
稻曲病菌生物学特性研究
2022-06-28 01:17胡建坤李保同刘永锋潘夏艳曹慧娟黄瑞荣
江西农业学报 2022年4期
黄 蓉,胡建坤*,李保同,刘永锋,潘夏艳,曹慧娟,黄瑞荣**
(1.江西省农业科学院 植物保护研究所,江西 南昌 330200;2.江西农业大学 国土资源与环境学院,江西 南昌 330045;3.江苏省农业科学院 植物保护研究所,江苏 南京 210014)
0 引言
稻曲病是由稻曲病菌Ustilaginoidea virens(Cooke)Tak引起的一种水稻穗部谷粒病害,该病广泛分布于亚洲、非洲、南美洲、欧洲各地的水稻主产区,其中,在中国、印度、菲律宾和日本等亚洲国家发生较重[1]。该病害在历史上属零星发生的次要病害,但随着灌溉与施肥水平的提高以及高产型品种的普及推广,已经发展成水稻的主要病害之一[2-3]。为了有效地控制稻曲病的发生与为害,保障国家粮食生产安全,人们针对该病害的发生规律与防治技术进行了研究,并多有报道[4-8]。认识病原菌的生物学特性是研究病害灾变规律的前提。国内有关稻曲病菌生物学特性的研究已取得一定进展,王疏等[9]报道了稻曲病菌厚垣孢子的萌发条件;张君成等[10]对稻曲病菌厚垣孢子和分生孢子的萌发特征进行了研究,认为基质养分对分生孢子的萌发有影响,并指出稻曲病菌生长缓慢,在人工培养基上需20~30 d产生厚垣孢子;李阳等[11]报道了在不同培养基中稻曲病菌孢子的萌发及病菌的生长特性;杨秀娟等[12]报道了不同培养基、温度、pH值对稻曲病菌生长及其分生孢子萌发的影响。然而,在自然条件下田间稻曲球上厚垣孢子的萌发情况还少见报道。南方稻田中稻曲球产生菌核的情况也有报道[5,13-14],但是否广泛存在尚未形成共识。稻曲病菌无性孢子在病害侵染循环中起到了重要作用,研究稻曲病菌不同颜色的孢子产生与萌发条件,有助于了解该病害的致病机制和传染途径[9]。鉴于目前人们对稻曲病菌生物学特性并不完全清楚,对病菌初侵染来源也存在不同见解,本文通过研究江西稻曲病菌的厚垣孢子在水中的萌发、菌丝在不同培养基中的生长与产孢、菌核形成条件及厚垣孢子的致病性,以期增进人们对稻曲病菌生物学特性的认识,能够从多视角深入了解病原菌的侵染机制。
1 材料与方法
1.1 试验材料
1.1.1 供试菌株 新鲜稻曲球采自江西省农科院水稻生产试验基地,采用组织分离法对病原菌进行分离,经纯化后转入PSA试管斜面中保存备用。
1.1.2 供试培养基 马铃薯蔗糖培养基(PSA):马铃薯200 g、蔗糖20 g、琼脂17 g、H2O 1 L;胁本哲氏培养基:马铃薯300 g、蔗糖15 g、蛋白胨5 g、Ca(NO3)2·4H2O 0.5 g、Na2HPO4·12H2O 2.0 g、琼脂17 g、H2O 1 L;大米粒培养基:大米加水煮熟后分装灭菌。
1.1.3 供试水稻品种 两优培九。
1.1.4 供接种用稻曲球 采自江西宜黄县、南昌县的稻田病株。
1.2 试验方法
1.2.1 不同成熟度稻曲球中厚垣孢子萌发特性观察 在前期研究[15]的基础上,2016年对江西稻曲病菌进行了系统的研究。在田间采集黄色或黑色稻曲球的病稻穗装入封口塑料袋中带回实验室,用小金属镊摘取黄色和黑色稻曲球各1~2粒,分别放入盛有15 mL清水和2%蔗糖溶液的培养皿中,用镊子夹住稻曲球,将球体上的厚垣孢子洗入培养皿水溶液和蔗糖溶液中,并用玻棒不断搅动使洗下的孢子全部悬浮在液体中。厚垣孢子在室温下孵育0、2、4、6、12、24、36和48 h,每个时间点从培养皿中分别用玻棒蘸取黄色和黑色厚垣孢子悬液置于载玻片中制作观察玻片2片在10×10视野下进行镜检,观察孢子的萌发状况。每片随机查看10个视野里的孢子数,统计萌发的孢子数,记载孢子总数累计2000个。0 h为从稻曲球中直接取孢子制片镜检,观测的视野与孢子数不限。计算厚垣孢子萌发率,并采用DPS软件中Duncan’s新复极差法进行方差分析。孢子萌发率计算方法如下:
厚垣孢子萌发率(%)=(萌发孢子数/观察孢子总数)× 100%
对冷藏保存的厚垣孢子活性进行测定。从冰箱中取出保存1年的稻曲球,用清水洗下球体上的厚垣孢子并在水中孵育12 h,取孢子悬液制作观察玻片,在光学显微镜下观察孢子萌发情况。
1.2.2 不同固体培养基对稻曲病菌菌丝生长的影响 在无菌条件下用打孔器将PSA培养基上培养好的稻曲病菌打取直径为5 mm的稻曲病菌菌饼,分别接种到马铃薯蔗糖培养基、胁本哲氏培养基、大米粒培养基3种固体培养基上,置于26 ℃恒温光照条件下培养,每处理接5皿,25 d后测量菌落直径并计算菌丝日平均生长率。
1.2.3 田间菌核调查 于不同年份的水稻黄熟期、收获期,在江西省内多县(市)开展早稻、一季稻和二晚稻的稻曲病穗上随机调查稻曲球着生菌核情况,并对采收晾晒的稻谷进行查看是否有菌核存在。如田间查找到菌核存在,则调查一定数量的稻曲球测算生长菌核球体所占比率。
1.2.4 稻曲病菌厚垣孢子致病性试验 2016年8月底采集的稻曲球在冰箱中冷藏数天后,取出并用清水洗下其上的厚垣孢子,并在室内常温下水孵育4 h,于水稻破口前5 d采用医用注射器将菌悬液注入稻苞内,孢子悬液含孢子数浓度≥1×106个/mL,接种菌液量为菌液溢出时停止;喷雾接种于9月中旬进行,分2次接种,第一次为破口前5 d,第二次为破口期。2017年接种体来源于上年8月底从稻曲病田采集并在冰箱冷藏保存的稻曲球上的厚垣孢子,于次年9月水稻孕穗破口前期至破口期从冰箱中取出,用清水洗下厚垣孢子并在水中孵育4 h;接种菌悬液量为30 L/667 m2。2种接种法均于下午4 h后在温网室的水泥池中进行,温网室配有遮阳网和微喷设施,接种后采用微喷定时喷洒清水保湿。试验以注射或喷雾清水为对照。
2 结果与分析
2.1 不同色泽稻曲球中厚垣孢子的萌发特性
试验观测到从田中取回的黄色与黑色稻曲球,有少数球体上的厚垣孢子少量产生芽管进入萌发状态(图1A、图1B),这一现象在不同年份取样检测都有所发现,但黑色球体上厚垣孢子产生芽管的现象并不多见。黄色稻曲球上的厚垣孢子在清水中孵育2 h即有少量萌发,产生芽管,其顶端产生1个分生孢子(图1C),4~6 h在第1个分生孢子旁又产生1~2个分生孢子(图1D)。
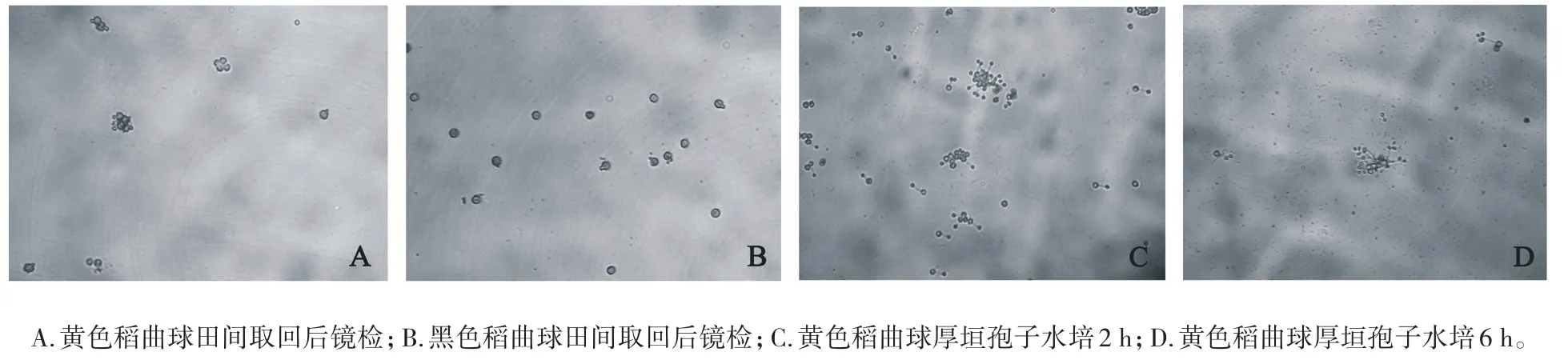
图1 稻曲病菌厚垣孢子在不同时间点的萌发情况
在清水中和2%蔗糖溶液中黄色和黑色稻曲球上的厚垣孢子萌发率存在显著差异,一是清水中的萌发率和萌发速率均比蔗糖溶液中的要高和快;二是黄色稻曲球上的厚垣孢子无论是在清水中还是在蔗糖溶液中,均比黑色稻曲球上厚垣孢子萌发率高,萌发速率快(表1)。黄色球体厚垣孢子的萌发率在清水中12 h之后基本不再增加,稳定在87%左右;在蔗糖溶液中24 h之后萌发率接近清水中的萌发率,两者无显著性差异。黑色球中的厚垣孢子的萌发率和萌发速率虽低于黄色球体的厚垣孢子的萌发率,但其自身在清水和蔗糖溶液中的萌发率在24 h之后也趋于一致,无显著性差异。镜检观测保存在冰箱中1年的稻曲球上的厚垣孢子的萌发情况,厚垣孢子经清水孵育24~48 h后观察到一定数量的萌发孢子,其萌发率可达10%左右。
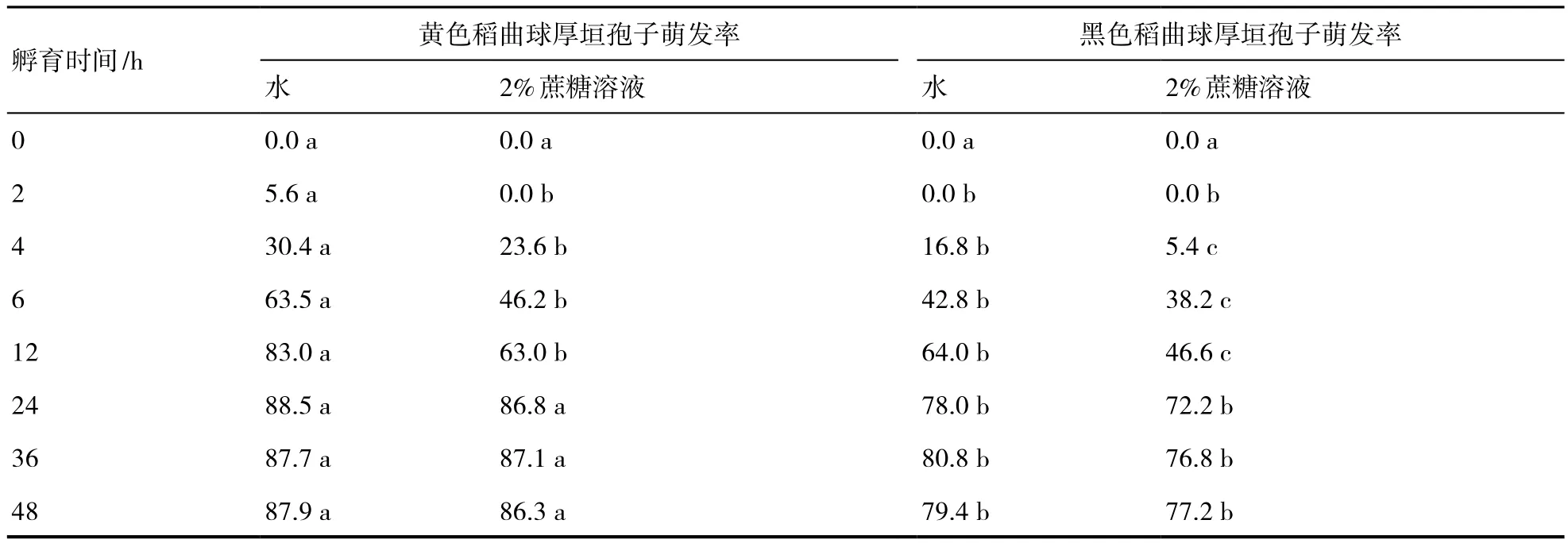
表1 黄色和黑色稻曲球中的厚垣孢子在不同基质中萌发率比较 %
2.2 分生孢子的萌发特性
分生孢子在清水中2~4 h从一端或两端萌发。厚垣孢子起始萌发一般从萌发的芽管中产生1个分生孢子,置于芽管顶端,4~6 h后顶部可见1~5个次生小分生孢子(图2A、图2B),6~8 h后分生孢子再次萌发产生枝状次生分生孢子梗,其顶端产生次生小分生孢子,12 h后二、三级次生分生孢子的分生孢子比达60%以上。孢子成熟脱落,镜检中可观察到大量的小孢子出现。分生孢子无色,卵圆形或椭圆形,次生分生孢子比老熟的分生孢子小。
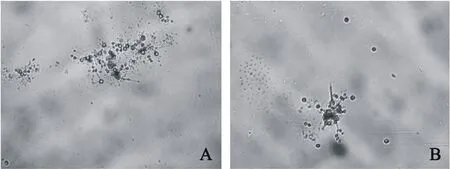
图2 稻曲病菌分生孢子萌发产生次生分生孢子
2.3 不同固体培养基对稻曲病菌菌丝生长的影响
测定的培养基中,稻曲病菌在PSA培养基上生长最快,大米粒培养基次之,胁本哲氏培养基质最慢(表2)。
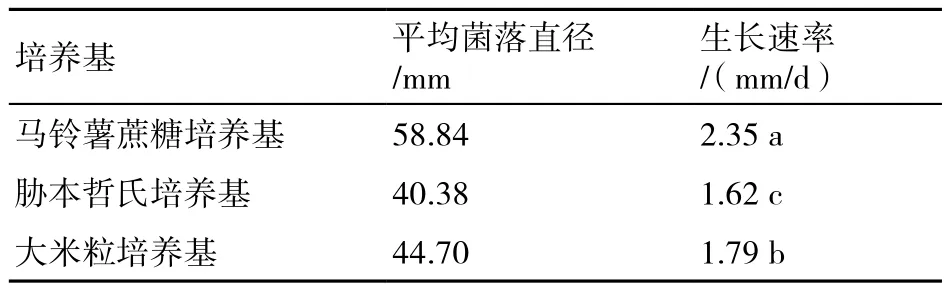
表2 不同培养基对稻曲病菌菌丝生长的影响
2.4 田间稻曲病菌菌核形成的情况
2008~2018年,随机取样调查了江西省南昌、井冈山、宜黄、鄱阳、高安、修水等多县(市)发生稻曲病的早稻、一季稻和双季稻稻田,感病程度有轻、中、重稻田,品种类型有籼、粳稻,熟期有早、中、晚熟。2008~2010年连续系统地调查了南昌与井冈山设立的南方及江西省水稻早中晚稻区域试验田播种的参试品种累计500余份(次)。结果表明,通常年份无论是早稻、一季稻和双季晚稻,也无论是籼稻或粳稻,即使在稻曲病重发田块稻曲球上也难以查找到菌核。但也有例外,2011年11月底在南昌县原省水稻良种场属地机插的近百亩稻田,主栽品种黄华占,已收获的晾晒稻子中及田间尚未收割的感染稻曲病的稻穗上均发现了一定数量的菌核。田间抽样调查500粒稻曲球,形成菌核的约占1%,多数球上菌核只有1粒,少数有2粒(图3)。但之后数年在其发生地及周边稻田均未调查到有稻曲菌核的发生。

图3 菌核形态
2.5 稻曲病菌厚垣孢子的接种效应
以当年或上一年度冷藏于冰箱中的稻曲病菌厚垣孢子为接种体,注射、喷雾接种孕穗期至破口期稻株,水稻抽穗后5~7 d,透过颖壳可看到颖内有颜色稍深的病组织形成;8~10 d病组织迅速膨大生长到颖壳外,并被一层灰白色的膜状物包裹,随后,病组织外层包膜破裂,黄色稻曲球形成。结果表明,以田间采集的稻曲球上的菌厚垣孢子为接种体,无论是注射还是喷雾接种均能诱发水稻稻曲病,可见稻曲病厚垣孢子是有效的接种体。
3 结论与讨论
采集不同色泽稻曲球上的厚垣孢子在清水和蔗糖溶液中孵育后镜检,发现厚垣孢子的萌发速率在清水和蔗糖溶液中均有显著差异,以清水孵育萌发更快。黄色稻曲球上的厚垣孢子在清水中孵育2 h即可观察到有萌动的孢子形成芽管萌发产生分生孢子,4~6 h 分生孢子产生次生分生小孢子,6~12 h分生孢子成熟脱落,孢子梗顶端产生新的分生孢子。黑色稻曲球上的厚垣孢子萌发需要在水中孵育4 h后产生分生孢子,萌发速率和萌发率均低于黄色厚垣孢子。稻曲球体上的厚垣孢子在水中的萌发特性于前期研究趋于一致[15]。但在本试验之外也观察到有少数接近黄色稻曲球上厚垣孢子萌发率的,或低于本试验结果的黑色稻曲球。王疏等[9]报道了田间采集的黄色厚垣孢子配制成孢子悬浮液在26 ℃温箱中处理10 h后开始有少量孢子萌发,到20 h后萌发率可达90%以上。李阳等[11]报道了厚垣孢子在蔗糖溶液中的萌发率高于蒸馏水中的萌发率,其振荡培养萌发产生分生孢子需要6 h。本研究结果较前人报道孢子萌发起始时间提前了4 h,这是否反映出不同地区的病菌孢子萌发特性存在差异,还是与研究者采样取材差异所导致的则有待进一步探讨。笔者检镜发现,田间鲜黄色稻曲球上的厚垣孢子可观察到有少数产生芽管的萌发现象,黑色球也存在这种现象,但不如黄色球常见,至今未见他人报道。王疏等[9]研究认为,黑色厚垣孢子不能萌发,而本研究未出现这一结果。镜检发现,突破外包膜2 d左右的稻曲球体上光泽度好的厚垣孢子萌发活力最强,之后萌发活力有所降低,但多数黄色稻曲球体上的厚垣孢子在清水中孵育3 h以上均能陆续进入萌发状态;在清水和蔗糖溶液中孵育48 h的厚垣孢子有10%以上未能萌发,李阳等[11]研究发现有不萌发的孢子存在,认为可能是不育孢子。但在此之外,这些“不育”孢子,是否可能为“休眠”孢子?其是否以休眠状态存在并经越冬越夏度过不良环境待来年休眠解除后在适宜的条件下萌发成为初侵染来源?镜检发现保存在冰箱中1年的黄色稻曲球上的厚垣孢子仍有10%左右的萌发率。王疏等[9]也报道了在4 ℃保存1年有具萌发活性的孢子存在。据此推测厚垣孢子应是病害重要的初侵染来源。
试验测试了PSA、大米粒和胁本哲氏3种培养基对稻曲病菌生产速率的影响,稻曲病菌以在PSA培养基中生长最好,大米粒培养基中生长次之,胁本哲氏培养基中生长相对较慢。这与李阳等[11]的研究报道趋于一致,但病菌培养产孢所需时间较李阳等报道的更长一些。李阳等[11]研究认为,厚垣孢子萌发必须经过分生孢子阶段才能形成菌丝或产生侵入结构。在自然情况下,厚垣孢子是病害的主要初侵染来源,可能以分生孢子及其萌发产生的菌丝侵入水稻。但前人也有报道采用稻曲病厚垣孢子接种一般难以获得成功的情况[16-19]。笔者取冷藏保存1年的稻曲球,清水洗下厚垣孢子并浸渍4 h后接种到孕穗末期、破口初期的水稻上诱发了一定比例的稻曲病株,证实厚垣孢子与其萌发产生的分生孢子、次生分生孢子及菌丝混合体是有效接种体。张俊喜等[20]采用厚垣孢子进行稻曲病接种也取得了成功。由此进一步推测厚垣孢子应是稻曲病初侵染重要来源之一。但也有学者认为菌核在稻曲病发生流行中可能发挥了主导作用[5,21-22]。这是否意味着稻曲病的发生与厚垣孢子及菌核均有关联,如何定论值得商榷。
王永强等[23]研究认为,在我国北方稻区,昼夜温差较大,产生菌核的现象相对比较普遍。而在浙江、安徽、福建、广西、四川等南方稻区采集的标样中,稻曲球上产生菌核的现象较为罕见。然而,南方稻区早年与近年也有田间稻曲球上产生菌核的报道[5,13-14]。笔者针对稻曲病菌核田间发生情况于不同年份在江西省多县(市)进行了调查,结果通常是少见稻曲球上产生菌核。但2011年在南昌县迟熟的染病稻穗上的稻曲球上找到了一定数量的菌核。当年9月中下旬江西持续多日相对低温天气,水稻生育期延长,尤其是机栽晚稻较人工栽插的生育期延长了近半月之久,导致11月底田间还未收割完晚稻,菌核在晚插迟熟的感病稻穗上的稻曲球中出现。故此推测稻曲病菌核的形成应在稻曲球形成后田间有一个相对低温的过程,当满足了稻曲病菌核形成的温度、湿度条件时,可促使球体致密菌丝体向菌核形成转化,从而产生一定数量的菌核。有关低温诱导稻曲病菌产生菌核前人已有报道[5,22-24],但南宁地区在没有明显的异于往年的寒冷低温天气情况下田间也发现有菌核形成[14],由此看来菌核的形成机制尚不明确,与气温的相关性还有待进一步研究论证。
猜你喜欢
农家致富顾问·上半月(2022年5期)2022-05-29
中国药学药品知识仓库(2021年11期)2021-11-25
中小学德育(2020年11期)2020-03-18
江苏农业科学(2019年11期)2019-07-22
热带作物学报(2019年4期)2019-06-11
安徽农业科学(2018年2期)2018-05-14
安徽农学通报(2017年23期)2017-12-27
教育界·上旬(2016年12期)2017-05-25
环球人文地理(2016年10期)2016-11-24
江苏农业科学(2015年5期)2015-10-20
