
翘嘴鲌生长激素受体1 基因内含子1、2 中微卫星多态性及与生长性状的关联分析
2022-07-01 02:51刘士力李飞贾永义郑建波迟美丽程顺卞玉玲顾志敏
水产学杂志 2022年3期
刘士力 ,李飞 ,贾永义 ,郑建波 ,迟美丽 ,程顺 ,卞玉玲,顾志敏
(1.浙江省淡水水产研究所,农业农村部淡水渔业健康养殖重点实验室/浙江省淡水水产遗传育种重点实验室,浙江 湖州 313001;2.上海海洋大学农业农村部淡水水产种质资源重点实验室,上海 201306)
鲌亚科中体型最大的翘嘴鲌(Culter alburnus Basilewsky)(俗称白鱼)肉质洁白、细嫩而不腥,是我国重要的淡水名优经济鱼类之一。翘嘴鲌是以活鱼为饵料的凶猛肉食性鱼类,能起到维持淡水水域生态系统稳定性的作用,具有良好的生态价值。自20世纪90 年代人工繁殖获得成功后,其养殖技术、饲料开发、产品加工等均得到快速发展[1],已经成为了稳定的养殖鱼类。开展翘嘴鲌的育种具有重要的经济意义,浙江省淡水水产研究所已开展了翘嘴鲌的群体选育、杂交育种、雌核发育及伪雄鱼诱导工作,培育了2 个水产新品种。
最早在1976 年,Tsushima 等就在兔的肝细胞膜上发现了生长激素受体(Growth hormone receptor,ghr)基因[2]。生长激素(Growth hormone,GH)必须通过与靶组织细胞表面的GHR 结合而启动细胞内的信号传导机制,发挥其生理功能。目前在哺乳动物中仅发现一种编码GHR 的基因[3],但在很多鱼类基因组中却发现了另外一种基因编码的生长激素受体GHR2[4-6]。鉴于GHR 的重要功能,其DNA 序列的多态性可能与生长性状相关。倪静等[7]分析了两个群体牙鲆(Paralichthys olivaceu)ghr基因5′ 端启动子区的微卫星,发现均与生长具有一定的相关性。目前已开发了一些翘嘴鲌的微卫星标记[8,9],开展了翘嘴鲌GH-IGFs 轴相关基因及微卫星的研究[10-12]。刘士力等[13]以翘嘴鲌转录组中获得的mRNA 为基础,获得了ghr1 和ghr2 的部分基因序列。经过初步研究,发现其中4 个多态性微卫星位点与生长性状具有一定相关性。ghr1 中内含子1、2序列较长,当时没有对其内含子1、2 中微卫星进行实验。本试验进一步研究其中8 个微卫星位点的遗传多样性,分析了微卫星基因型和翘嘴鲌的体长、体质量的相关性,可为翘嘴鲌ghr1 基因微卫星特征研究及分子标记辅助育种提供基础数据。
1 材料与方法
1.1 材料
实验用翘嘴鲌雌鱼40 尾、雄鱼68 尾采自浙江省淡水水产研究所八里店综合实验基地,人工催产后放入产卵池产卵,收集受精卵孵化后养殖在同一条件下。12 月龄时剪取少量尾鳍,用无水乙醇浸泡于-20℃保存备用。
用于翘嘴鲌DNA 提取的试剂购自生工生物工程(上海)股份有限公司;PCR 反应试剂购自宝日医生物技术(北京)有限公司。
1.2 方法
1.2.1 DNA 提取
采用苯酚-氯仿法提取样本DNA。用1%琼脂糖凝胶检测提取DNA 完整性。DNA 原液于-20℃保存备用。
1.2.2 引物设计
根据GenBank 公布的翘嘴鲌ghr1 基因及其侧翼DNA 序列(登录号:KX925976),在内含子1 中有3 个微卫星位点,内含子2 中有5 个微卫星位点。应用Primer Premier 6.0 软件分别设计8 对引物用于微卫星位点的扩增,上游引物的5′ 端加上Tail A 碱基序列,通用引物Tail A 5′端用FAM进行修饰。具体引物由生工生物工程(上海)股份有限公司合成(表1)。

表1 翘嘴鲌ghr1 基因内含子1、2 中微卫星引物序列Tab.1 Primer sequences used for microsatellite amplification in the intron 1 and 2 of the ghr1 gene in topmouth culter C.alburnus
1.2.3 微卫星扩增及分型
微卫星扩增的PCR 反应体系和程序参照本实验室之前的操作[12]。PCR 产物用1%琼脂糖电泳检测后送生工生物工程(上海)股份有限公司进行微卫星分型。
1.2.4 数据分析
采用POPGENE 1.3.1 计算微卫星座位在该群体内的观测等位基因数(Observed number of alleles,Na)、观察杂合度(Observed heterozygosity,Ho)、期望杂合度(Expected heterozygosity,He)、有效等位基因数(effective number of alleles,Ne)和多态信息含量(Polymorphism information content,PIC)。运用SPSS 13.0 的一般线性模型(General Linear Model,GLM)程序分析微卫星位点与翘嘴鲌体质量和体长的关系。显著性分析由单因素方差分析(ANOVA)方差分析的Duncan 氏法进行多重比较,P<0.05 为差异显著。结果以平均值±标准误(Mean±SE)表示。
2 结果与分析
2.1 翘嘴鲌ghr1 基因内含子1、2 中微卫星位点检测结果
琼脂糖电泳结果表明:所设计引物扩增的产物条带清晰,无杂带,长度符合预期。将这8 个微卫星位点在24 个样本中进行多态性检测,结果表明除了ghr1-2 中(CA)6微卫星位点外,ghr1 中的另外7个位点均具有多态性。将这7 个微卫星位点在120尾同批繁殖、同塘养殖的翘嘴鲌中进行分析。
位于第1 内含子中的位点ghr1-1 和ghr1-3 分别具有16 和9 个等位基因,ghr1-1 中的优势等位基因为451 bp、453 bp 和455 bp,3 个总频率占54.7%。ghr1-3 中的优势等位基因261 bp 频率为43.0%,微卫星重复次数为9 次。重复次数为7、8、9、10 次的基因型占全部基因型的93.9%。
位于第2 内含子中的位点ghr1-4 具有非常高的多态性。检测到34 个等位基因,具有76 种基因型。由于基因型较多,没有频率超过5%的基因型。在ghr1-5 和ghr1-6 均观察到3 个位点,其中ghr1-5 检测到3 个等位基因,分别为277 bp、279 bp和281 bp,对应的微卫星核心重复次数分别是3、4和5 次,其中等位基因281 bp 的频率最高,达到了96.2%;ghr1-6 检测到3 个等位基因,分别为375 bp、377 bp 和379 bp,对应的微卫星核心重复次数分别是7、8 和9 次,其中等位基因377 bp 的频率最高,达到了91.2%。ghr1-7 为本次研究唯一的四碱基重复微卫星,检测到17 个等位基因,具有62 种基因型。ghr1-8 有7 个等位基因,其中等位基因456 bp 频率为49.2%,占有绝对优势。
2.2 翘嘴鲌ghr1 基因内含子1、2 中微卫星位点的遗传多态性参数
遗传特性分析结果表明,翘嘴鲌ghr1 基因内含子1 和2 中的这7 对引物观测等位基因数(Na)为3~34(平均12.714),期望杂合度(He)为0.074~0.929(平均0.625),香农指数(I)为0.1882~2.9185(平均1.617),多态信息含量(PIC)为0.073~0.898(平均0.608),有效等位基因数(Ne)为0.074~0.898(平均5.839),其中ghr1-4、ghr1-7 的Ne 均大于10,具有良好的多态性。ghr1-5 和ghr1-6 的Ne 小于2,PIC小于0.25,属低度多态位点。其余5 个微卫星位点PIC 均大于0.5,属于高度多态位点。
2.3 翘嘴鲌ghr1 基因内含子1、2 中微卫星位点与体长和体质量的关联分析

表2 翘嘴鲌ghr1 基因内含子1、2 中微卫星位点的等位基因频率Tab.2 The allele frequency of two microsatellite loci in the intron 1 and 2 of gene in topmouth culter C.alburnus ghr1
除去基因型个体数目小于5%的数据后进行单因素方差分析。ghr1-4 位点由于有76 种基因型,但各基因型的个体数目不够,因此被排除在分析之外。ghr1-5 位点281/281 型占92.5%,另外两种基因型较少,也被排除在分析之外。因此共有5 个位点参与关联分析。
微卫星位点ghr1-3 对于体长的关联分析结果表明:257/259 型个体占样本数的7.5%,其体长最长,也是体质量最高的个体;259/261、259/263 和261/263 型的体长较短,显著低于257/259 型个体。对于体质量的分析结果表明,257/259 型个体除了显著高于这3 种基因型外,还高于257/261 和261/261 型(P<0.05)。其余位点不同基因型个体体长和体质量均有差异,但差异均不显著(P>0.05)。
3 讨论
微卫星DNA(Microsatellite DNA)是真核生物基因组重复序列中的重要组成部分,稳定性好、可重复性强、检测技术高效简便,但筛选多态性微卫星是一个相对繁琐的过程。以前常用的方法有磁珠富集法、跨物种扩增和转录组数据筛选等。但这些方法都有其局限性,如磁珠富集法和所采用的探针有很大的关系。目前采用较多的是CA 和GATA 探针,这两种探针的微卫星在基因组中含量更高一些,但忽略了探针以外其他微卫星的研究。跨物种扩增由于进化和变异的关系,扩增的产物不一定包含微卫星,扩增产物长度的变化可能不是微卫星造成,这会给实验结果带来误差。转录组中获得的微卫星往往多态性不高,限制了其应用范围。功能基因中的微卫星更有可能与生长性状相关,但某些内含子较长,不易扩增。通过基因组直接筛选是最为准确的方法,近年来高通量测序技术不断完善,测序的通量增大,读长增加,成本大幅下降。本实验通过翘嘴鲌基因组序列分析获得了ghr1 的内含子1,2 的序列,研究了其中的微卫星多态性。

表3 7 个微卫星位点在120 尾翘嘴鲌中的多态性信息Tab.3 Polymorphism information for 7 microsatellite loci in 120 individuals of topmouth culter C.alburnus
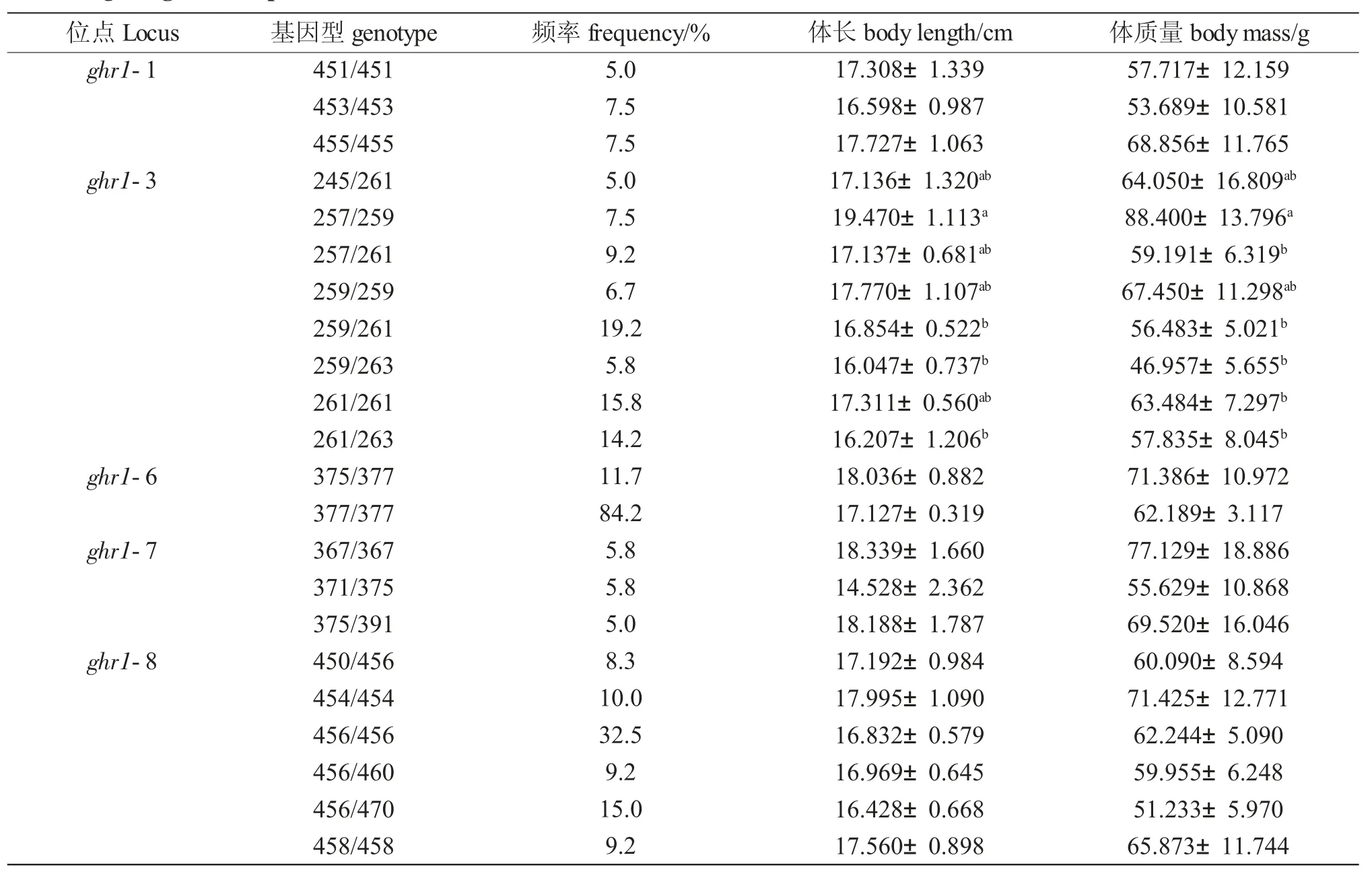
表4 翘嘴鲌ghr1 基因内含子1、2 中微卫星位点不同基因型与生长性状的关联分析Tab.4 Association analysis between growth traits and different genotypes of microsatellite locus in the intron 1 and 2 of ghr1 gene in topmouth culter C.alburnus
本试验检测了翘嘴鲌群体中ghr1 基因内含子1、2 中的8 个微卫星位点,表明7 个具有多态性。ghr1-6 和ghr1-7 属于低度多态性位点(PIC<0.25),另外5 个属于高度多态性位点(PIC>0.5)。ghr1-4 和ghr1-7 有效等位基因数高于10,在个体识别中具有良好的应用前景。ghr1-7 为四碱基重复微卫星(GATA)12,在120 个样品中检测到17 个等位基因,具有62 种基因型,多态信息含量为0.898,具有良好的多态性。其余为二碱基重复微卫星,5 个为CA重复类型,1 个为GA 重复类型,1 个为CA 和GA的复合型。在这7 个二碱基重复微卫星位点中,ghr1-2 不具有多态性,ghr1-5 和ghr1-6 虽然分别具有3 个等位基因,但其中优势等位基因均在90%以上,有效等位基因数低于1.2,PIC 小于0.25,属于低度多态性位点。对这些微卫星重复次数的观察,发现ghr1-5 位点分别为277 bp、279 bp 和281 bp,对应的微卫星核心重复次数分别是3、4 和5 次。其中等位基因281 bp 的频率最高,达到了96.2%;ghr1-6 检测到3 个等位基因,分别为375 bp、377 bp和379 bp,对应的微卫星核心重复次数分别是7、8和9 次,重复次数均为连续。ghr1-4 为复合型微卫星,检测到34 个等位基因,具有非常高的多态性,没有观察到明显的规律。其他微卫星位点中占10%以上的位点区域都是连续的。如ghr1-8 中,456 占49.2%,其相邻的454 和458 均为11.3%,3 者合计占71.8%。推测这些微卫星核心重复次数在变异过程中倾向于1 次增加或减少了1 个重复。
研究人员经常在ghr1 基因中获得一些和生长性状关联的分子标记。陶文静等[14]对5 尾鲤(Cyprinus carpio)的4 个ghr 基因进行检测,共找到38 处SNPs 位点,采用PCR-RFLP 方法大规模检测了其中5 个SNP 位点,发现均与增重显著相关。半滑舌鳎(Cynoglossus semilaevis)ghr1 外显子8 和9 中各存在1 个SNP 位点。在外显子8 的SNP 位点中不同基因型的样本的在ghr1 的表达和生长性状均有显著区别[15]。除了SNP 位点外,ghr1 基因序列中的SSR 也有可能与生长性状关联。倪静等在牙鲆ghr基因5′端启动子区发现一个多态性微卫星位点,在2 个养殖群体的不同基因型样本之间生长性状均有显著差异[7]。本团队已初步研究了翘嘴鲌ghr1 中的微卫星,外显子2 和内含子8 中分别包含(CT)6和(AC)5的微卫星序列。但这2 个微卫星位点在所研究的群体中均没有多态性。内含子1,2 相对较长,当时没有对其中的微卫星进行研究。在获得翘嘴鲌基因组序列后,对其中包含微卫星的研究变得简便。通过120 尾同批繁殖、同塘养殖的翘嘴鲌中进行实验。微卫星位点的关联分析表明,ghr1-3 位点中高于或等于5%的基因型有8 个。257/259 型个体占样本数的7.5%,其体长和体质量均高于其他基因型,其体质量与245/261 和259/259 差异不显著,但显著高于另外5 种基因型(P<0.05)。接下来将在不同地理种群或者近缘物种做进一步研究。本研究结果从分子水平上为翘嘴鲌的保种、选育和高效育种提供了理论依据。
猜你喜欢
作物学报(2022年12期)2022-10-14
中国典型病例大全(2022年9期)2022-04-19
安徽农学通报(2022年6期)2022-04-07
新课程·下旬(2018年9期)2018-11-14
新课程·下旬(2018年8期)2018-11-10
中国医药导报(2018年14期)2018-08-30
医学信息(2017年1期)2017-02-28
中学生理科应试(2016年4期)2016-11-19
青少年科技博览(中学版)(2015年10期)2015-01-11
中国当代医药(2009年22期)2009-05-14
