
柑橘大实蝇成虫的梳理行为
2022-07-12 02:44何章章桂连友张智亮周仁迪华登科刘文茹姜振宇汤建涛
昆虫学报 2022年6期
何章章, 桂连友,*, 杨 璇, 田 甜, 张智亮, 周仁迪,华登科, 刘文茹, 姜振宇, 汤建涛
(1. 长江大学农学院, 湖北省农林病虫害预警与调控工程技术研究中心, 湖北荆州 434025; 2. 长江大学计算机科学学院,湖北荆州 434025; 3. 湖北省农业科学院, 农业质量标准与检测技术研究所, 武汉 430064)
梳理行为是动物维持健康生存的一种非常重要的活动,其包含了身体表面护理的所有形式(Szebenyi, 1969; Goldmanetal., 1972; Pageetal., 1990; Newland, 1998; Rogers and Newland, 2000; Mooringetal., 2004)。作为地球上数量最多、分布最广的动物群体,昆虫会经常性地进行自我梳理(Jander, 1966),其用于梳理的器官包括足(Farish, 1972; Böröczkyetal., 2013)、口器(Farish, 1972)、翅(Zack, 1978)和腹部(Jander and Jander, 1978)等。昆虫梳理行为的目的是通过清除自身表面的覆盖物、分泌物、寄生虫、病原体等,保持自身感觉的可靠和灵敏,增加自身对农药的抗药性、收集身体上附着的花粉粒作为食物、以及预防感染(Zhangetal., 2020)。目前,对昆虫梳理行为的研究主要集中于梳理行为的类型(Basibuyuk and Quicke, 1999)、梳理时间(Qiaoetal., 2018)、频次占比(魏朝明等, 2006)、梳理行为的功能(Russoetal., 2020)及调控(Yanagawaetal., 2020; Yildiz, 2020)等。
昆虫梳理行为的研究方法主要有肉眼观察(魏朝明等, 2006; Zhukovskayaetal., 2013)和录像视频观察等(何章章等, 2018),这2种方法均颇为耗时和费力。因此,Zhang等(2020)开发了一种基于计算机视觉识别技术的柑橘大实蝇Bactroceraminax成虫梳理行为检测与统计系统,该系统是利用背景与目标提取方法分离出目标,并与基于时空上下文提取的特征进行融合,使用卷积神经网络(convolutional neural network, CNN)模型对融合后的特征图像进行检测的针对柑橘大实蝇成虫的梳理行为规律设计的一种新的统计方法。
柑橘大实蝇是主要分布于我国的危害柑橘类果实的重要害虫,也是对外检疫对象之一(汪兴鉴和罗禄怡, 1995; Guietal., 2011; Xiaetal., 2018)。柑橘大实蝇成虫于橘园中出土羽化后,初羽化的成虫会离开橘园一段时期进行营养补充,待生殖腺逐渐发育成熟,之后再回到橘园(汪兴鉴和罗禄怡, 1995; 罗杰等, 2016),于柑橘幼果上产卵为害。我们前期研究发现初羽化柑橘大实蝇成虫会从橘园迁移到附近的板栗林中,并在板栗雄花花序上取食,从而进行营养补充(Heetal., 2019, 2020),其后我们又通过录像观察法研究了柑橘大实蝇成虫在中国板栗雄花花序上的取食行为,发现其取食过程中会对自身的口器、足和翅等进行频繁的梳理,由此认为柑橘大实蝇成虫的梳理行为对其正常的取食有着十分重要的意义(何章章等, 2018)。因此,通过采用录像技术和基于计算机视觉识别技术的柑橘大实蝇成虫梳理行为检测与统计系统,对柑橘大实蝇成虫的梳理行为的特征图谱、单次平均耗时和频次占比进行分析,以明确柑橘大实蝇成虫梳理行为规律,研究结果可为挖掘防控柑橘大实蝇的新靶标,制订有效的害虫绿色防控策略提供理论依据。
1 材料与方法
1.1 供试虫源
2019年从湖北松滋市陈店镇(30°18′N, 111°77′E)未采取防控措施的柑橘园采集柑橘大实蝇虫果,带回长江大学昆虫生态学实验室,埋在河沙(厚度5~7 cm)中,幼虫化蛹后,分期分批收集蛹,转移到盛有河沙(厚度5~7 cm)的塑料盆中(直径17 cm,高11 cm),保持土壤湿度20%左右。收集羽化后的成虫,置于养虫笼(35 cm×35 cm×35 cm)中饲养,饲喂的食物是酵母菌浸粉(纯味型,青岛海博生物技术有限公司)和蔗糖(化学纯,北京康普汇维科技有限公司)按1∶3的质量比混匀的混合物(何章章等, 2018)。以饲养的8日龄成虫作为供试虫源。
1.2 实验地点
实验于湖北省荆州市长江大学生态实验室进行,室内温度为22℃,相对湿度为65%,光照为自然光。
1.3 梳理行为观察
梳理行为观测系统由摄像机(图1: C)、实验平台和固定结构(图1: D) 3部分组成(华登科等, 2016)。实验采用 Sony 摄像机(DX-AX60, Sony, Exmor RS CMOS成像器,帧数30 fps,像素92.1万)由铁架台和三脚架固定。透明的塑料培养皿(35 mm×20 mm)由一个圆盘状的底和一个盖组成(图1: A, B),除了不能飞行,成虫在培养皿内可以自由活动。拍摄时,首先需用双面胶带(广东省东莞市旭生包装材料有限公司)将坐标纸(25 cm×17 cm)平整粘贴在垂直于地面的墙上,以作为标尺刻度(1 cm),然后利使用双面透明胶带将透明的塑料培养皿底粘贴于坐标纸的中心,最后将1头饱食的柑橘大实蝇成虫置入培养皿中,盖上培养皿盖,并用透明胶带固定培养皿盖。培养皿盖对应的外壁上方垂直空间即为摄像机的拍摄位点。试验重复20次,雌雄试虫各10头。录像时间为9:00-10:00时,时长1 h。

图1 柑橘大实蝇成虫梳理行为观察平台Fig. 1 Observation platform of grooming behaviorsof Bactrocera minax adultsA: 坐标纸粘在墙上Graph paper is stuck to the wall; B: 培养皿粘在坐标纸Petri dishes are glued to graph paper; C: 录像机Video cassette recorder; D: 观察平台Observation platform.
1.4 梳理行为视频处理
本研究在Zhang等(2020)将柑橘大实蝇的梳理行为分为6种类型的基础上,将其足的梳理行为细分为了前足、中足和后足梳理3种类型;将雌成虫的腹部梳理行为细分为了产卵器和腹部梳理2种类型,这样柑橘大实蝇雌雄成虫分别有9和8种梳理行为,另外增加1种非梳理行为。与Zhang等(2020)将柑橘大实蝇的梳理行为分为6种类型一样,均利用Adobe Premiere Pro CC 软件(Adobe公司),通过人工观察分析,获取各种类型梳理行为的特征图像,再采用计算机视觉识别技术的柑橘大实蝇成虫行为检测与统计系统(Zhangetal., 2020)处理分析影像文件,获取成虫各种类型梳理行为的频次和时间等参数。
1.5 数据分析
单个梳理行为单次平均耗时(s)=某一梳理行为的总时间/某一梳理行为的总次数;
连续2种梳理行为频次占比(%)=某连续2种梳理行为次数/所有连续2种梳理行为的总次数;
单个梳理行为频次占比(%)=某一梳理行为发生总次数/所有单一梳理行为发生的总次数。
所有数据使用DPS数据处理系统进行分析。柑橘大实蝇成虫各种梳理行为类型之间的单次平均耗时和频次占比差异采用单因素方差分析(One-way ANOVA)、Tukey氏多重比较;柑橘大实蝇雌雄成虫之间的单个梳理行为的单次平均耗时和频率占比差异、以及雌雄成虫之间的梳理和非梳理行为总时间的差异均采用Student氏t检验分析;成虫梳理行为与非梳理行为之间的总时间差异采用配对t检验分析。
2 结果
2.1 梳理行为特征图谱
柑橘大实蝇的雌雄成虫梳理行为分别可分为9和8种类型(表1;图2)。两者均有前足、后足、复眼、触角、口器、腹部、翅和中足的梳理,雌成虫还多了梳理产卵器行为。柑橘大实蝇成虫梳理行为分为前、中和后3个部分,前部由触角、复眼、口器和前足组成,依靠前足梳理;中部为中足,依靠后足或者前足梳理;后部分由腹部、翅、产卵器和后足组成,依靠后足梳理。

表1 柑橘大实蝇成虫梳理行为与非梳理行为Table 1 Grooming and non-grooming behaviors of Bactrocera minax adults

图2 柑橘大实蝇成虫的梳理行为特征图谱Fig. 2 Ethogram of grooming behaviors of Bactrocera minax adults小图中字母为梳理行为代号(同表1)。Letters inside figures represent the codes of grooming behaviors as in Table 1.
前部梳理行为:成虫梳理前足分2种类型,一种类型是左右前足向上抬起,其胫节、跗节和前跗节相互摩擦,此过程中,两前足的胫节、跗节和前跗节中的至少一个部分保持密切接触,同时前后互相摩擦;另一种类型是一只前足静止,另一只前足沿着静止的前足的腿节至前跗节方向上单向移动刮擦。成虫梳理后足方式和过程与梳理前足方式和过程相同,分为2种类型,一种为相互摩擦,另一种为一只后足摩擦另一只后足。成虫梳理复眼时,头部向下低垂,前足向上抬起弯曲,利用同侧的前足的胫节沿触角至口器的方向单向移动刮擦复眼,也可同时利用同侧的前足刮擦同侧的复眼。成虫梳理触角时,头部向下低垂,前足向上抬起弯曲,利用同侧的前足的跗节和前跗节沿着触角基部至顶端方向单向移动刮擦触角,也可同时利用同侧的前足刮擦同侧的触角。成虫梳理口器时,口器向外伸出,前足向上抬起弯曲,两前足的跗节和前跗节夹住口器,沿着口器基部到顶端方向单向移动刮擦口器。成虫在梳理前足、复眼、触角和口器时,多以4只足(2只中足和2只后足)或5只足(1只前足、2只中足和2只后足)支撑其身体平衡。
中部梳理行为:成虫梳理中足分2种类型,一种类型是利用前足梳理中足,此过程中,两前足与一只中足的胫节,跗节和前跗节相互刮擦;另一种类型是利用后足梳理中足,此过程中,两后足与一只中足的胫节,跗节和前跗节相互摩擦。成虫在梳理中足时,3只足抬起,另外的3只足(1只中足和2只前足、或1只中足和2只后足)支撑其身体平衡。
后部梳理行为:成虫梳理腹部时,后足向上抬起弯曲,利用两后足的跗节和前跗节沿着胸部至腹部的方向单向移动刮擦腹部。成虫梳理翅的过程有多种方式:(1)两后足向后弯曲,其跗节和前跗节抬起分别置于同侧翅的背面,沿胸部至腹部的方向,以刮擦的方式同时梳理左右翅的背面部分;(2)成虫后足抬起,两后足的跗节与其翅的边缘接触,沿胸部至腹部的方向刮擦其翅的边缘;(3)成虫后足弯曲,两后足的胫节和跗节沿头部至腹部的方向以刮擦的方式梳理翅的腹面;(4)成虫一只后足在同侧的翅的背面,另一只后足在其翅的腹面,两足夹住一侧的翅,沿头部至腹部的方向以刮擦的方式梳理翅的两面。成虫的梳理产卵器时,后足抬起并向后伸展,分别位于产卵器基节的两侧,通过其跗节和前跗节以刮擦的方式梳理产卵器。成虫在梳理后足、腹部、翅和产卵器时,多以4只足(2只前足和2只中足)支撑其身体平衡。
2.2 梳理行为总时间与非梳理行为总时间之间的差异
柑橘大实蝇的雌成虫非梳理行为的总时间(2 349.76±137.87 s)显著长于其梳理行为的总时间(1 250.24±137.87 s)(t=3.99,df=9,P<0.05);雄成虫非梳理行为的总时间(2 533.79±181.87 s)显著长于其梳理行为的总时间(1 066.21±181.87 s)(t=4.03,df=9,P<0.05)。雌雄成虫之间非梳理行为的总时间(t=0.81,df=18,P>0.05)和梳理行为的总时间(t=0.82,df=18,P>0.05)均没有显著差异。雌、雄成虫梳理行为总时间分别占其非梳理行为总时间的53.2%和42.1%。
2.3 单个梳理行为的单次平均耗时和频次占比差异
在柑橘大实蝇的雌成虫(F=25.36,df=8,72,P<0.05)和雄成虫(F=26.90,df=7,63,P<0.05)中的各类型梳理行为之间的单次平均耗时均存在显著差异(表2)。其中,除了雌雄成虫梳理翅的行为和梳理中足行为的单次平均耗时均分别显著长于其梳理前足、后足、复眼、触角、口器、腹部行为的单次平均耗时,雄成虫梳理翅行为的单次平均耗时显著高于其梳理中足行为的单次平均耗时外,成虫其余梳理行为类型之间的单次平均耗时均无显著差异。除了柑橘大实蝇雌虫梳理产卵器行为外,雌雄成虫之间的同一类型的梳理行为的单次平均耗时均无显著差异(前足:t=0.85,df=18,P>0.05; 后足:t=0.39,df=18,P>0.05; 复眼:t=1.13,df=18,P>0.05; 触角:t=1.85,df=18,P>0.05; 腹部:t=1.01,df=18,P>0.05; 口器:t=0.82,df=18,P>0.05; 翅:t=1.89,df=16,P>0.05; 中足:t=0.55,df=18,P>0.05)。
在柑橘大实蝇的雌成虫(F=23.21,df=8,72,P<0.05)和雄成虫(F=24.13,df=7,63,P<0.05)中的各类型梳理行为之间的频次占比均存在显著差异(表2)。其中,除了柑橘大实蝇的雌、雄成虫梳理前足行为的频次占比均分别显著高于其梳理后足、复眼、触角、口器、腹部、翅、产卵器(仅雌成虫)和中足行为的频次占比,雌成虫梳理触角行为的频次占比显著高于其产卵器行为的频次占比,雄成虫梳理触角行为的频次占比显著高于其梳理翅行为的频次占比外,成虫其余梳理行为类型之间的频次占比均无显著差异。除了梳理产卵器行为之外,柑橘大实蝇雌雄成虫之间相同梳理行为类型的频次占比均无显著差异(前足:t=0.06,df=18,P>0.05; 后足:t=0.43,df=18,P>0.05; 复眼:t=1.90,df=18,P>0.05; 触角:t=0.39,df=18,P>0.05; 腹部:t=1.48,df=18,P>0.05; 口器:t=1.25,df=18,P>0.05; 翅:t=1.09,df=18,P>0.05; 中足:t=0.92,df=18,P>0.05)。

表2 柑橘大实蝇雌雄成虫各类型梳理行为之间的平均耗时和频次占比比较Table 2 Comparison of the average time of one grooming and the proportions of frequencies among various typesof grooming behaviors of female and male adults of Bactrocera minax
2.4 连续2种梳理行为的频次占比差异和梳理模型
柑橘大实蝇成虫梳理行为与非梳理行为之间发生转换次序是可逆的。不论雌成虫(图3: A; 表3)或者雄成虫(图3: B; 表4),当其完成任何一种类型的梳理行为后,下一步都可能转换到非梳理行为,雌、雄成虫其转换频次占比的范围分别为7.4%~46.7%和5.2%~61.2%;当雌(图3: A; 表3)、雄成虫(图3: B; 表4)完成非梳理行为后,下一步都可能转换到任何一种梳理行为类型,雌、雄成虫其转换频次占比的范围分别为6.7%~24.6%和8.6%~18.3%。上述转换次序之间的频次占比均不相同,但均超过5%;依照统计学观点,这些转换经常发生。
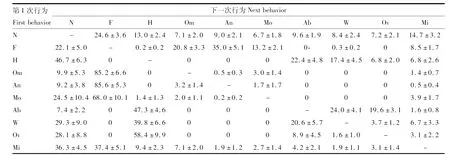
表3 柑橘大实蝇雌成虫连续2种行为(非梳理行为和梳理行为)频次占比(%)Table 3 Proportions of frequencies (%) of two consecutive behaviors (non-grooming behaviors and grooming behaviors)of female adults of Bactrocera minax

表4 柑橘大实蝇雄成虫连续2种行为(非梳理行为和梳理行为)频次占比(%)Table 4 Proportion of frequencies (%) of two consecutive behaviors (non-grooming behaviors and grooming behaviors)of male adults of Bactrocera minax
柑橘大实蝇雌、雄成虫发生其可逆转换类型分别为27个和22个,如梳理前足-梳理触角、梳理触角-梳理前足等;发生其单向转换类型分别为3个和5个,如梳理前足-梳理后足、梳理口器-梳理后足等,且其频次占比均低于5%,依照统计学观点,这些转换偶然发生。
依据统计学小概率原理,即一般认为等于或小于0.05或0.01的概率为小概率事件(秦秉杰, 2019),柑橘大实蝇成虫连续2种梳理行为之间发生次序转换的频次占比以5%作为分界线。柑橘大实蝇成虫连续2种梳理行为之间发生次序转换的频次占比是不相同的(图3; 表5),可分为大于5%, 0~5%和0 3种类型。依照统计学观点,其频次占比超过5%的连续2种梳理行为类型(♀: 21个;♂: 16个)是经常发生的,如成虫梳理前足-梳理复眼等;其频次占比为0~5%的连续2种梳理行为(♀: 22个;♂: 16个)是偶尔发生的,如成虫梳理前足-梳理后足等;其频次占比为0%的连续2种梳理行为(♀: 29个;♂: 24个)不会发生,如梳理前足-梳理腹部等。
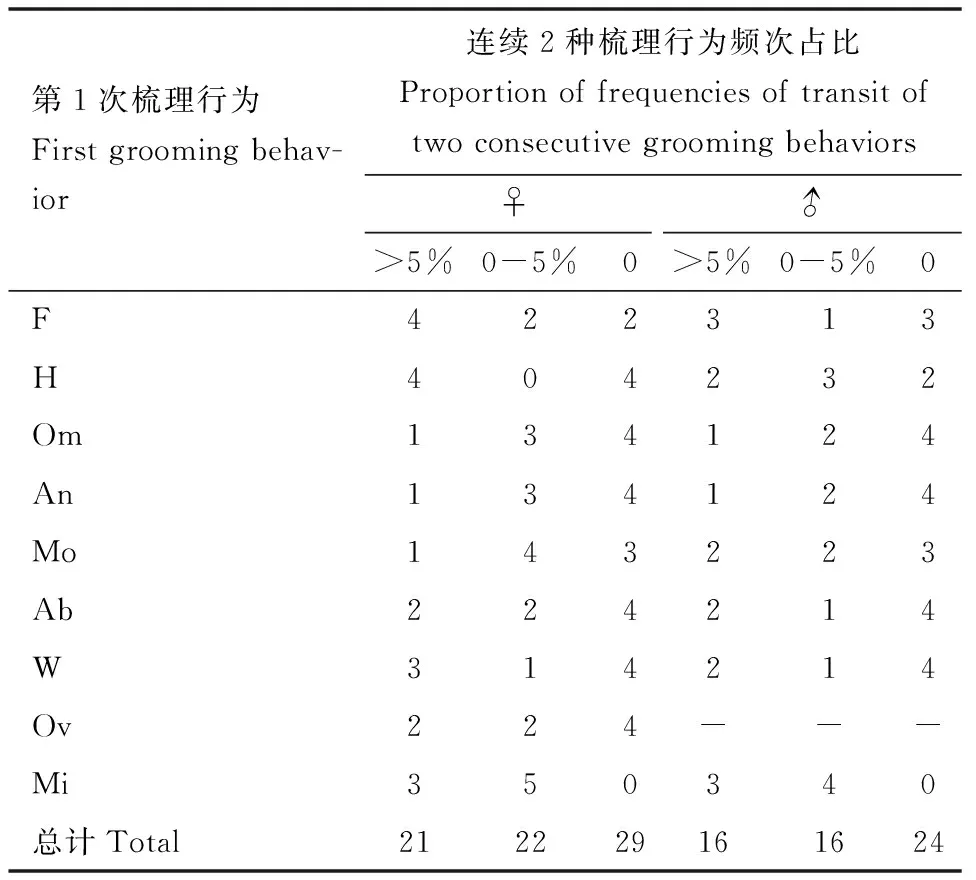
表5 柑橘大实蝇成虫连续2种梳理行为频次占比差异Table 5 Proportions of frequencies of two consecutivegrooming behaviors of Bactrocera minax adults
在柑橘大实蝇成虫梳理行为模型中(图3),假如以连续2种梳理行为中的后一种梳理行为(或包含非梳理行为,下同),作为下一个连续2种梳理行为的前一种梳理行为,可依次类推出连续3种或连续3种以上的梳理行为。如梳理前足-梳理复眼-梳理前足、梳理后足-梳理腹部-梳理翅-梳理后足等。或者说,可通过连续2种梳理行为的连接可以建立整个梳理行为模型。

图3 柑橘大实蝇雌成虫(A)和雄成虫(B)的梳理行为模型Fig. 3 Grooming behavior model of female (A) andmale (B) adults of Bactrocera minax红线表示2种类型的梳理行为之间的转换次序是可逆的,黑线表示2种类型的梳理行为之间的转换次序只能单向的。依照统计学观点分类,实线表示2种类型的梳理行为之间发生转换的频次占比大于5%,代表这2种类型的梳理行为之间的转换经常发生;虚线表示2种类型的梳理行为频次占比为0~5%,代表这2种类型的梳理行为之间的转换偶尔发生。The red line indicates that the two types of grooming behaviors can transit reversibly, and the black line indicates that the two types of grooming behaviors can only be converted in one way. According to statistical point of view, the solid line indicates that the proportion of frequencies of the two types of grooming behaviors is more than 5% (transit often occurs), and the dotted line indicates that the proportion of frequencies of the two types of grooming behaviors is 0-5% (transit occasionally occurs).
3 结论与讨论
柑橘大实蝇雌、雄成虫的梳理行为类型分别为9和8种,两者均具有前足、后足、复眼、触角、口器、腹部、翅、和中足梳理,而雌成虫还有产卵器梳理。除了产卵器梳理行为,双翅目(Szebenyi, 1969)、膜翅目(Basibuyuk and Quicke, 1999; Reberetal., 2011; Hamiduzzamanetal., 2017)、鞘翅目(Valentine, 1973; Santosetal., 2017)、直翅目(Bret and Ross, 1986; Kuriwada and Reddy, 2016)和蜚蠊目(El-Awami and Dent, 1995; Zhukovskaya and Lychakov, 2016)的昆虫都具有这些梳理行为,柑橘大实蝇雌虫特殊的产卵方式,可能是其梳理产卵器的原因。冀雁蔚等(2008)研究发现黄猩猩果蝇Drosophilarelanogaster会梳理胸部,而同为双翅目的果蝇(Richard and Dawkins, 1976)和柑橘大实蝇,却未有此种类型的梳理行为。在泥蜂科总科的昆虫中,只有方头泥蜂亚科(Crabroninae)和短翅泥蜂科(Trypoxylonidae)的昆虫具有梳理胸部的行为(Farish, 1972)。除了梳理类型的不同,不同种类的昆虫梳理姿势和梳理方式也不尽相同。柑橘大实蝇成虫通过后足梳理翅,而蜻蜓目(Jander and Jander, 1978)和螳螂Mantodea(Zack, 1978)则是通过腹部的振动和翅的振动来梳理翅。柑橘大实蝇成虫通过前足梳理触角,而美洲蟑螂Periplanetaamericana和德国小蠊Blattellagermanica除利用前足外,还会通过口器梳理触角(Böröczkyetal., 2013),膜翅目昆虫的前足还具有一个特殊的结构,即触角清洁器(Farish, 1972)。昆虫的梳理行为具有特殊的功能,口器、复眼和触角上遍布着感觉器官(Spruijt, 1992),通过梳理行为来保持这些感觉器官的清洁。蜜蜂可以通过足的梳理,收集身体上附着的花粉粒(Rath, 1999)。柑橘大实蝇成虫通过足的梳理收集身体上附着的蜜露和花蜜等,并以此作为其食物来源。昆虫梳理足来保持其足跗节黏附垫的附着力(Hosoda and Gorb, 2011),梳理翅则是为了清洁和平整鳞片,以减少飞行中的空气阻力(Thelen and Farish, 1977)。昆虫本身会不停分泌新的表皮碳氢化合物,在体表形成疏水层,减少体内水分散失(杨锦锦等, 2015),柑橘大实蝇梳理行为目的之一可能是为了保持体表碳氢化合物的均匀分布,维持身体水分平衡(Böröczkyetal., 2013)。
柑橘大实蝇成虫在饱食的状态下,梳理行为的耗时占成虫活动总时间的1/3,而在取食的过程中,梳理行为的耗时约占活动总时间的1/5(何章章等, 2018),取食过程中的梳理行为的耗时缩短,这与姚松林等(2005)的研究结果类似,其研究发现,饥饿后的刀角瓢虫Serangiumjaponicum雌成虫捕食烟粉虱Bemisiatabaci卵的取食行为的耗时延长, 梳理行为的耗时缩短。柑橘大实蝇的梳理行为是个复杂的过程,若其各个阶段一成不变,则可能无法适应环境的改变。柑橘大实蝇成虫梳理前足的频率最高,前足作为梳理行为的执行器官,保证其本身的清洁是保持触角、复眼和口器清洁的基础。柑橘大实蝇成虫各被梳理器官之间的形态、面积大小、感知重要性、易污染性等均存在不同程度的差异,这也导致了柑橘大实蝇成虫不同类型梳理行为的耗时和频次占比不同。柑橘大实蝇成虫梳理前足的频次占比最高,但梳理翅的单次平均耗时却最长。这表明相较翅而言,在较短的时间内,柑橘大实蝇成虫就能将前足梳理干净。柑橘大实蝇成虫翅的表面积较大,并且成虫梳理翅的部位包括正面、腹面和侧缘,因此,成虫梳理翅的耗时也会相对较长。动物行为一般可分为定型行为(stereotyped behavior)和后天习得行为(learned behavior)。魏朝明等(2006)研究结果发现,果蝇在去翅以后,仍然会做出与正常近似的梳理翅的动作,即所谓的“真空行为”。在培养皿洁净的环境中,柑橘大实蝇成虫也会进行梳理行为,包括但不限于梳理布满感觉器官的头部、前足、后足、腹部、翅、中足和产卵器,这种现象也从侧面说明了柑橘大实蝇成虫的梳理行为可能属于定型行为(Wilson, 1962)。
柑橘大实蝇成虫可以从任何一种类型的梳理行为转为非梳理行为,也可以从非梳理行为转为任何一种类型的梳理行为,这与魏开炬等(2015)的研究结果类似,其研究发现除了无法从排泄行为转到梳理行为,大突肩瓢虫Synonychagrandis的梳理行为和非梳理行为可以相互转换,这种转换可能是昆虫强适应性的表现。成虫的梳理行为与爬行、静息等非梳理行为可交替发生,且柑橘大实蝇成虫梳理行为的次序并不固定,其中存在着较高频次的固定连续转换顺序的梳理行为,如当成虫梳理复眼、梳理触角和梳理口器时,梳理前后均可能梳理前足;梳理腹部、翅和产卵器(雌成虫)时,梳理前后均可能梳理后足,表明不同类型的梳理行为之间具有内在的联系。
本实验通过对柑橘大实蝇梳理行为的研究,对柑橘大实蝇成虫的梳理行为进行了描述、分类和建立梳理行为模型,可为进一步研究药物对行为的影响、梳理行为的遗传等方面为理解昆虫多样性提供基础。也可以通过干扰昆虫的梳理行为(如沉默梳理基因)达到抑制昆虫活动行为,为挖掘防控柑橘大实蝇的新靶标,制订有效的害虫绿色防控策略提供理论依据。
猜你喜欢
湖北农业科学(2022年9期)2022-06-01
中国南方果树(2022年2期)2022-04-15
华人时刊(2020年19期)2021-01-14
小猕猴学习画刊(2019年4期)2019-05-08
扬子江(2019年1期)2019-03-08
新农村(2018年32期)2018-04-02
现代农业科技(2017年13期)2017-08-09
农业科技与装备(2017年1期)2017-06-09
现代园艺(2016年7期)2017-01-09
现代园艺(2016年9期)2016-07-16
