蜡球,统计每个蜡球内的若蚜数量。重复3次。
1.2.5藓层覆盖率和厚度调查:从10月至翌年3月,每月中旬在藓块上设置10 cm×10 cm的样方,随机测量不同部位的藓层厚度,每个样方测10次,取平均值;并用网格法测量每个样方的藓层覆盖率。重复3次。
1.3 数据分析
采用生物统计学分析软件SPSS 22.0进行相关性分析以及使用邓肯氏新复极差检验(Duncan’s new multiple range test, DMRT)法进行差异显著性分析。若蚜死亡率的计算公式为:(当月中旬若蚜数-下月中旬若蚜数)/当月中旬若蚜数×100%。
2 结果
2.1 角倍蚜秋迁蚜及越冬若蚜的行为习性
秋迁蚜产越冬若蚜:角倍蚜秋迁蚜迁飞到侧枝匐灯藓藓枝,约1 h开始产越冬若蚜,可连续产蚜或间隔产蚜。产若蚜前,秋迁蚜在藓枝上呈“之”字形爬行,并不停地上下摆动触角,寻找合适的产蚜部位。一旦找到合适的产蚜部位,便停止爬行,通过腹部的蠕动产出若蚜,时间持续约30 s。初产若蚜为淡黄色,外包一层薄薄的半透明黏膜,以腹部末端黏在藓上,其体位方向与在母体内一致。随后,被黏膜包裹的若蚜通过触角、前足、中足和后足的扭动和前后摇晃,黏膜从若蚜头部顶端向腹部末端滑落并与虫体分离,约20 min后若蚜可自由爬行,寻找合适的藓枝取食。初产若蚜爬行能力较强,24 h后,若蚜体表便可看到明显的白色蜡粉,但仍可爬行,通常产出3 d后固定取食,即使有轻微惊扰也不移动。
越冬若蚜的取食和转移:角倍蚜越冬若蚜通常在拟叶基部,即拟叶与拟茎的夹角处分泌棉絮状的蜡丝包裹自身,并持续分泌蜡丝形成疏松的蜡球,在蜡球内取食并越冬(图1: A)。在越冬期间若蚜通常不移动,但遇到不良的环境条件,如藓枝生长不良、拟叶枯黄等,若蚜会在蜡球侧面或顶端破开一个洞口,爬出并寻找新的藓枝取食,并重新分泌蜡丝形成新的蜡球,但在移动过程中约有80%的若蚜会死亡。表明若蚜虽然具有转移取食并重新形成蜡球的能力,但转移过程中死亡率较高。

图1 角倍蚜越冬若蚜在侧枝匐灯藓上形成的蜡球Fig. 1 Wax balls formed by the overwintering nymphs of Schlechtendalia chinensis on the moss Plagiomnium maximovicziiA: 蜡球中的角倍蚜若蚜A nymph in a wax ball of P. maximoviczii; B: 角倍蚜若蚜群集取食后侧枝匐灯藓枯黄Withered mosses after being fed by a colony of S. chinensis nymphs.
越冬若蚜泌蜡形成蜡球:角倍蚜若蚜通常在靠近地面的新萌发的匐匍枝上取食并形成蜡球,通常1头若蚜形成1个蜡球,但当秋迁蚜迁飞数量大,若蚜数量较多时,会形成紧密相连的密集蜡球,甚至1个蜡球内有2头若蚜,导致藓枝上的拟叶皱缩、卷曲,甚至枯萎(图1: B);或者当匐匍枝直立生长,离开土表时藓枝也通常会生长不良,造成若蚜转移,或在蜡球内死亡。蜡球内的若蚜可持续分泌蜡丝,使蜡球保持白色;当蜡球颜色变黄或变暗时,挑开蜡球会发现若蚜通常已经死亡,因此可通过蜡球的颜色来判断若蚜的生存状态。
2.2 角倍蚜越冬若蚜和春迁蚜形态特征
角倍蚜越冬若蚜各龄期之间的体型大小差异较大(表1)。随着龄期的增加,越冬若蚜体长和体宽显著增加,同时触角和口器的长度也在增加,其中4龄若蚜的体长和体宽分别为1龄若蚜的2.18和2.01倍。除了1龄若蚜的触角为4节,2-4龄若蚜和春迁蚜的触角均为5节。虽然1龄若蚜的口器长度最短,但其占体长的比达到0.28,可达后足基节,而2-4龄若蚜的口器仅达中足基节或前中足间。
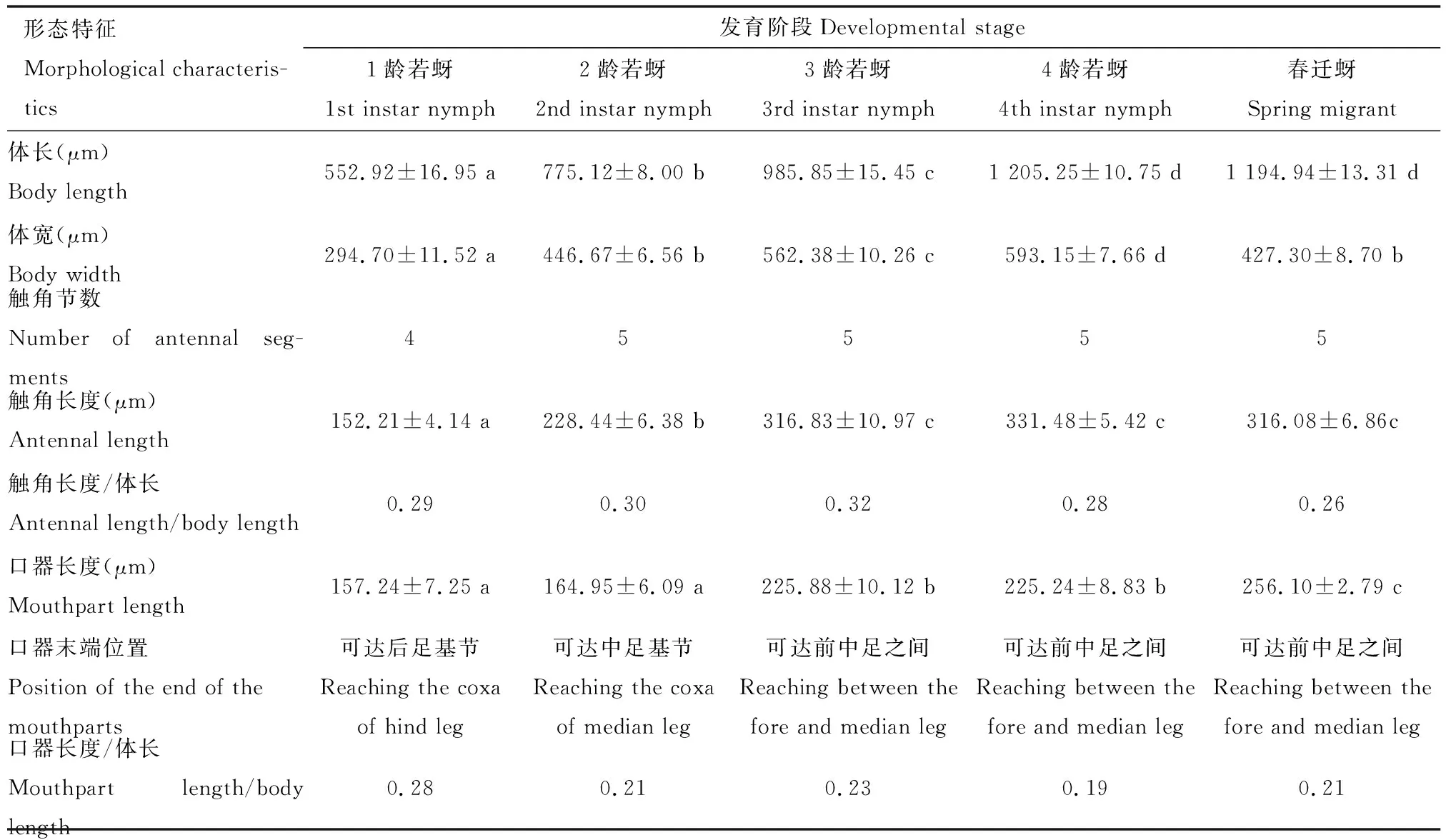
表1 角倍蚜越冬期间各发育阶段的形态特征比较Table 1 Comparison of the morphological characteristics of Schlechtendalia chinensisat different developmental stages during overwintering
角倍蚜1-4龄越冬若蚜的体色是由淡黄色逐渐加深至深褐色(图2)。1龄若蚜初产时为淡黄色,然后逐渐加深至黄褐色,2龄若蚜为黄褐色,3龄若蚜为深黄褐色,4龄若蚜为深褐色,局部黑色。1、2龄若蚜中后胸没有翅芽,而3和4龄若蚜中后胸有明显可见的翅芽,因此翅芽的发育可作为判断若蚜龄期的依据。3龄若蚜翅芽为深黄褐色,而4龄若蚜为深褐色,3和4龄若蚜翅芽末端都伸至腹部1-2节,并且胸部背面隆起。春迁蚜体色为黑褐色,胸部背面隆起,2对翅透明,翅脉明显,停息时双翅呈屋脊状斜盖于体背,翅长约为体长的1.5倍。

图2 角倍蚜越冬若蚜不同龄期的形态特征Fig. 2 Morphological characteristics of overwintering nymphs of Schlechtendalia chinensis of different instars
2.3 角倍蚜越冬若蚜的种群数量动态
角倍蚜越冬期间若蚜的死亡率极高,从10月中旬的13.10万头/m2急降至翌年3月中旬的1.05万头/m2(图3),总死亡率高达91.98%,每月的死亡率分别为45.29%(月均温15.02℃)、26.28%(月均温8.43℃)、49.84%(月均温4.27℃)、46.54%(月均温9.05℃)和25.88%(月均温13.00℃),其中10月中旬至11月中旬、12月中旬至翌年2月中旬的死亡率高,均超过45%,11月中旬至12月中旬和2月中旬至3月中旬的死亡率相对较低,但仍超过25%(图4)。
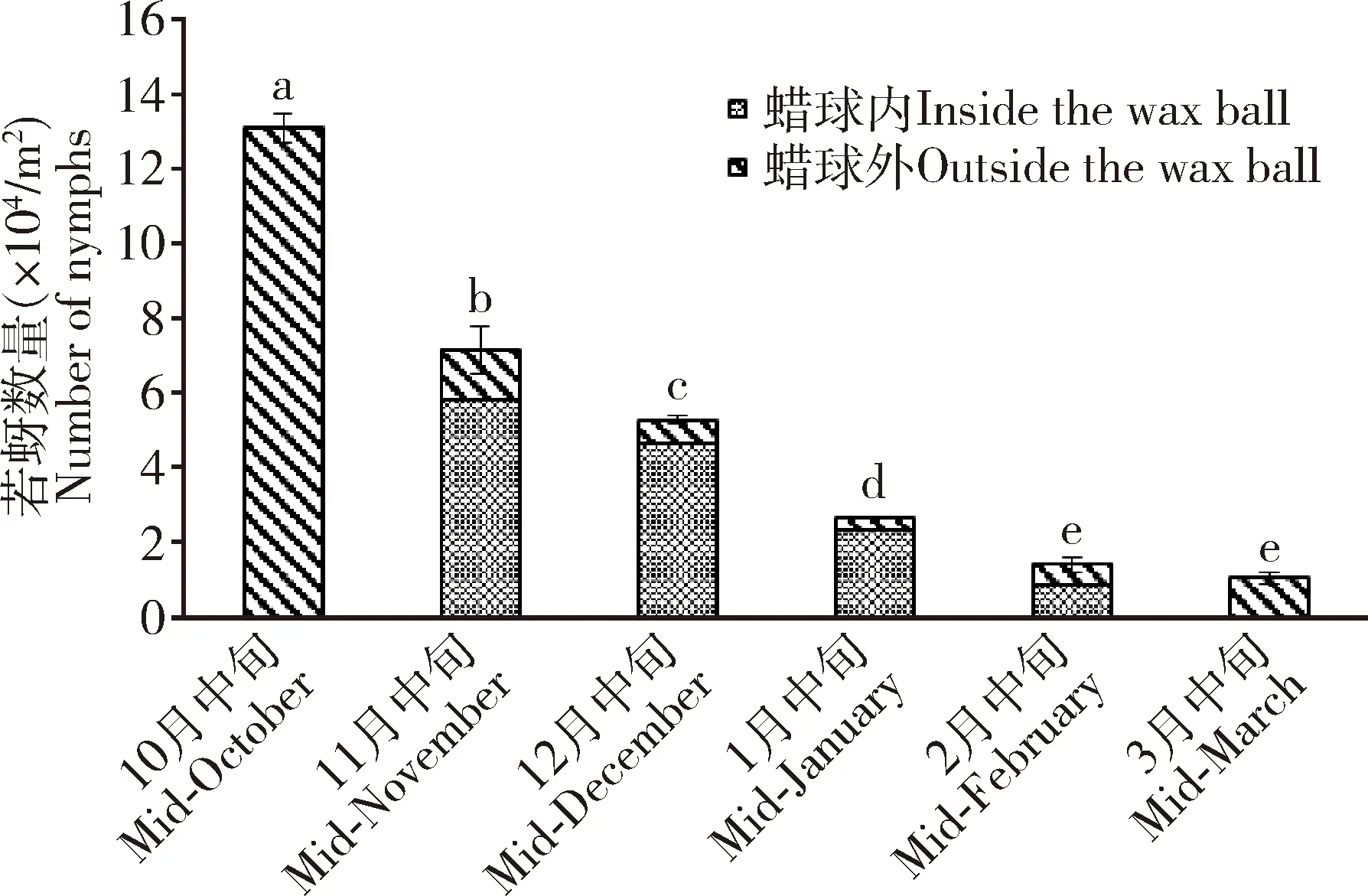
图3 越冬期间角倍蚜若蚜种群数量动态Fig. 3 Population dynamics of Schlechtendalia chinensisnymphs during overwintering图中数据为平均值±标准误;柱上不同小写字母者表示不同时间的若蚜数量差异显著(P<0.05, DMRT法)。Data in the figure are mean±SE. Different lowercase letters above bars indicate significant difference in the number of nymphs at different time (P<0.05, DMRT method).

图4 越冬期间角倍蚜若蚜的死亡率Fig. 4 Mortalities of Schlechtendalia chinensisnymphs during overwintering
角倍蚜越冬若蚜大部分时间在蜡球内渡过,但仍有少部分若蚜位于蜡球外。从10月中旬至翌年3月中旬,每月蜡球外若蚜数占总若蚜数的比例分别为100.00%, 18.84%, 12.30%, 10.69%, 37.65%和98.41%,蜡球外若蚜数呈现“两头高、中间低”的变化(图3),其中10月中旬若蚜仍位于蜡球外(虽有蜡丝包裹但蜡球尚未完全成形,仍可看见若蚜),11月中旬到翌年2月中旬绝大部分若蚜位于蜡球内,且1月中旬蜡球外若蚜比例最低,此时也是一年中气温最低的时期,2月中旬若蚜准备迁飞,部分已经转移到蜡球外,3月中旬几乎全部从蜡球内出来,即将迁飞。11月中旬到翌年3月中旬,蜡球外若蚜数量占总若蚜数的比例与冬季气温变动呈正相关(相关系数为0.882),表明蜡球内外若蚜数量的变化与冬季气温变化和若蚜的个体发育与迁飞习性密切相关。
每个蜡球内通常有1头角倍蚜若蚜,但由于受不良环境条件、天敌和若蚜密度等的影响,也会出现蜡球内的若蚜为0或2头的现象。从10月中旬至翌年3月中旬,每个蜡球内有2头若蚜的比例很低,为0~3.33%;每个蜡球内有1头若蚜的数量逐渐减少,但2月中旬至3月中旬急剧下降;与此相对应的是:每个蜡球内有0头若蚜(空蜡球)的数量逐渐增加,但2月中旬至3月中旬急剧上升(图5)。反映了越冬期间若蚜极高的死亡率,其中2月中旬至3月中旬,空蜡球的比例从43.33%快速上升到96.67%,表明若蚜大部分转移到蜡球外,准备迁飞。
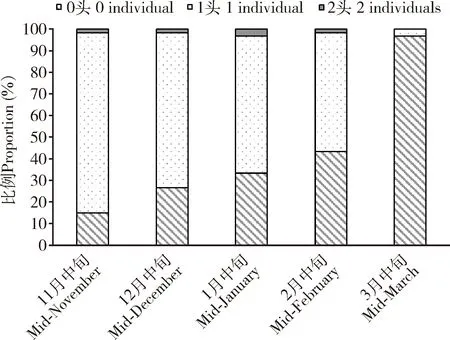
图5 角倍蚜若蚜越冬期间每个蜡球内若蚜数量的比例Fig. 5 Proportions of the number of Schlechtendaliachinensis nymphs in a wax ballduring overwintering
2.4 角倍蚜越冬若蚜的龄期分布
角倍蚜越冬若蚜个体间的发育进度很不整齐,并与当地的气温变化密切相关。10月中旬全部为1龄若蚜,至11月中旬,有86.05%发育成2龄若蚜,而至12月中旬仅有18.01%的发育成3龄若蚜,到1月中旬,3龄若蚜的比例增加到40.25%,表明这2个月若蚜的发育进度较10-11月明显放缓;到2月中旬,有68.24%的若蚜发育成4龄若蚜,若蚜的发育进度较 11月-翌年1月明显加快;到3月中旬,
88.89%的若蚜已经发育成春迁蚜,开始迁飞(表2)。可以看出,越冬若蚜在11月中旬至翌年1月中旬发育进度较慢,而在10月中旬至11月中旬和1月中旬至3月中旬发育相对较快。这与当地的气温变化规律基本一致,即11月中旬至翌年1月中旬的气温较低,10月中旬至11月中旬、1月中旬至3月中旬的气温逐渐升高的变化规律相一致。

表2 越冬期间角倍蚜若蚜不同龄期的百分比Table 2 Percentage of Schlechtendalia chinensis nymphs at different instars during overwintering
2.5 藓层覆盖度和厚度变化
角倍蚜若蚜越冬期间侧枝匐灯藓的厚度和覆盖率逐步增加。从10月中旬到翌年3月中旬,藓层厚度从1.13 cm增加到1.52 cm,增加了34.51%,除11月中旬相对于10月中旬显著增加(P<0.05)外,其余相邻两月之间的厚度差异不显著(P>0.05)(图6: A);藓的覆盖率从91.81%增加到99.69%,增加了8.58%,其中从10月中旬到11月中旬增加量最多,达到4.64%,占总增加量的58.88%,以后4个月间仅有小幅增加,相邻两月之间的覆盖率差异不显著(P>0.05)(图6: B)。

图6 角倍蚜若蚜越冬期间侧枝匐灯藓藓层厚度(A)和覆盖率(B)变化Fig. 6 Changes in the thickness (A) and coveragerate (B) of the moss Plagiomnium maximoviczii duringthe overwintering of Schlechtendalia chinensis nymphs图中数据为平均值±标准误;柱上不同小写字母表示不同时间的侧枝匐灯藓藓层厚度和覆盖率差异显著(P<0.05, DMRT法)。Data in the figure are mean±SE. Different lowercase letters above bars indicate significant difference in the thickness and coverage rate of the moss P. maximoviczii at different time (P<0.05, DMRT method).
3 讨论
目前,生产上采用的无土植藓方法培育五倍子蚜种虫,已经取代了传统的土壤植藓养蚜,成为五倍子人工培育的主要生产方式。本研究基于无土植藓养蚜方法,发现越冬若蚜的总死亡率为91.98%,累计死亡率呈“S”型变化。而赖永祺等(1990)基于传统的土壤植藓养蚜方法,发现越冬若蚜的总死亡率为98.50%,累计死亡率呈“对数曲线”型变化。可见,基于两种不同的植藓方法,越冬若蚜的死亡率和数量动态变化有差异显著,其中无土植藓方法优于传统土壤植藓方法。这是由于无土植藓方法更有利于保持藓块湿度的稳定,提高了若蚜的存活率(查玉平等, 2019)。
越冬若蚜如此高的死亡率,可能与其特殊的生物学特性和生活微环境有关。首先,越冬若蚜自身没有形成完善的抗寒机制。多数昆虫在长期进化过程中,形成了休眠、滞育或以卵越冬等生态适应策略,以抵御冬季低温、干旱或冰冻等不良环境条件(匡海源, 1964)。角倍蚜以若蚜越冬,没有观察到休眠或滞育的现象(赖永祺等, 1990; 张传溪等, 1997),而是靠若蚜分泌蜡丝形成蜡球的方式越冬。其次,越冬时间长,仅靠蜡球不足以适应温湿度等的变化。越冬若蚜的蜡球比较疏松,虽有一定的防水、保温功能(Weietal., 2020),但只能部分隔离降水和露水,不足以应对越冬期间长达5个月的温度和湿度的激烈变化或冰冻等,因此在越冬期间,月平均死亡率均超过25%,尤其是在蜡球未完全形成的10-11月,以及温度最低的12月-翌年2月,月平均死亡率更是超过了45%。最后,由于越冬若蚜不滞育,需要从藓枝上持续取食,但藓类属于低等植物,形态结构简单,抗旱能力弱,尤其是中后期藓层厚度和覆盖度的增加,藓枝很容易由于缺水或营养不足而枯萎,迫使若蚜离开蜡球转移取食,在转移过程中由于缺乏蜡球保护而大量死亡。
针对越冬若蚜死亡率高的问题,在五倍子生产过程中,可以通过有针对性的技术措施,减少降水和温湿度变化对若蚜生存的影响,从而提高若蚜的存活率和五倍子的产量。例如在10-11月蜡球形成过程中和2-3月若蚜离开蜡球迁飞前,对培育若蚜的藓块适时覆盖薄膜避雨,避免雨水对若蚜的影响,提高若蚜存活率;在若蚜越冬期间,通过控制藓上若蚜密度、覆盖遮阳网和薄膜、适时补水等方法,注意调控藓枝的正常生长,防止藓枝过密和过厚,减少若蚜的转移,提高若蚜的存活率,从而提高五倍子产量。
猜你喜欢
建筑材料学报(2024年2期)2024-03-06 08:56:02
浙江农业科学(2024年2期)2024-02-27 15:03:24
湖南城市学院学报(自然科学版)(2022年6期)2022-11-14 01:31:12
今日农业(2021年21期)2021-11-26 05:07:00
农村农业农民·B版(2018年2期)2018-03-22 08:47:22
农村.农业.农民(2018年4期)2018-01-23 02:07:28
现代园艺(2017年11期)2017-06-28 11:32:51
四川水泥(2016年8期)2016-07-18 12:06:31
中国农业文摘-农业工程(2016年5期)2016-04-12 05:38:13
现代农业(2016年4期)2016-02-28 18:42:06
