
不同种源青檀种子表型性状的变异分析
2022-07-10 01:30:58沈芷若李子娟王书洁郑思静万志兵孟子恒李昀坤
凯里学院学报 2022年3期
沈芷若,李子娟,王书洁,郑思静,万志兵*,孟子恒,李昀坤
(1.黄山学院,安徽黄山 245041;2.博村林场,安徽黄山 245000)
青檀(Pteroceltis tatarinowii)为榆科青檀属唯一落叶树种,第三纪残遗植物[1].青檀在我国西北、东北、华北和中南地区的19 个省均有分布,常生于石灰岩山地、山谷、溪边,是石灰岩地区造林的先锋树种及钙质土壤的指示植物[2],青檀树皮纤维是我国宣纸的独特制作原料[3].因此,青檀具有很高的经济价值和生态价值,是集材用、药用、造纸、饲料、生态防护诸价值于一体的多用途树种[4-5].
青檀在我国分布范围广泛,在不同分布区地理环境条件的影响及长期遗传变异活动下,形成了许多具有差异的表型特征,即表型性状是基因表达与环境条件共同作用的结果.林木种子的性状具有较稳定的遗传特征,在林木遗传研究上具有重要的价值[6].搞清青檀种子表型变异情况和遗传结构,可获得其变异趋势的一般规律,为种源试验采样和试验设计提供重要依据.汪殿蓓等[7]对湖北大贵寺国家森林公园野生青檀种子形态特征、发芽率进行了观测,结果显示种子均长为4.42 mm,种子均宽为4.11 mm,种子均厚为2.88 mm.唐国梁等[8]对山东省5 个天然群体的青檀种子变异进行研究,显示青檀种子具有丰富的变异性状,主要来源于群体内的变异,其原因可能与其长期野外生长分化、周围环境变化有关.曹忆等[9]对6 个自然青檀居群种子形态特征进行比较分析,结果表明6 个居群的果实和种子的变异程度不同,其中安徽LY 和河南JL 的果实变异程度大,各居群种长、种宽、种厚均达到显著差异.
虽然前人对青檀种子性状地理变异有过类似的研究,但是有采种地较少、种源范围不够广泛的局限性.本研究加大了青檀种子的采集范围,从青檀的分布区中挑选了19个代表性的种源,进行种子表型形态及遗传变异分析,探讨不同地理环境对青檀种子性状变异的影响.了解青檀的遗传多样性及其结构可加深人们对其进化历史及适应机制的认识[10-11],为种源地理类群划分、确定青檀优良种源、开展青檀良种选育及后续种源试验提供数据参考.
1 试验材料与方法
1.1 试验材料种子来源及种源地的自然概况
青檀种子于2020年9月在不同地点采集,共收集到10省16市19个种源地的单株种子,各种源地的自然概况见表1.采种时选择天然成熟母树进行随机取样,所选植株处于亚优势地位,植株间距不少于50 m.采种时分单株进行采种,装袋并标上标签,带回实验室观测,并将种子自然干燥,后置于通风阴凉处储存备用.
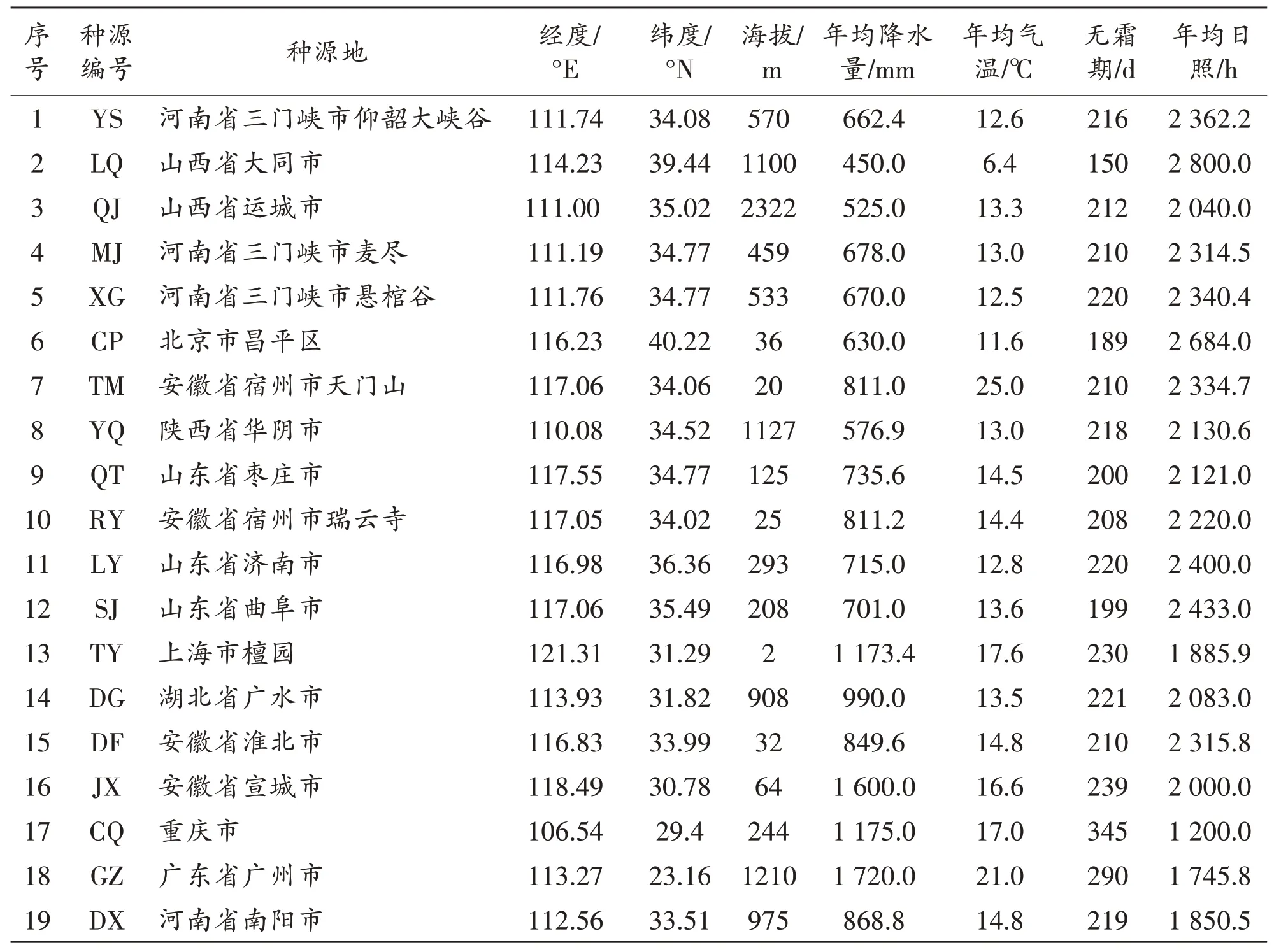
表1 青檀种源地的自然概况
1.2 试验方法
果实阴干,去除果翅后,从每个种源每个单株中随机取出30粒种子,用游标卡尺分别测定种子的长、宽、厚,每粒种子重复3次.所有测量结果均保留2位小数.
1.3 数据处理及分析
试验数据用Microsoft Excel 软件和SPSS18.0软件进行统计分析,对不同种源青檀种子表型性状指标进行方差分析、邓肯多重比较;计算各指标的变异系数,测定各表型性状的离散程度;运用相关分析计算各指标与环境间的相关性;利用聚类分析对19 个采种地的种子性状进行比较并分类.
2 结果与分析
2.1 不同种源青檀种子的表型性状方差分析
对19个青檀种源种子的长、宽、厚等性状进行测定,由表2可知:种长均值为0.38~0.98cm,种长出现最小和最大均值的种源分别是山西大同LQ和北京CP;种宽均值为0.36~1.57cm,种宽出现最小和最大均值的种源分别是上海TY和北京CP;种厚均值为0.20~0.35,种厚出现最小和最大均值的种源分别是北京CP 和山西运河QJ.19 个种源中,北京CP 种源种长、种宽最大,但其种厚最小,种子的表型特征尤为突出.由此看出,各种源中表型性状并不同时减小或增大,各种源种子的表型性状变化趋势不具有一致性.
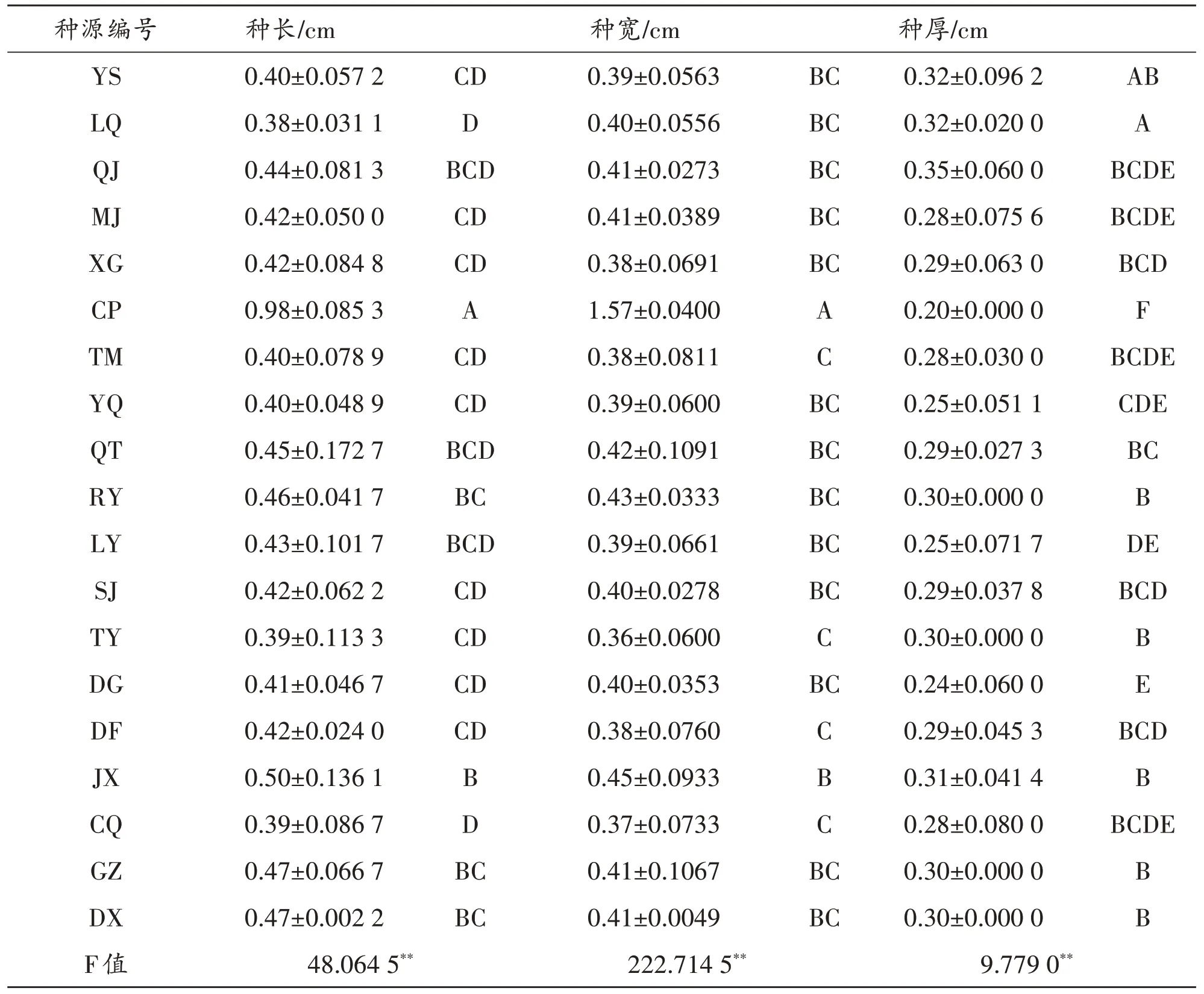
表2 不同种源青檀种子表型性状均值
由表3 可知,不同种源青檀种子在种长、种宽、种厚等性状差异均达极显著水平(P<0.01),表明各种源间种子表型较为丰富,变异较为明显.F值大小顺序为种宽(222.714 5)>种长(48.064 5)>种厚(9.779 0).

表3 不同种源青檀种子表型性状方差分析
2.2 不同种源青檀种子的表型性状的变异分析
变异系数可表现性状的离散程度,19个青檀种源种子表型性状的变异系数见表4.可以看出:在三个表型性状中,变异幅度最大的是种宽,为8.68%;最小的是种厚,为7.78%;总体上表型性状变异幅度不大,说明种子的表型性状具有一定的稳定性.在种源水平上,19 个种源的变异系数平均变化为0.50%~12.52%.南阳DX种源的变异系数最小,为0.05%;宿州TM种源的变异系数最大,为12.52%.将不同种源间的变异系数均值从小到大排序:河南DX、北京CP、宿州RY、曲阜SJ、广水DG、运河QJ、广东GZ、大同LQ、三门峡XG、上海TY、三门峡MJ、宣城JX、淮北DF、三门峡YS、枣庄QT、华阴YQ、济南LY、重庆CQ、宿州TM.以上分析表明:不同种源间及不同种源内各青檀种子表型性状值都表现变异幅度不一致.
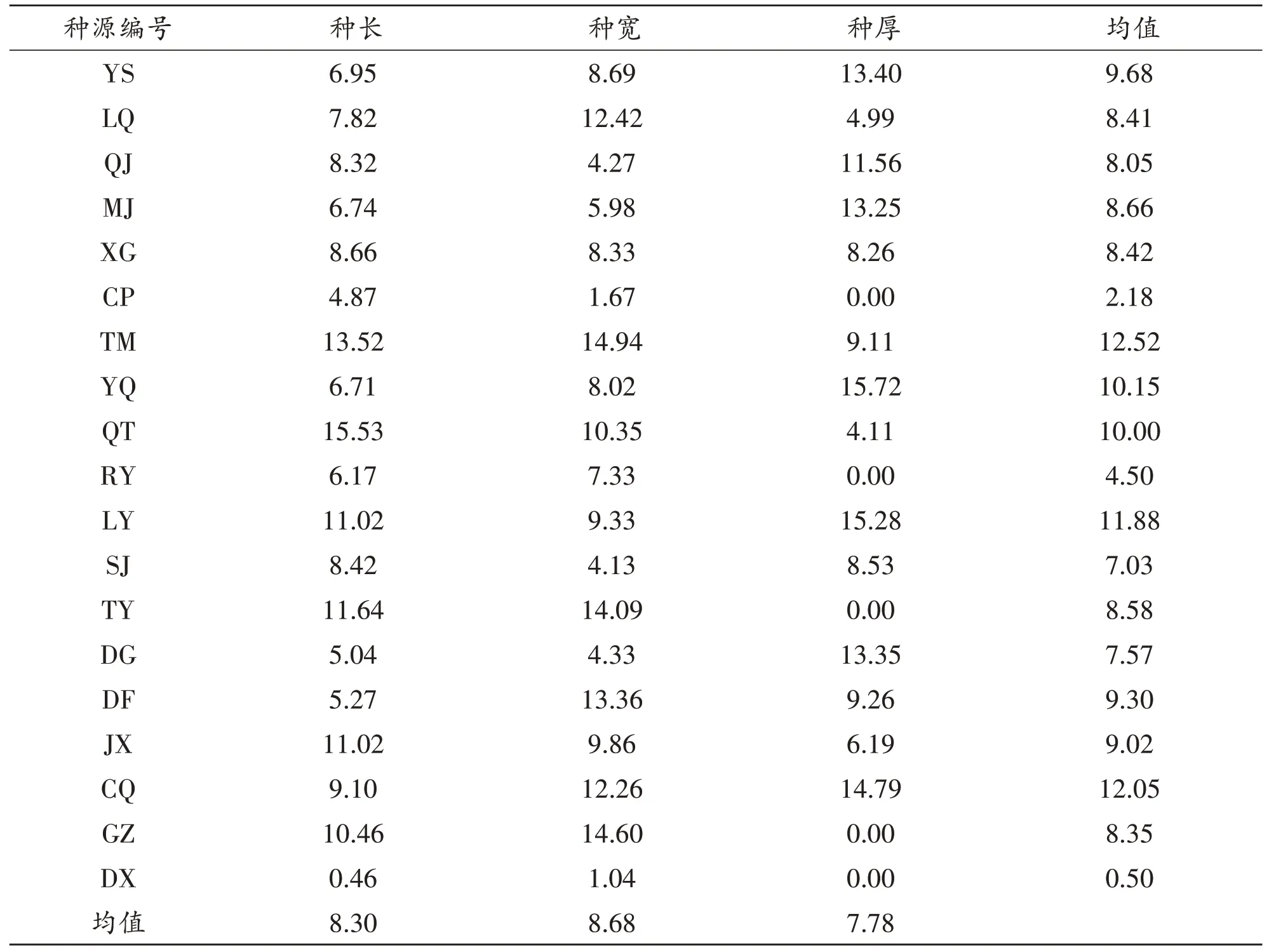
表4 19个青檀种源种子表型性状变异系数/%
2.3 青檀种子表型性状与气象因子的相关性
青檀种子表型性状与7 个气象因子的相关性均不显著,说明气象因子对青檀种子表型变异影响较小,气象因子与这些表型性状存在较弱的相关性或可能没有规律(见表5).种长和种宽都与经度、纬度、年均日照呈正相关,与海拔、年均降水量、年均气温、无霜期呈负相关;种厚与经度、海拔、年均降水量呈正相关,与纬度、年均气温、无霜期、年均日照呈负相关;青檀种子的长、宽、厚与年均气温呈负相关性,即种源地年均气温越低其种子相对较大,表明种源地年均气温低可能更有利于青檀种子的生长发育.
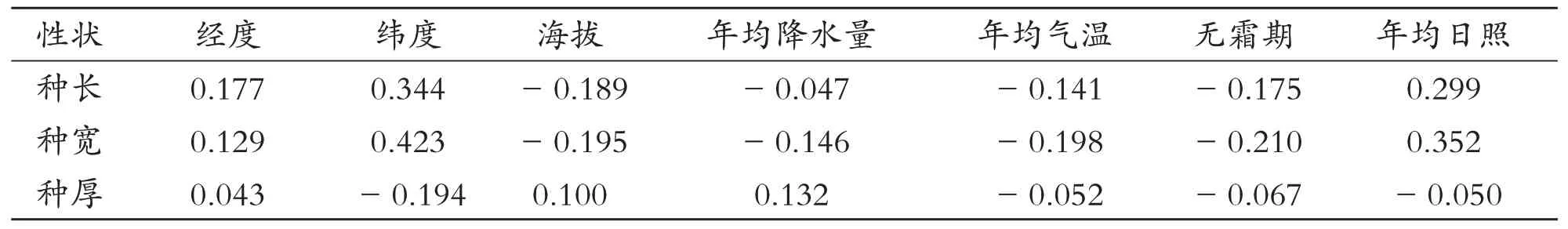
表5 青檀种子表型性状与气象因子的相关系数
2.4 青檀不同种源种子表型聚类分析
根据青檀种长、种宽、种厚3个表型特征,对不同种源青檀种子表型性状进行聚类分析,结果(见图1)可知,以欧式距离5 为阈值,供试19 个青檀种源可分为4 个大类,第Ⅰ大类由北京CP 种源单独组成,其在4 个大类中种子的长度、宽度最大,但是厚度最小,且变异系数较小;第Ⅱ大类由运城LQ 种源单独组成,其在4个大类中种长最小;第Ⅲ大类包括华阴YQ、济南LY、广水DG 等种源,这些地区的种长、种宽、种厚都较小;第Ⅳ大类包括三门峡YS、运城QJ、三门峡MJ、三门峡XG、宿州TM、枣庄QT、宿州RY、曲阜SJ、上海TY、淮北DF、宣城JX、重庆CQ、广东GZ、河南DX 等种源,这些种源的种子表型性状差异不大.
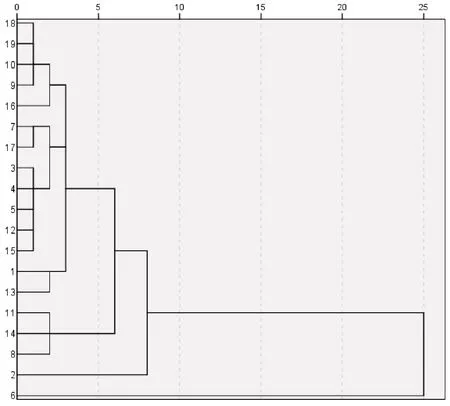
图1 青檀种子表型性状聚类分析
3 结论与讨论
对19个种源青檀种子表型性状进行统计分析,结果表明各种源间种子表型性状变化趋势不具有一致性,但其在种长、种宽、种厚等表型性状均达极显著水平(P<0.01),说明种子性状已经产生了显著性地理变异.F 值大小顺序为种宽>种长>种厚.19个种源间青檀种子表型性状变异系数均值大小顺序是:种宽>种长>种厚,可见,种源内的种子厚较其他性状稳定;南阳DX 种源的变异系数最小,为0.05%;宿州TM 种源的变异系数最大,为12.52%,表明不同种源间种子表型性状值表现变异幅度不一致.青檀种子表型性状与7个气象因子的相关性均不显著,说明气象因子对青檀种子表型变异影响较小,可能种子性状还受到其他因素如栽培方法、遗传信息及生态因子的影响.青檀种子的长、宽、厚与年均气温呈负相关性,表明种源地年均气温低可能更有利于青檀种子的生长发育.通过聚类分析可将供试19个青檀种源划分为4类,第Ⅰ大类仅北京CP种源,其种子的长度、宽度最大,但是厚度最小,且变异系数较小;第Ⅱ大类仅运城LQ种源,其在所有种源中种长最小;第Ⅲ大类包括3个种源,其种长、种宽、种厚都较小;第Ⅳ大类包括14个种源,这些种源的种子表型性状差异不大.
林木的多样性是长期遗传和环境影响的结果[12],不易受环境影响较为稳定的性状,可揭示群体的遗传与变异规律,进而客观评价其遗传多样性[13].表型变异可反应基因型对环境的适应情况[14],青檀各种源间种子表型较为丰富,变异较为明显,为开展优良种源、家系选择及遗传改良工作创造前提条件.种源选择是林木遗传改良的重要手段[15].利用种子性状差异,国内一些专家学者开展了红榉[16]、山核桃[17]、文冠果[18]、沙拐枣[19]等优良种源及家系的选育工作.本文对不同种源青檀种子表型性状指标进行比较,为种源选择奠定前期基础.但这些表型性状指标同时也受环境因素的影响,仅用表型性状指标的差异不能充分反映植物本身的遗传变异,基因型和环境条件哪个影响作用更大,还有待进一步深入研究.
猜你喜欢
种子(2023年7期)2023-09-24 07:40:54
特产研究(2022年6期)2023-01-17 05:05:02
辽宁林业科技(2021年1期)2021-03-16 09:17:08
文萃报·周二版(2019年23期)2019-09-10 07:22:44
天津诗人(2018年4期)2018-11-14 00:49:08
商情(2018年5期)2018-03-28 10:28:14
现代园艺(2017年21期)2018-01-03 06:41:32
文苑·经典美文(2016年8期)2016-05-14 09:31:59
广西林业科学(2016年3期)2016-03-16 05:43:29
中国康复理论与实践(2015年10期)2015-12-24 05:42:44
