
板栗林不同生境下八月竹叶片形态可塑性分析*
2022-07-09 13:49:42林上平柯宇鑫邓先俊杜旭华朱强根
世界竹藤通讯 2022年3期
林上平 柯宇鑫 邓先俊 杜旭华 朱强根*
(1 丽水学院生态学院 浙江丽水 323000;2 浙江省龙泉市林业局 浙江龙泉 323700;3 浙江农林大学林业与生物技术学院 浙江临安 311300;4 国家林业局竹子研究开发中心 浙江杭州 310012)
植物在生长发育过程中因受环境影响而产生一系列可塑性反应[1],如当光照、温度、水分和空气等生态因子变化时,植物会表现出一系列外部形态或内部生理上的变化,表现为植物对环境的适应性或表达响应[2]。作为植物的重要组成部分,叶片发挥着光合、蒸腾和气体交换等功能的主体作用[3],其形态特征(包括叶长、叶宽、叶周长、叶面积和比叶面积等) 对异质环境表现出强烈的敏感性和可塑性。因此,研究不同生境的叶片适应性在植物生态学领域具有重要意义。
叶片形态直接影响植物对光的接受能力、生物量的投资效率以及内外物质与能量的交换进程,而叶片形态的差异在很大程度上受环境的影响。例如,成向荣等[4]在研究披针叶茴香对变化光环境的响应中发现,披针叶茴香的平均叶长在低光照下比其他光照下的更大,且叶片面积随光照强度的减弱而增大。此外,水分也能影响植物叶片形态,叶片大小、叶片面积和叶生物量在土壤水分供应逐渐减少情况下表现出下降趋势[5]。植物可塑性是植物对其生长环境的适应性表现,往往朝着最优生长方向进化。目前,研究植物可塑性比较可靠的方法是异速生长分析[6],其主要优点在于能够量化参数间的相互关系,探究参数间的内在关系。本研究的目的是充分认识不同林冠下八月竹叶片形态的变化规律。
八月竹(Chimonobambusa szechuanensis) 为禾本科竹亚科寒竹属竹种,秆高2.5~4.0 (6.0) m,秆粗1.5~2.0 cm,是一种低矮的小径竹,原产于四川地区。本研究以套种于板栗林 (Castanea mollissima) 中的八月竹为研究对象,通过分析其叶片形态,了解其对不同生境的适应性规律,从而丰富引种栽培八月竹模式的理论储备,推动林下经济的高质量快速发展。
1 材料与方法
1.1 试验地概况
试验地位于浙江省西南部丽水龙泉市道太乡(北纬27°42′—28°20′,东经118°42′—119°25′),属于亚热带季风气候,年平均温度17.6 ℃,年降水量为1 699.4 mm,无霜期为263 d。板栗是当地的主要经济树种,其中大部分板栗林已经经营了20 年以上。引种套种八月竹的板栗林平均密度为45 株/667 m2,平均株高为5.0 m。
于2018 年选取经营22 年的板栗林0.4 hm2套种八月竹,套种栽植的八月竹有的在板栗林投影区内(林内),有的在投影区外(林窗),母竹种植平均密度为120 株/667 m2;另选取裸地0.2 hm2栽植八月竹,即纯林栽培。通过3 年的培育,各种植区八月竹的林地覆盖率均已达到80%。
1.2 试样采集与分析
2021 年9 月,在试验地板栗林不同生境下(林内、林窗、纯林)随机采集八月竹叶片,每类生境条件采取成熟完整叶片50 片,及时装入带水的密封袋中以防止叶片失水。将采集的不同生境下的八月竹叶片带回实验室,用扫描仪进行扫描分析,记录叶长(LL,cm)、叶宽(LW,cm)、叶周长(LP,cm)和叶面积(LA,cm2)等叶片形态特征数据。
1.3 数据处理
首先,运行SPSS 单样本K-S 检验叶片形态数据的正态分布,结果符合正态分布,进行下一步处理。在进行异速生长分析前,对数据进行对数(以10 为底) 转换。异速生长关系用方程y=bx^α来描述,线性转化为lny=lnb+αlnx,其中,b为二者关系的截距,α是二者关系的斜率,即异速生长指数,当α=1 时,即呈等速生长;当α大于或小于1时,即呈异速生长。采用标准主轴回归(SMA) 的方法在SMATR 软件中计算,当斜率存在差异时,进行多重比较;当斜率不存在差异时,则进一步采取Wald 显著性检验分析组间SMA 主轴是否存在沿y轴或斜率方向上的移动,并进行多重比较。
2 结果与分析
2.1 八月竹叶长与叶周长、叶面积的异速生长分析
2.1.1 叶长与叶周长
叶长和叶周长的关系采用SMA 分析,结果显示(图1),在不同生境条件下,二者的异速生长指数差异不显著(P=0.588),共同斜率为0.993(95%CI:0.976~1.011,P<0.01),与1.0 没有显著差异(P>0.05),即呈等速生长关系。进一步分析可知,与纯林相比,林窗和林内SMA 沿着共同斜率方向出现显著的正向移动,移动量分别为0.234 (P<0.01)、0.251 (P<0.01),表明叶长和叶周长整体上均表现为林窗和林内极显著大于纯林,平均叶长和平均叶周长的值分别为16.290、16.396、14.690 cm 和35.350、35.700、32.055 cm。
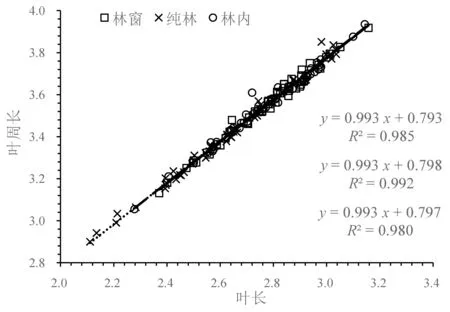
图1 不同生境下八月竹叶长与叶周长的异速生长关系Fig.1 Allometric relationship between leaf length and leaf circumference of Ch. szechuanensis in different habitats
2.1.2 叶长与叶面积
分析叶长与叶面积的关系显示(图2),在不同生境条件下,二者的异速生长指数没有显著差异(P=0.151),共同斜率为1.693 (95%CI:1.601~1.787,P<0.01),极显著大于1.0 (P<0.01),即呈异速生长关系。进一步从SMA 的截距分析得到,纯林和林内较林窗出现沿y轴的极显著正向移动,移动量分别为0.091 (P<0.01)、0.074 (P<0.01),而纯林、林内之间没有发生显著沿y轴移动(P>0.05),表明在调查的同等叶长范围内,在相同叶长情况时,纯林和林内比林窗有更大的叶面积。与纯林相比,林窗和林内SMA 沿着共同的斜率发生了显著的正向移动,移动量分别为0.313 (P<0.05)、0.409 (P<0.01),表明在整体上叶长和叶面积均表现为林窗显著大于纯林和林内极显著大于纯林,叶长和叶面积的平均 值 分 别 为16.290、16.396、14.690 cm 和20.573、22.284、19.521 cm2。

图2 不同生境下八月竹叶长与叶面积的异速生长分析Fig.2 Allometric relationship between leaf length and leaf area of Ch. szechuanensis in different habitats
2.2 八月竹叶宽与叶周长、叶面积的异速生长分析
2.2.1 叶宽与叶周长
分析显示(图3),在不同生境条件下,叶宽、叶周长的异速生长指数之间差异不显著(P=0.741),共同斜率为1.213 (95%CI:1.082~1.360,P<0.01),极显著大于1.0 (P<0.01),即呈异速生长关系。进一步分析得出,在共同斜率下,林窗、纯林和林内SMA 截距分别为2.864、2.706 和2.784,Wald 检验表明异速生长常数之间均存在极显著差异(P<0.01)。与纯林和林内相比,林窗SMA 的截距出现显著的沿y轴正向移动,移动量分别为0.158 (P<0.05)、0.080 (P<0.05);纯林较林内SMA 的截距存在显著的沿y轴正向移动,移动量为0.078 (P<0.05),表明在调查的同等叶宽范围内,在叶宽一致的情况下,叶周长表现为林窗>林内>纯林。

图3 不同生境下八月竹叶宽与叶周长的异速生长关系Fig.3 Allometric relationship between leaf width and leaf circumference of Ch. szechuanensis in different habitats
2.2.2 叶宽与叶面积
分析可知(图4),在不同生境条件下,叶宽、叶面积的异速生长指数之间没有显著差异(P=0.169),共同斜率为2.051 (95%CI:1.886~2.227,P<0.01),极显著大于1.0 (P<0.01),即呈异速生长关系。进一步分析,从SMA 的截距来看,林窗、林内较纯林存在沿着y轴的极显著正向移动,移动量分别为0.183 (P<0.01)、0.115 (P<0.01),林窗、林内之间则没有出现显著移动(P>0.05),表明在调查的同等叶宽范围内,当叶宽相同时,林窗、林内比纯林有更大的叶面积。与林内相比,林窗和纯林SMA 沿着共同斜率方向出现显著负向移动,移动量分别为0.023 (P<0.05)、0.274 (P<0.05),表明在整体上,不同生境下叶宽和叶面积均表现为林内>林窗和纯林。
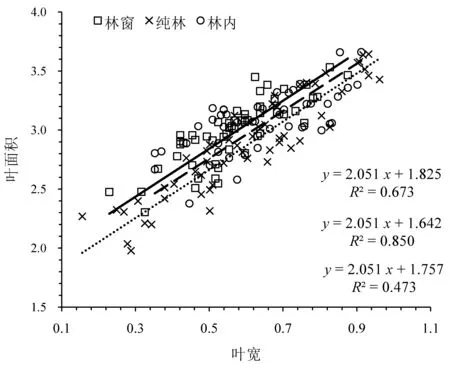
图4 不同生境下八月竹叶宽与叶面积的异速生长关系Fig.4 Allometric relationship between leaf width and leaf area of Ch. szechuanensis in different habitats
2.3 八月竹叶长与叶宽、叶周长与叶面积的异速生长分析
2.3.1 叶长与叶宽
分析显示(图5),在不同生境条件下叶长与叶宽的异速生长指数差异不显著(P=0.831),共同斜率为0.820 (95%CI:0.727~0.923,P<0.01),极显著小于1.0 (P<0.01),即呈异速生长关系。进一步分析得出,在共同斜率下,林窗、林内和纯林的截距分别为1.825、1.757 和1.642,Wald 检验表明异速生长常数之间均存在极显著差异(P<0.01)。与纯林和林内相比,林窗SMA 的截距出现沿y轴的极显著正向移动,移动量分别为0.183 (P<0.01)、0.068 (P<0.01);林内较纯林SMA 的截距存在沿y轴的极显著正向移动,移动量为0.115 (P<0.01),表明在调查的同等叶长范围内,相同叶长下的叶宽在不同生境下表现均为林窗>林内>纯林。
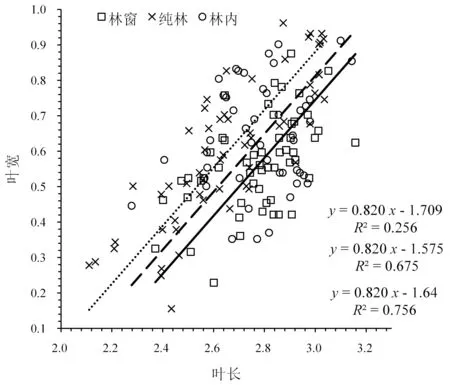
图5 不同生境下八月竹叶长与叶宽的异速生长关系Fig.5 Allometric relationship between leaf circumference and leaf width of Ch. szechuanensis in different habitats
2.3.2 叶周长与叶面积
分析显示(图6),不同生境条件下叶周长与叶面积的异速生长指数没有显著差异 (P=0.154),共同斜率为1.698 (95%CI:1.615~1.783,P<0.01),极显著大于1.0 (P<0.01),即呈异速生长关系。进一步分析,从SMA 的截距来看,纯林、林内较林窗出现沿y轴的极显著正向移动,移动量分别为0.086 (P<0.01、0.068(P<0.01),纯林、林内之间则没有出现显著移动(P>0.05),表明在调查的同等叶周长范围内,在相同叶周长下,纯林和林内比林窗有更大的叶面积。与纯林相比,林窗和林内SMA 出现沿着共同的斜率显著和极显著的正向移动,移动量分别为0.305 (P<0.05)、0.407 (P<0.01),林窗和林内之间没有显著移动(P>0.05),表明在整体上,不同生境下叶周长和叶面积均表现为林窗和林内>纯林。
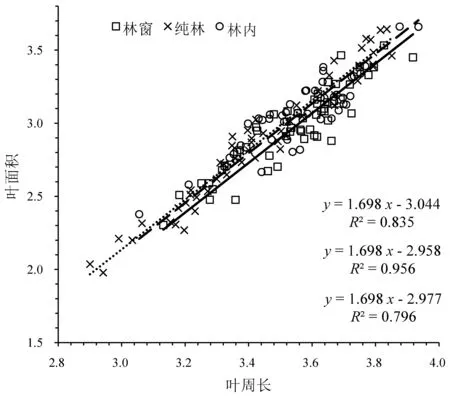
图6 不同生境下八月竹叶周长与叶面积的异速生长关系Fig.6 Allometric relationship between leaf width and leaf area of Ch. szechuanensis in different habitats
3 讨论与结论
植物通过表型[7-9]、生理结构[10-11]和物质与能量分配[12-13]等方面的改变,产生对异质生境的适应性,提高个体和种群的生存机会,属于植物的一种生活策略。本研究对板栗林不同生境下八月竹叶片形态特征进行异速生长分析,结果表明不同生境下叶片形态间关系的异速生长指数均无显著差异,即异速生长指数在不同生境下趋于稳定,这与不同生境下中华蚊母树叶片形态所表现的一样[14]。叶长与叶周长之间的异速生长指数与1.0 没有显著差异,即呈等速生长关系,表明叶长和叶周长的增长速度比较接近,而其他指标间关系的异速生长指数均与1.0 存在显著差异,即呈异速生长关系。
竹类属于克隆植物,具有很强的可塑性。大量研究表明,不同生境对竹类有显著影响,特别是在内部结构和外部形态上。筇竹竹秆结构在不同土壤水分条件下存在显著差异,在水分条件充足的情况下,通过增加维管束的长、宽来汲取更多的水分和养分;反之则产生大密度的小导管来适应相对缺水的环境[15]。高节竹的叶片和竹秆的形态受到海拔的显著影响,在中海拔的竹叶长、周长和面积均为最大,在海拔低的情况下则表现为最小[16]。林冠结构可以通过影响辐射分布和光诱导叶片生理性状的变化来影响森林生产力[17]。进一步Wald 显著性检验分析发现,不同生境组分之间SMA 均存在沿y轴或斜率方向上的显著移动。表明在调查同等的叶长范围内,在叶片宽度上,纯林比林窗和林内分别大1.143 cm 和1.067 cm。叶长和叶周长整体上均表现为林窗和林内极显著大于纯林,平均叶长和平均叶周长的值分别为16.290、16.396、14.690 cm 和35.350、35.700、32.055 cm。叶片大小整体上表现为林内>林窗>纯林。这与张萌等[18]研究的矢竹叶片对不同林冠的响应类似。多年经营的板栗林冠幅具有面积广、厚度大等特点,对生长在林内的八月竹起到遮荫效果,影响了林内的光照和辐射量的分布。植物在荫蔽环境下,由于光照条件较弱,往往会通过改变叶片形态、生物量分配以及植物组织结构等来提高光能利用率[19]。因此,林内的八月竹叶面积明显增大,以便接受更多的光照辐射来促进自身光合作用。在紫耳箭竹克隆形态可塑性对典型冠层结构及光环境的响应研究中,紫耳箭竹在低光环境下也是通过增加比叶面积、叶面积而提高分株对光能的利用能力[20]。
综上所述,板栗林不同生境条件下八月竹叶片形态特征间异速生长指数没有受到显著影响,但在叶片大小上发生了可塑性响应,总体上表现为林内>林窗>纯林。在光照辐射弱的林内,八月竹通过增大叶片面积,提高对光能的利用率,对不同生境产生可塑性。板栗林套种八月竹是一种新的林下经营模式,具有经济价值和生态效益,因此具有重要的研究价值。在今后的研究中,可以从叶片光合生理、呼吸作用以及水分代谢等生理生化层面深入研究板栗林不同生境条件下八月竹的可塑性,为农林生产提供丰富的理论指导。
猜你喜欢
西南林业大学学报(2022年5期)2022-10-17 09:56:38
农业机械学报(2020年1期)2020-03-13 00:27:38
新疆农业科学(2020年1期)2020-02-14 03:22:46
亚热带植物科学(2019年2期)2019-07-22 13:16:12
森林工程(2018年4期)2018-08-04 03:23:10
西部林业科学(2018年4期)2018-02-14 22:43:50
方圆(2018年23期)2018-01-07 09:06:18
江苏农业科学(2016年10期)2017-02-05 14:45:09
广西林业科学(2016年4期)2016-03-16 05:44:58
西南林业大学学报(2014年6期)2014-07-24 19:02:11
