
多年生黑麦草细胞分裂素信号通路B 类ARR转录因子LpARR10 的耐镉功能分析
2022-07-08 06:24:48张晴邢静姚佳明殷庭超黄心如何悦张敬徐彬
草业学报 2022年5期
张晴,邢静,姚佳明,殷庭超,黄心如,何悦,张敬,徐彬
(南京农业大学草业学院,江苏 南京 210095)
随着工业的不断进步,重金属资源的需求也日益增加,而重金属矿产资源的大量开采会导致矿区土地和水体遭受不同程度污染,其中重金属镉(Cd)是污染最为严重及生物毒性最强的元素之一[1]。近几年,随着《美丽乡村建设》和《山水林田湖草生态保护修复工程指南(试行)》政策的推行[2],重金属矿区及周边的土壤治理、植被恢复及园林绿化显得至关重要。草类是重金属修复的理想材料,不仅能吸收土壤重金属元素,还具有环境美化作用。因此,挖掘草类耐镉基因资源,并通过分子育种手段培育耐镉新品种具有重要的现实意义。
植物的耐逆反应受细胞分裂素、乙烯、脱落酸和茉莉酸等内源激素的调控[3]。细胞分裂素在植物耐逆反应中起到正调控作用,已有很多研究表明外源或内源增加(过量表达细胞分裂素合成基因ipt)能够有效提高植物的耐高温、干旱和高盐等能力[4-6]。近年来,有研究表明外源细胞分裂素处理也能够缓解重金属镉对植物造成的损伤[7-8]。例如,在镉胁迫下,外部喷施6-BA 提高了玉米(Zea mays)幼苗体内叶绿素和脯氨酸的含量,促进了叶片过氧化物酶(peroxidase,POD)和过氧化氢酶(catalase,CAT)活性的升高,并增加了玉米幼苗生物量[7]。另外,Gemrotová 等[8]的研究结果表明,在镉胁迫条件下,细胞分裂素拮抗剂PI-55 和细胞分裂素降解抑制剂INCYDE[2-chloro-6-(3-methoxyphenyl)aminopurine]处理对药用鳞芹属(Bulbine)植物幼苗的芽和根生长均具有显著影响,其中INCYDE 处理显著提高了其芽和根的生长。上述研究结果表明细胞分裂素有助于保护植物免受高Cd胁迫带来的损伤。然而,细胞分裂素调控植物耐镉的生理生化途径尚不清楚。细胞分裂素下游Arabidopsis Response Regulator(ARR)家族转录因子是调控细胞分裂素信号传导的关键节点[9]。根据核心功能域的相识度和 C 端功能域的长度,ARRs 可划分为两类,即 A 类和 B 类 ARRs[10-11]。B 类 ARRs 具有转录激活活性,通过促进细胞分裂素响应基因的转录,发挥正调控细胞分裂素信号传导的功能[12-13]。有研究证明,B 类ARRs 能够延缓拟南芥(Arabidopsis thaliana)的衰老,并提高其耐逆性[14-15]。因此,本研究推测细胞分裂素可能通过其信号传导通路B 类ARR 转录因子发挥调控耐镉的分子功能。
多年生黑麦草(Lolium perenne)是重要的早熟禾亚科冷季型草,具有牧草和草坪草两种生态型,在我国长江流域及以南的丘陵地区和人工草地中广泛种植[16]。多年生黑麦草的耐重金属镉的能力弱,限制了其在矿区土壤修复的利用。因此,培育耐镉黑麦草新品种具有重要意义。本试验以多年生黑麦草为研究材料,从已发表的多年生黑麦草转录组数据库中筛选出5个受逆境诱导的ARR 转录因子,其中LpARR10受镉诱导表达。因此,该研究拟通过分析LpARR10 的进化关系及功能域,探究LpARR10基因在镉逆境条件下的表达模式,评价LpARR10过量表达转基因株系的耐镉能力,以明确LpARR10基因的调控耐镉性的功能及途径。研究结果有助于进一步认识植物的耐镉机制,并为培育耐镉性强的黑麦草新品种提供基因资源。
1 材料与方法
试验于2018年9月-2020年7月在南京农业大学草业学院草类生理生化与分子生物学实验室进行。
1.1 LpARR10 功能域、进化与基因结构分析
从本研究团队已发表的多年生黑麦草全长转录组数据库(SRA 号:PRJNA335527)[17]中获取LpARR10编码区序列(coding sequence,CDS),采用NCBI-blastn 在线软件预测其蛋白功能域。从Tair 网站获取拟南芥A 类和B 类ARR 转录因子序列,利用MEGA 6.0 软件绘制LpARR10 和拟南芥A 类和B 类ARR 转录因子的遗传进化图。根据LpARR10的CDS 设计基因全长引物(表1),以多年生黑麦草(品种:Buena Vista)的gDNA 为模版,克隆并测定LpARR10的基因序列。根据其CDS 和基因组序列,利用BioXM 软件预测其外显子和内含子等基因结构。
1.2 LpARR10 基因的表达分析
多年生黑麦草种子播于陶粒土中,放置于智能人工气候室(夜间20 ℃,10 h;白天25 ℃,14 h;光强为750 mmol·m-2·s-1)。每 3 d 浇灌一次 1/2 霍格兰氏(Hoagland’s)营养液[18],待种子萌发 14 d 后挑取生长情况一致的幼苗水培于 Hoagland’s 营养液中,预培养 2 周后用 25 μmol·L-16-BA 和 200 μmol·L-1CdCl2处理其根部[19],并在0、0.5、2、6、12、24 和 48 h 后取根和叶片样品,冻存于-80 ℃冰箱,每个处理有 4个生物学重复。采用 Plant RNA Kit(Omega Bio-Tec,美国)提取样品的 mRNA,利用 Takara cDNA synthesis kit(含基因组 DNA 酶)将 mRNA 反转录为cDNA。采用罗氏荧光定量系统(Roche Light Cycler®480-Ⅱ)测定目的基因的相对表达量,目的基因及内参基因LpeLF4A的引物序列见表1。
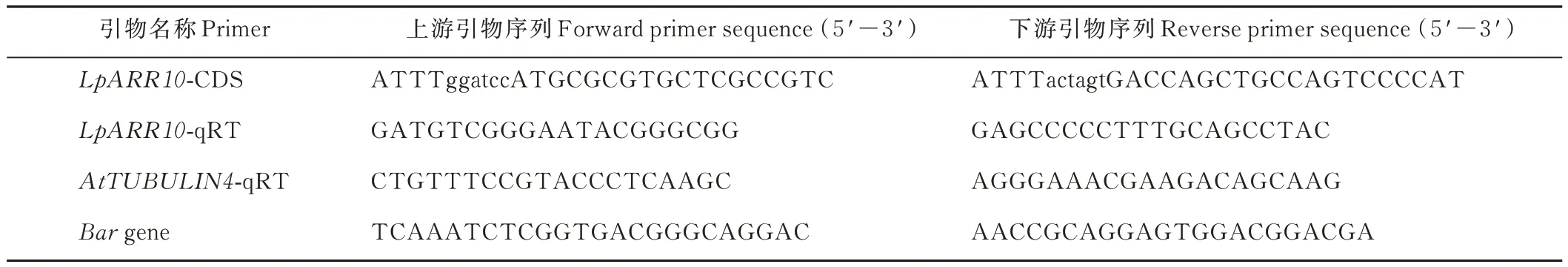
表1 试验所用引物Table 1 Primers used in the study
1.3 载体构建及拟南芥的遗传转化
以多年生黑麦草cDNA 为模版,采用NEB Q5 高保真酶克隆LpARR10编码区序列,经双酶切连接到入门载体 pEND-linker,利用 LR 反应(Gateway LR Enzyme Mix)将测序正确的 pEND-LpARR10与 pEarleygate-103 过量表达载体进行同源重组,获得35S∶∶LpARR10-GFP表达组件,将构建好的载体转入农杆菌中(AGL1菌株)。采用 LB 液体培养基(10 g·L-1胰蛋白胨,5 g·L-1酵母提取物,10 g·L-1氯化钠,pH=7.4)将含有过量表达载体的农杆菌培养至OD600约为1.0,离心后弃掉LB 培养基,用5%的蔗糖溶液(含有0.02% Silwet L-77)悬浮菌液,暗培养2 h 后侵染野生型拟南芥“Columbia”的花序[20]。一周后再次侵染花序,以提高转基因成功率。收取T0代种子,利用20%的84 消毒液浸泡消毒20 min,无菌水冲洗4 次,用0.2%的琼脂糖溶液将种子铺于含有20 μmol·L-1草铵膦(Basta)的 1/2 MS 培养基(主要成分为 650 mg·L-1NH4NO3,1.9 g·L-1KNO3,440 mg·L-1CaCl2·2H2O,370 mg·L-1MgSO4·7H2O,700 mg·L-1KH2PO4)上,培养一周后,筛选出抗性转基因植株。根据载体抗草铵膦基因(Bar)设计常规PCR 引物,以抗性转基因株系基因组DNA 为模版,利用2×PCR 预混液(Takara)进行PCR 扩增,检测抗性转基因株系基因组DNA 中是否有Bar基因,以筛选和鉴定阳性LpARR10过量表达转基因株系。获得稳定遗传的T3代阳性转基因株系,每个株系选取4 棵,分别提取叶片的总RNA 并进行反转录得到cDNA,用拟南芥TUBULIN4基因为内参基因进行定量PCR 反应,检测各转基因株系中LpARR10基因的表达。
1.4 转基因拟南芥的耐镉性分析
拟南芥野生型(wild type,WT)和T3代转基因株系种子用20%的84 消毒液浸泡消毒20 min,无菌水冲洗4次,用0.2%低熔点琼脂糖将种子铺于1/2 MS 和加有180 μmol·L-1CdCl2的1/2 MS 培养基中,4 ℃黑暗春化48 h,竖直放置于培养室中培养,10 d 后观察植株表型并统计根长和鲜重指标。本试验对转基因和野生型拟南芥进行Cd 处理的浓度(180 μmol·L-1)为预试验筛选得到。该试验独立重复3 次,单次试验每个株系至少种植4 皿用于数据的统计分析(4个生物学重复)。
1.5 数据统计与分析
使用SPSS 23.0 进行试验数据的计算、统计和差异显著性分析(P<0.05),数据以平均值±标准误差的形式呈现。利用SigmaPlot 12.5、MEGA 6.0 和Excel 2013 软件作图。
2 结果与分析
2.1 LpARR10 是 B 类 ARR 转 录因子
进化分析表明LpARR10 与拟南芥B 类ARR 转录因子聚为一组,且与AtARR10 和AtARR12 亲缘关系较近(图1),说明LpARR10 为B 类ARR 转录因子。LpARR10具有6个外显子,编码598个氨基酸(基因登录号为ALT32069.1,图2a)。LpARR10 的氨基酸序列与模式植物大麦(Hordeum vulgare)、拟南芥、水稻(Oryza sativa)和高粱(Sorghum bicolor)同源ARR 转录因子的相似度较高,且与大麦HvARR12 的同源性最高(图2b)。氨基酸序列分析表明LpARR10 具B 类ARR 转录因子特异的磷酸受体结构域(phosphoaccepter receiver domin of type B Arabidopsis response regulators,REC domain of B-type ARRs)、金属及磷酸化结合位点和YXXK 功能域(active site)(图 2b)。

图1 LpARR10 与拟南芥ARR 转录因子的系统进化树Fig.1 Phylogenetic tree of LpARR10 and Arabidopsis ARR transcription factors

图2 LpARR10 的基因组结构及其同源基因的氨基酸序列和功能域分析Fig.2 Genetic structure,amino acid sequence and function domain analysis of LpARR10 and its homologus genes
2.2 LpARR10 受细胞分裂素和Cd 诱导表达
LpARR10在多年生黑麦草根和叶片中均表达,因此,本研究分别测定了LpARR10在叶片和根中的表达变化。与处理前相比,细胞分裂素处理2 和6 h 后叶片中LpARR10的表达量显著升高,然而,在处理0.5、12、24 和48 h 后没有显著变化(图3a)。在根部,细胞分裂素处理均显著提高了LpARR10的表达量,其中在处理6 h 时表达量最高,是处理前的17.48 倍,说明根部对细胞分裂素处理更加敏感(图3b)。
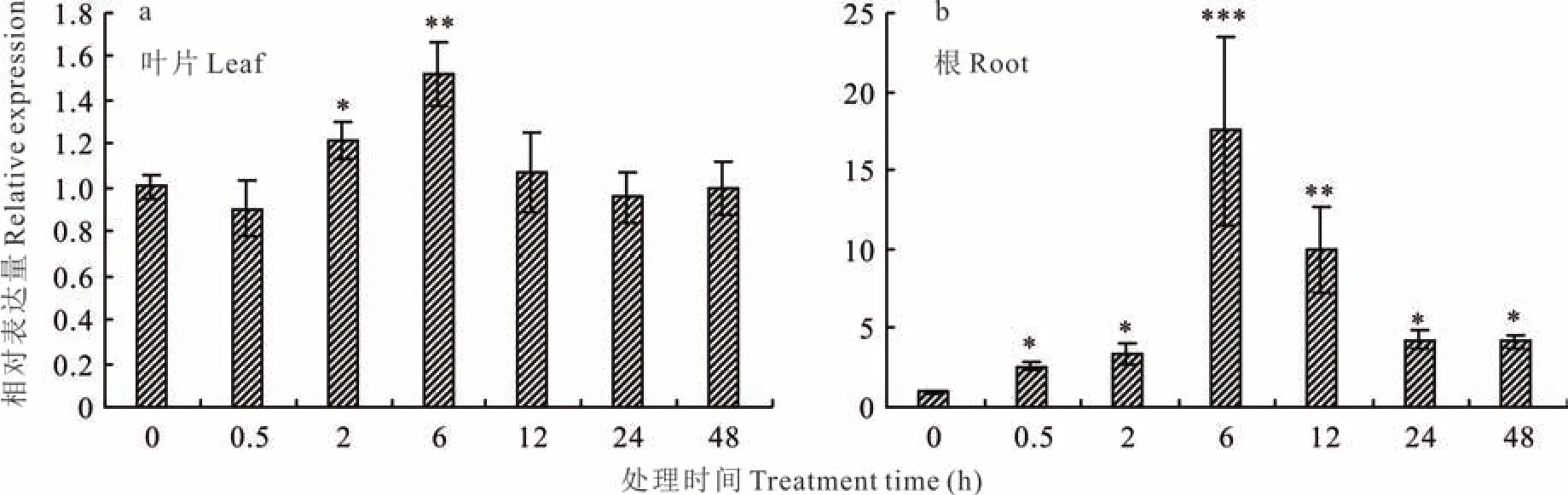
图3 外源细胞分裂素处理下多年生黑麦草叶片和根中LpARR10 基因表达分析Fig.3 Expression analysis of LpARR10 gene in perennial ryegrass leaves and roots treated with cytokinin
在叶片中,在镉处理2 和6 h 后,LpARR10基因的表达量显著升高,分别为处理前的1.41 和1.37 倍,随后表达量下降,而在 12、24 和 48 h 时与 0 h 无显著差异(图 4a)。在根中,Cd 处理 0.5 h 后即显著地提高了LpARR10基因的表达量,并在处理24 h 后,LpARR10的表达量达到峰值,其中12、24 和48 h 的基因表达量分别为处理0 h 时的3.85、9.53 和5.92 倍(图4b)。LpARR10的表达对镉和细胞分裂素的响应模式相似,在处理后期,表达量在根部增加的倍数明显高于叶部。
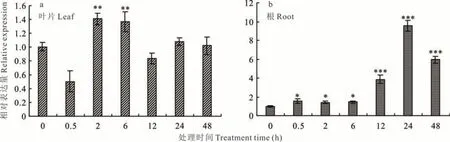
图4 镉处理下多年生黑麦草叶片和根中LpARR10 基因表达Fig.4 LpARR10 gene expression in leaves and roots of perennial ryegrass under cadmium treatment
2.3 LpARR10 过量表达转基因株系的鉴定
收取T0代种子,经草铵膦(Basta)抗性筛选,共得到3个抗性转基因株系(图5)。根据抗除草剂Bar基因设计特异性引物,进行PCR 检测,野生型拟南芥没有检测出目的基因条带,而转基因植株均检测出目标条带(图5a),且T3代转基因株系的种子均能在草铵膦筛选板上萌发(图5b),说明3个株系的T3代均为纯合转基因植株。进一步采用定量PCR 方法检测野生型和转基因株系中黑麦草LpARR10基因的表达量,结果表明LpARR10在3个转基因株系中均表达(图5c)。
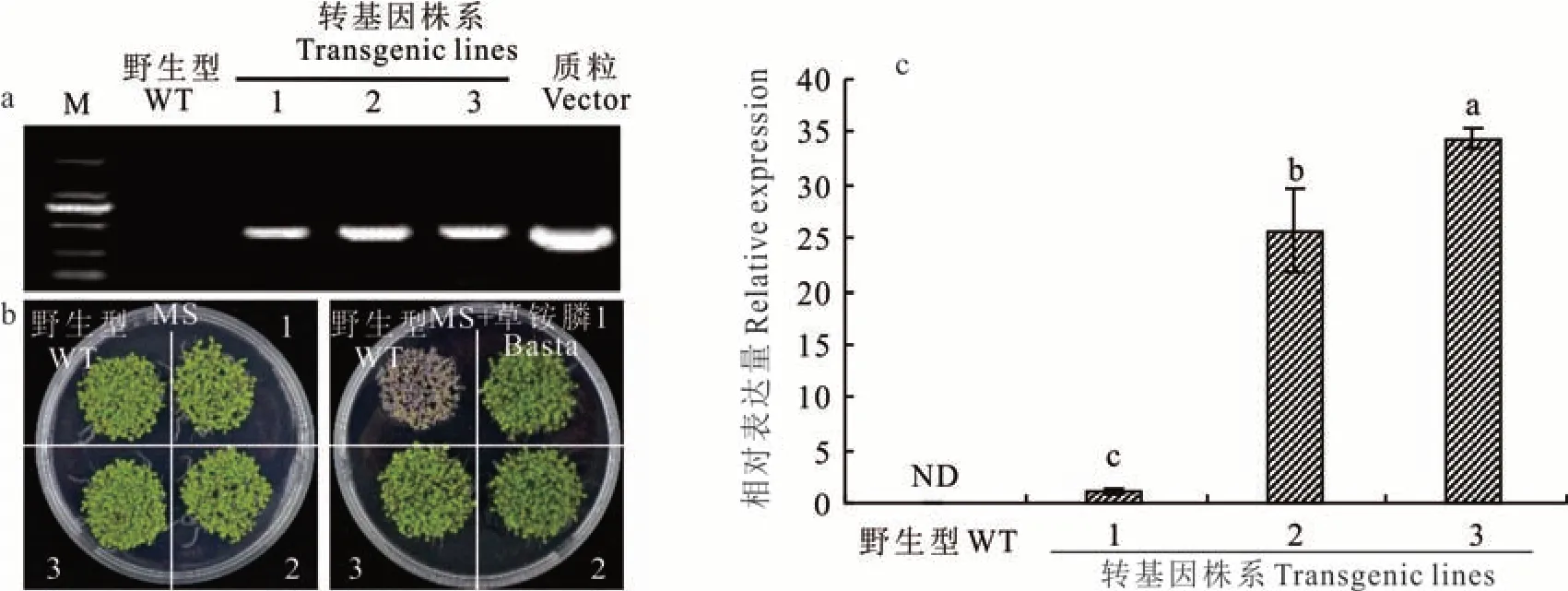
图5 LpARR10 转基因阳性株系的鉴定Fig.5 Identification of LpARR10 overexpression transgenic lines
2.4 过量表达LpARR10 提高了拟南芥耐镉能力
在无镉处理的条件下(1/2 MS 培养基),野生型与转基因株系的生长状况无明显差异(图6)。当处于180 μmol·L-1Cd 胁迫时,拟南芥植株叶片变黄,地上部和根系生长受到抑制。在野生型拟南芥中,Cd 处理使得根长和地上部鲜重分别降低了88.47%和72.82%。然而,在Cd 处理条件下,3个转基因株系的根长和地上部鲜重均显著高于野生型对照,说明LpARR10基因正调控拟南芥耐镉功能。尽管3个转基因株系中目标基因LpARR10的表达量有显著差异,但在正常和镉处理条件下,转基因株系之间的表型没有显著差异。
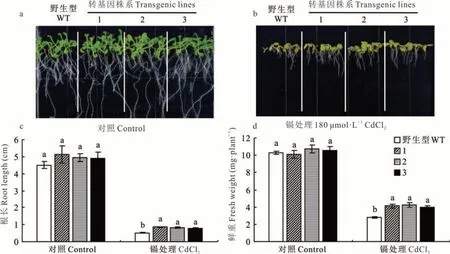
图6 镉处理条件下转基因株系和野生型拟南芥鲜重和根长Fig.6 Fresh weight and root length in transgenic lines and wild type Arabidopsis when treated with Cd
3 讨论
土壤是植物根部生长发育的环境,为植物提供必需的养分,也是很多微生物和小动物的栖息地。土壤重金属镉含量超标会降低土壤微生物和小动物群落丰度,限制植物的正常生长和发育,严重威胁生态系统安全[21]。植物可以从重金属污染土壤中吸收镉元素,从而降低其在污染土壤中的含量,是一种低成本、可持续、效率高的土壤修复方式[22]。草坪草物种多样性高,耐逆性强,可通过修剪地上部来降低土壤镉含量,是污染土壤植物修复较理想的选择。因此,挖掘草坪草耐镉基因、培育耐镉和富集镉的草坪草新品种具有重要意义。本研究以重要的冷季型草坪草-多年生黑麦草为材料,克隆出其B 类ARR 转录因子编码基因LpARR10,明确了LpARR10受镉胁迫诱导的表达模式,证明了LpARR10的耐镉功能,丰富了多年生黑麦草耐镉调控的分子途径。LpARR10可作为多年生禾本科草坪草分子育种重要的基因资源。
ARR基因是细胞分裂素信号传导关键基因,由于其首先在拟南芥中被克隆,因此命名为拟南芥响应调控因子[23-24]。高等植物ARRs 可分为A 和B 两类,是植物在生长、发育和抵御逆境过程中必不可少的一类转录因子。A 类ARRs 是细胞分裂素信号传导负调控因子,它可以通过竞争B 类ARRs 的磷酸转运蛋白(arabidopsis his phosphotransfer proteins,AHPs)和抑制B 类ARRs 的转录,发挥负反馈调控细胞分裂素信号传导的功能[9]。与A类ARRs 不同的是,B 类ARRs 含有较长的C 末端和典型的磷酸受体结构域(REC domain of B-type ARRs),具有转录激活活性,可以激活细胞分裂素响应基因的表达,发挥正调控细胞分裂素信号传导的功能[10-11]。有研究证明,拟南芥B 类ARR 转录因子ARR10能够参与细胞分裂素通路以调节拟南芥的生长和发育;ARR10和ARR12参与了细胞分裂素介导的拟南芥原生木质部分化的调控[25-26]。B 类ARR 转录因子也能参与生长素的信号传导,调控生长素相关基因的表达,进而影响植物的生长、发育[13]。B 类ARR 家族基因也能够调控植物逆境胁迫相关基因的表达,进而提高植物的耐逆性[14]。但是,B 类ARRs 在耐镉功能方面尚未见报道。本研究克隆得到了多年生黑麦草B 类ARR 转录因子LpARR10,将其与拟南芥所有ARRs 进行进化分析,结果显示LpARR10 与B 类ARRs 聚为一组,且与拟南芥AtARR10 和AtARR12 的亲缘关系较近。氨基酸序列分析表明LpARR10的C 端较长,且具有典型的B 类ARRs 磷酸受体结构域(REC domain of B-type ARRs)及金属和磷酸化结合位点,说明LpARR10属于B 类ARRs。本研究进一步在拟南芥中过量表达LpARR10,与野生型对照相比,转基因株系的耐镉能力显著提高,说明B 类ARRs(LpARR10)正调控植物的耐镉功能。本研究发现3个转基因株系中LpARR10上调表达的表达倍数不同,但它们的耐镉表型却没有明显差异,说明LpARR10可能存在翻译后修饰。前人研究表明磷酸转运蛋白(AHPs)能够磷酸化B 类ARRs,使其变成活化状态,进而发挥生物学功能[9-10]。本研究发现LpARR10 的第 79 位丝氨酸(serine,S)是 B 类 ARRs 成员保守的磷酸化位点,因此,本研究推测LpARR10的第 79位丝氨酸(S79)是介导AHPs 磷酸化LpARR10的关键位点。利用Crispr/Cas9 技术对多年生黑麦草基因组进行单碱基位点编辑已经实现[27],因此,后续利用Crispr/Cas9 技术对LpARR10第79 位丝氨酸位点进行突变,可以明确LpARR10翻译后修饰的途径及其发挥传递细胞分裂素信号功能的分子机制。
细胞分裂素是调控植物生长、发育、衰老和耐逆反应重要的植物激素之一。有研究表明外源喷施细胞分裂素和细胞分裂素降解抑制剂能够提高植物的耐镉能力[7-8],然而,其调控耐镉的分子途径尚未可知。本研究发现,LpARR10基因受细胞分裂素和镉处理上调表达,并且细胞分裂素和镉对于LpARR10表达的调控模式高度一致。例如,在叶中,LpARR10均在细胞分裂素和镉处理2 和6 h 时显著上调表达,其他处理时间无显著差异。在根部,LpARR10的表达在整个细胞分裂素和镉处理周期均显著高于0 h,上调表达倍数明显高于叶部。上述试验结果表明LpARR10可能是多年生黑麦草响应细胞分裂素和镉处理的关键节点,是细胞分裂素调控耐镉反应的关键基因。异源过量表达试验也证明,LpARR10的确具有正调控拟南芥的耐镉功能。综上所述,本研究初步明确了细胞分裂素通过其信号传导通路基因LpARR10发挥正调控植物耐镉的功能。
4 结论
本研究通过克隆多年生黑麦草细胞分裂素信号传导通路LpARR10基因,并根据进化和氨基酸序列分析,发现LpARR10 与拟南芥B 类ARR 转录因子聚为一组,且具有高等植物B 类ARRs 高度保守的磷酸受体结构域(REC domain of B-type ARRs)及金属和磷酸化结合位点,说明LpARR10 为B 类ARRs。定量表达分析表明,无论在叶部还是在根部,LpARR10均受细胞分裂素和镉处理诱导上调表达,但在根部上调表达的倍数明显高于叶部。另外,LpARR10在细胞分裂素和镉处理条件下表达模式较为一致,说明其可能是细胞分裂素调控植物耐镉功能的关键节点。异源过量表达LpARR10显著提高了拟南芥的耐镉能力,表明细胞分裂素极可能通过LpARR10调控耐镉的功能。该研究结果对深入揭示多年生黑麦草B 类ARR 转录因子的功能,进一步培育耐镉多年生黑麦草的新品种具有重要的理论意义。
猜你喜欢
奥秘(创新大赛)(2022年8期)2022-09-14 05:44:42
亚热带植物科学(2022年1期)2022-05-17 12:39:32
中国果业信息(2021年10期)2021-12-07 04:06:06
湖南农业大学学报(自然科学版)(2021年3期)2021-07-02 02:17:38
养殖与饲料(2020年6期)2020-02-18 16:01:26
中国种业(2019年8期)2019-08-26 08:41:18
四川农业科技(2019年5期)2019-07-01 09:46:46
浙江柑橘(2016年1期)2016-03-11 20:12:31
中学生物学(2016年8期)2016-01-18 09:08:20
中学生物学(2013年6期)2013-04-29 21:24:30
