
miR-146b-5p对骨肉瘤细胞增殖抑制及克隆形成的机制研究
2022-06-27 02:24姜富祥阿尔宾
现代检验医学杂志 2022年3期
姜富祥,阿尔宾,高 飞,王 兴
(内蒙古自治区巴彦淖尔市医院脊柱外科,内蒙古巴彦淖尔 015000)
骨肉瘤(osteosarcoma,OS)是一种高度恶性的骨架癌,起源于间充质组织中梭形细胞的未成熟类骨质基质,是青少年最常见的原发恶性骨肿瘤[1-2]。目前手术、放化疗是OS的主要治疗方式,针对OS的靶向治疗药物仍在研究[3],然而临床治疗效果有限,故深入研究OS的发生发展机制,找寻针对OS的新的有效治疗方法及分子标志物意义重大。微小核糖核酸分子(micro RNA,miRNA)是具有15~25个核苷酸的小非编码RNA(small non-coding RNA)调控分子,其可与基因3’UTR区结合发挥功能,具有广泛的生物学功能,研究证实miRNA参与了细胞凋亡、新陈代谢、发育、增殖和细胞分化等生物学过程[4-7]。近年发现,miR-146b-5p与许多人类恶性肿瘤有关,例如在乳腺癌中,lncRNA NEAT1通过抑制miR-146b-5p表达可促进人乳腺癌细胞的增殖、迁移和转移[8]。miR-146b-5p通过调节IRAK1/NF-κB通路提高非小细胞肺癌(non-small cell lung cancer,NSCLC)对表皮生长因子受体(epidermal growth factor receptor, EGFR)酪氨酸激酶抑制剂的敏感度[9]。证实miR-146b-5p可抑制上皮间质转化及细胞迁移和侵袭等多种致癌途径[10-12],在人类多种肿瘤的发生发展中扮演重要角色,可作为新的肿瘤分子研究靶标。有研究显示,miR-146b-5p在OS中过表达可显著抑制肿瘤细胞增殖,促进其凋亡,但miR-146b-5p在OS细胞增殖中的相关分子机制尚在探索阶段。故本研究通过体外细胞试验拟探究miR-146b-5p在OS中的表达与其对OS细胞增殖和克隆形成的影响及相关分子机制,以期为OS的临床诊治提供新的研究靶标。
1 材料与方法
1.1 研究对象 人骨肉瘤细胞U2OS细胞购于中国科学院上海细胞库,培养在含10g/dl胎牛血清DMEM培养基中,加入青霉素100 IU/ml和链霉素100 µg/ml,在37℃,5ml/dl CO2培养箱中培养。选取2018年1月~2020年12月内蒙古自治区巴彦淖尔市医院病理科留存的30例骨肉瘤组织及对应正常组织样品,均经病理确认为骨肉瘤,术前未行任何相关治疗,组织样本来源于手术获取,低温液氮保存备用。
1.2 仪器及试剂 DMEM培养液(美国HyClone公司);链霉素、青霉素、胰酶(美国Gibco公司);Lipofectamine 2000(北京优尼康生物科技有限公司);酶标仪(瑞士Tecan公司);MTS细胞增殖检测试剂盒、克隆形成检测试剂盒(美国corning公司);PCR仪(美国ABI公司);逆转录试剂盒[生工生物工程(上海)股份有限公司];双荧光素酶测定系统(北京Promega公司);蛋白免疫印迹实验电泳仪、转膜槽(美国Bio-Rad公司),DUSP16抗体(美国SigmaAldrich公司);miR-146b-5p引物 序 列,miR-146b-5p mimics,miR-146b-5p ASO(反义核酸)及siRNA-DUSP16和无义序列siCTL由上海吉玛生物制药有限公司设计合成;DUSP16-3’UTR-WT野生型质粒和DUSP16-3’UTR-MUT突变型质粒报告基因载体(美国Promega公司)。
1.3 方法
1.3.1 细胞转染及分组:待细胞生长密度达80%左右时,按照Lipofectamine 2000试剂盒说明书将NC-mimics,miR-146b-5p mimics,NC-ASO,miR-146b-5p-ASO及siRNA-DUSP16和siCTL分别转染至U2OS细胞,设为miR-146b-5p阴性过表达组(NC-mimics组),miR-146b-5p过表达组(miR-146b-5p mimics组)和miR-146b-5p阴性抑制组(NC-ASO),miR-146b-5p抑制组(miR-146b-5p-ASO组)及DUSP16敲低表达组(shDUSP16),DUSP16对照组(siCTL组);更换DMEM培养液培养48 h,qRT-PCR验证转染效率。
1.3.2 实时荧光定量PCR实验(qRT-PCR):取转染后各组细胞采用Trizol试剂盒提取细胞总RNA,反转录为cDNA,以此为模板配置RCR反应体系行RT-PCR扩增分析。miR-146b-5p引物序列:上 游5’- CGTGCCACTGACAAGTGAAT-3’,下 游5’-CGACCAGAACTCAGTCGACA-3’;内参GAPDH上 游5’- GTCACAGCATTTGCTCGTATTG-3’,下 游5’-CTCCTGCAACATCGTGATCGG-3’;反应条件:95 ℃ 2 min,95 ℃ 15 s,60 ℃ 30 s,72 ℃ 10 s,40个循环。采用2-ΔΔCt法计算目的基因相对表达。试验重复三次取平均值。
1.3.3 细胞增殖实验:取对数生长期各组细胞以1×107个/孔接种于96孔板,每孔设3个生物复孔,37 ℃,5ml/dl CO2孵育待细胞贴壁后进行转染;配制MTS反应液,每孔加入MTS液 100 μl,继续培养48 h,在490 nm处测量各孔吸光度值(A值),绘制细胞生长曲线,试验重复三次取平均值。
1.3.4 克隆形成实验:待各组细胞贴壁后进行转染,每孔设3个生物复孔,培养至出现肉眼可见的克隆时,终止培养,收集细胞,PBS洗涤两次,去除多余洗涤液,用含0.5g/dl结晶紫的甲醇溶液固定15 min,水洗、晾干,Giemsa染色30 min,在590 nm处测A值,试验重复三次取平均值。
1.3.5 双荧光素酶报告基因实验:将转染后各组细胞接种到6孔板,待其生长密度为90%时将含有海肾荧光素酶的DUSP16-3’UTR-WT野生型和DUSP16-3’UTR-MUT突变型质粒及miR-146b-5p转染到细胞中,采用双荧光素酶测定系统进行荧光素酶活性测定,试验重复三次取平均值。
1.3.6 蛋白免疫印迹实验:取对数生长期各组细胞加入裂解液提取总蛋白,并测定其浓度及纯度;取定量蛋白制样,SDS-PAGE电泳,转膜,5g/dl脱脂牛奶室温封闭1 h,加入一抗4 ℃孵育过夜;洗膜3次,加入二抗室温孵育1 h,洗膜3次,化学发光,暗房显影,凝胶成像系统拍照分析。
1.3.7 miR-146b-5p靶标预测:通过microRNA网站进行预测miR-146b-5p与DUSP16的结合位点,行进一步研究。
1.3.8 DUSP16与骨肉瘤预后的相关性:通过公共osteosarcoma R2数据库http://r2.amc分析DUSP16与骨肉瘤预后的关系。
1.4 统计学分析 应用SPSS 20.0统计软件进行数据分析,数据以均数±标准差(±s)表示,两组间差异比较采用t检验;多组间差异比较采用oneway ANOVA分析,组间两两比较采用LSD-t检验;骨肉瘤组织及其对应正常组织中表达差异比较采用配对t检验;P<0.05为差异有统计学意义。
2 结果
2.1 骨肉瘤中miR-146b-5p表达 见图1。qRTPCR检测显示,30组骨肉瘤组织中miR-146b-5p表达(4.096±1.237)较邻近正常组织(5.895±0.834)显著降低,差异有统计学意义(t=6.605,P<0.001)。
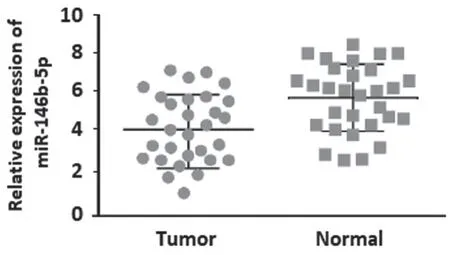
图1 miR-146b-5p在骨肉瘤组织及邻近正常组织中的表达
2.2 过表达和敲低miR-146b-5p效率验证 qRTPCR检测显示,转染miR-146b-5p mimics组细胞中miR-146b-5p表达(4.562±0.026)较NC-mimics组(1.003±0.002)明显升高,差异有统计学意义(t=236.393,P<0.001);转染miR-146b-5p-ASO组细胞中miR-146b-5p表达(0.394±0.002)较NCASO组(1.251±0.003)明显降低,差异有统计学意义(t=411.689,P<0.001),表明过表达和敲低miR-146b-5p表达的骨肉瘤细胞系构建成功,可用于后续实验。
2.3 miR-146b-5p对骨肉瘤细胞增殖及克隆形成的影响 见表1。增殖实验检测显示,miR-146b-5p过表达明显抑制了骨肉瘤细胞的增殖能力(t=13.233~34.599,均P<0.01),敲低miR-146b-5p表达则显著促进了骨肉瘤细胞的增殖(t=7.610~9.247,均P<0.01),差异均有统计学意义。细胞克隆实验检测显示,miR-146b-5p mimics组细胞克隆形成率(48.912±2.032)较NCmimics组(160.834±9.031)明显降低,差异有统计学意义(t=20.942,P<0.001);miR-146b-5p-ASO组细胞克隆形成率(239.658±11.034)较NCASO组(128.502±2.413)明显升高,差异有统计学意义(t=17.046,P<0.001)。
表1 过表达和敲低miR-146b-5p对骨肉瘤细胞增殖的影响(A值,± s)
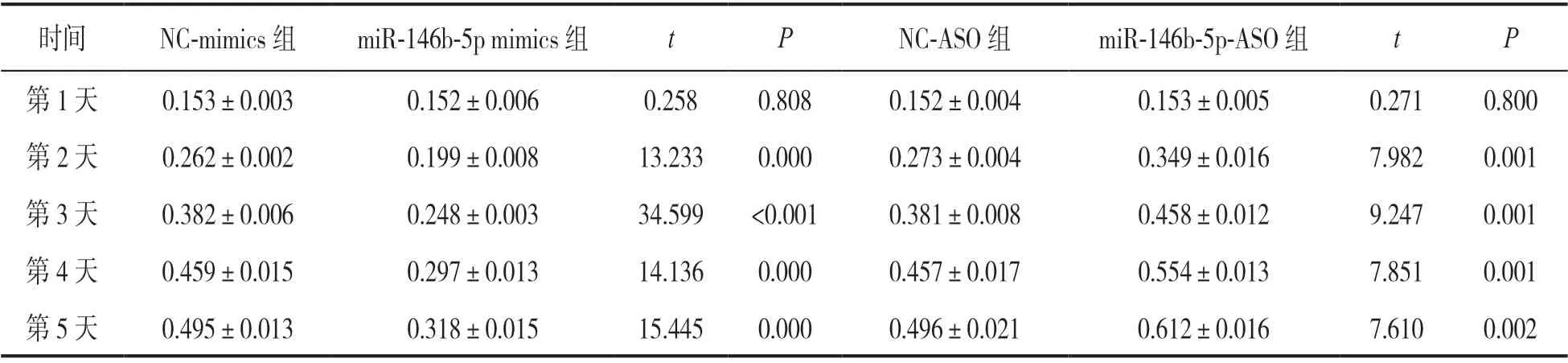
表1 过表达和敲低miR-146b-5p对骨肉瘤细胞增殖的影响(A值,± s)
时间 NC-mimics组 miR-146b-5p mimics组 t P NC-ASO组 miR-146b-5p-ASO组 t P第1天 0.153±0.003 0.152±0.006 0.258 0.808 0.152±0.004 0.153±0.005 0.271 0.800第2天 0.262±0.002 0.199±0.008 13.233 0.000 0.273±0.004 0.349±0.016 7.982 0.001第3天 0.382±0.006 0.248±0.003 34.599 <0.001 0.381±0.008 0.458±0.012 9.247 0.001第4天 0.459±0.015 0.297±0.013 14.136 0.000 0.457±0.017 0.554±0.013 7.851 0.001第5天 0.495±0.013 0.318±0.015 15.445 0.000 0.496±0.021 0.612±0.016 7.610 0.002
2.4 miR-146b-5p靶基因的预测验证及调控关系见图2。 检索microRNA网站发现,miR-146b-5p与DUSP16 3’UTR区域存在靶向结合位点(图2a)。研究将DUSP16-3’UTR-WT和DUSP16-3’UTRMUT质粒载体及miR-146b-5p 共转染至U2OS细胞,经双荧光素酶实验验证显示,转染miR-146b-5p mimics组DUSP16-3’UTR-WT荧光素酶活性(0.394±0.002)较NC-mimics组(1.001±0.002)显著降低,差异有统计学意义(t=371.710,P<0.001);而DUSP16-3’UTR-MUT荧光素酶活性(0.998±0.003)较NC-mimics组(1.001±0.001)无显著改变,差异无统计学意义(t=1.643,P=0.176)(图2b),证实DUSP16是miR-146b-5p的潜在靶基因。进一步发现,miR-146b-5p mimics组细胞中DUSP16表达(0.231±0.013)较NC-mimics组(1.011±0.002)明显降低,差异有统计学意义(t=102.715,P<0.001),相反的miR-146b-5p-ASO组细胞中DUSP16表达(3.264±0.026)较NC-ASO组(1.009±0.003)明显升高,差异有统计学意义(t=149.232,P<0.001),说明miR-146b-5p负向调控DUSP16表达。
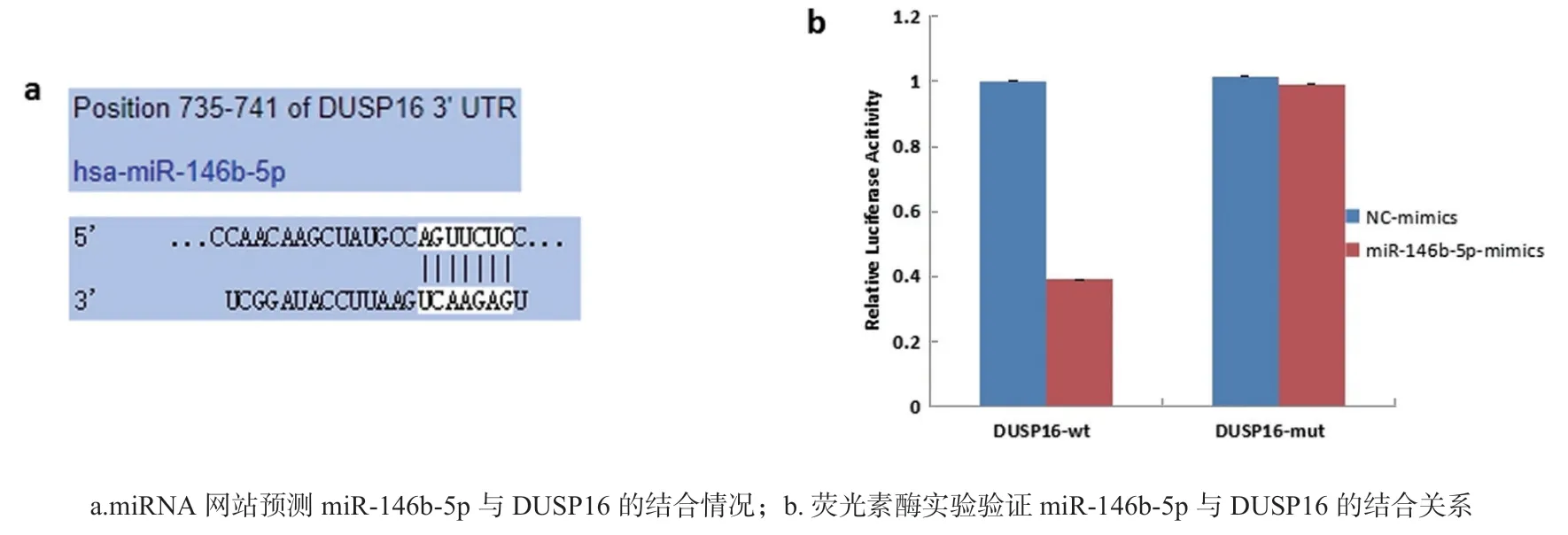
图2 miR-146b-5p靶基因的预测及验证
2.5 DUSP16在骨肉瘤中的表达特征及miR-146b-5p与DUSP16的调控关系验证 见图3,表2。30组骨肉瘤组织中DUSP16表达(5.683±0.457)相比邻近正常组织(4.665±0.531)显著高表达,差异有统计学意义(t=7.959,P<0.001),与miR-146b-5p在骨肉瘤中的表达呈显著负相关(r=-0.667,P<0.05);通过osteosarcoma R2数据库http://r2.amc分析发现DUSP16具有明显的高表达预后差特征(图3a)。细胞增殖实验证实,过表达miR-146b-5p显著抑制细胞增殖,在miR-146b-5p mimics组细胞中过表达DUSP16后细胞增殖速率回归到正常水平,在NC-mimics组中过表达DUSP16后细胞增殖速率较之前明显升高(图3b,表2),说明miR-146b-5p通过负调控DUSP16表达在骨肉瘤细胞中发挥功能。
表2 miR-146b-5p调控DUSP16对细胞增殖的影响(A值,±s)
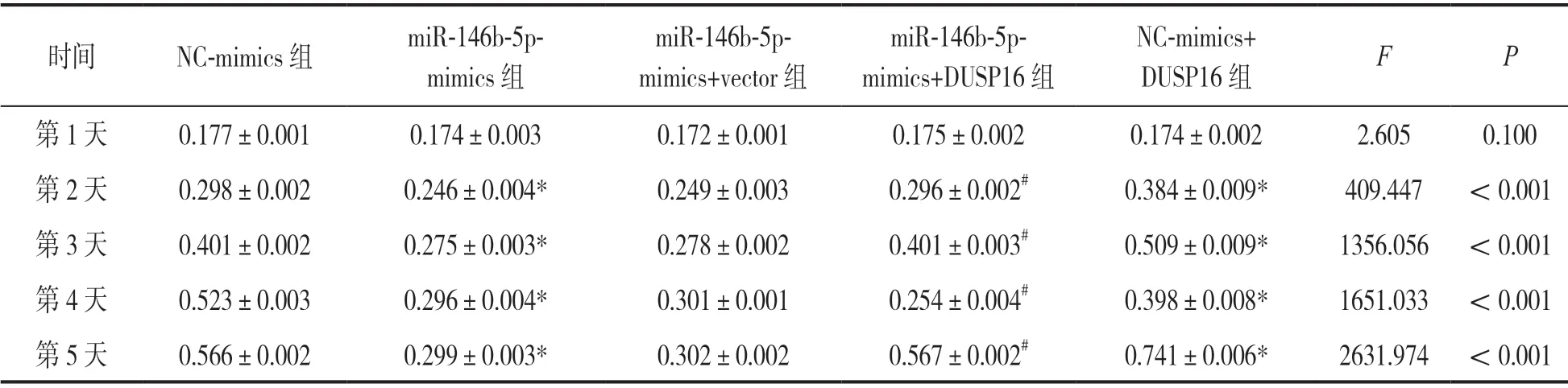
表2 miR-146b-5p调控DUSP16对细胞增殖的影响(A值,±s)
注:*与NC-mimics组相比,t=13.338, 33.359, 60.381, 96.851; 12.568, 32.564, 59.051, 95.763; 22.059, 28.593, 33.250, 63.479, 均P<0.001;#与miR-146b-5p-mimics组相比,t=12.825, 33.359, 11.172, 97.214, 均P<0.001。
时间 NC-mimics组 miR-146b-5pmimics组miR-146b-5pmimics+vector组miR-146b-5pmimics+DUSP16组NC-mimics+DUSP16组 F P第1天 0.177±0.001 0.174±0.003 0.172±0.001 0.175±0.002 0.174±0.002 2.605 0.100第2天 0.298±0.002 0.246±0.004* 0.249±0.003 0.296±0.002# 0.384±0.009* 409.447 <0.001第3天 0.401±0.002 0.275±0.003* 0.278±0.002 0.401±0.003# 0.509±0.009* 1356.056 <0.001第4天 0.523±0.003 0.296±0.004* 0.301±0.001 0.254±0.004# 0.398±0.008* 1651.033 <0.001第5天 0.566±0.002 0.299±0.003* 0.302±0.002 0.567±0.002# 0.741±0.006* 2631.974 <0.001

图3 骨肉瘤中DUSP16表达特征及其与miR-146b-5p的调控关系验证
2.6 miR-146b-5p调控DUSP16参与P53信号通路在骨肉瘤中发挥作用 见图4。研究经介导转染siRNA敲低DUSP16表达,发现siDUSP16组P53磷酸化水平(1.342±0.019)较转染无义序列siCTL组(0.452±0.013)显著升高(t=66.960,P<0.001),同时P53靶基因P21(肿瘤抑制子)表达(3.136±0.145)也较siCTL组(1.110±0.014)显著升高(t=24.089,P<0.001),差异均有统计学意义(图4a),说明在骨肉瘤中DUSP16可参与调控P53信号通路。进一步验证发现,miR-146b-5p mimics组细胞中P21表达(4.995±0.214)较NC-mimics组(1.001±0.002)显著升高(t=32.325,P<0.001);在过表达miR-146b-5p组细胞中再次过表达DUSP16后,P21表达(0.986±0.012)较单独过表达miR-146b-5p组显著降低(t=32.398,P<0.001)(图4b),差异均有统计学意义。由此说明miR-146b-5p通过靶向调控DUSP16表达在骨肉瘤中发挥作用是通过P53信号通路来实现的。
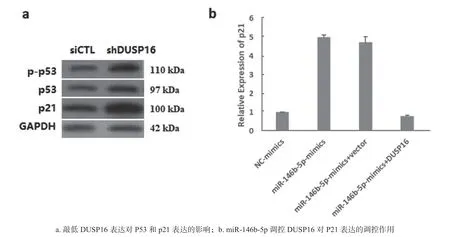
图4 miR-146b-5p调控DUSP16参与P53信号通路在骨肉瘤中发挥作用
3 讨论
骨肉瘤(OS)占儿童恶性肿瘤的2.4%,恶性程度高,易侵犯周围软组织,常发生远处转移至肺及骨骼等部位,约10%~20% OS患者诊断时已发生早期转移,预后明显恶性[13]。且据报道,非转移性OS患者5年生存率可达80.5%,转移性OS患者5年生存率仅为15%~30%[14]。故基于分子生物学角度探究OS发生、转移的分子机制,找寻新的有效的OS治疗靶标对提高患者预后意义显著。
近年研究发现miRNA可通过碱基互补方式与靶基因3’-UTR区结合,降解mRNA或抑制mRNA翻译,在肿瘤发生发展过程中调节细胞多种生物学过程,参与肿瘤的恶性发展[15,6,8]。miRNAs表达失调是肿瘤发生及转移的诱因[8]。大量研究证实,不同miRNAs异常表达与OS细胞增殖、迁移和侵袭及不良预后相关,如miRNA-505,miR-210-5p和microRNA-320a等[16-18],由此可见mRNAs在OS中的发生及转移中起着重要作用。miR-146b是近年研究新发现的一种miRNA,位于第10号染色体,包含miR-146b-3p和miR-146b-5p两种形式,其中后者在人肝脏、脾脏组织中呈不同程度高表达[12]。而在肿瘤的研究中发现,miR-146b-5p缺失通过IL-17A途径能够促进T细胞急性淋巴细胞白血病的迁移和侵袭[19]。通过碘参与下调miR-146b-5p可抑制甲状腺乳头状癌细胞的增殖[10]。lnc-AL445665.1-4可能通过与miR-146b-5p的相互作用参与了子宫多发平滑肌瘤的发生发展[11]。lncRNA-NEAT1通过miR-146b-5p/NOTCH1信号通路促进了T细胞急性淋巴母细胞白血病(T-ALL)细胞的增殖[20]。此外还发现,miR-146b-5p在乳腺癌、胰腺癌、前列腺癌等人类恶性肿瘤中表达下调[21],提示其可能作为抑癌基因发挥作用,参与恶性肿瘤的发展进程,可作为肿瘤治疗的新靶点。本研究显示,骨肉瘤组织中miR-146b-5p显著低表达,其过表达能够抑制OS细胞的增殖和克隆形成,发挥抑癌基因特性,故探究其发挥作用的分子机制意义重大。
本研究检索生物学信息数据库发现miR-146b-5p与DUSP16存在结合位点,经荧光素酶实验证实DUSP16是miR-146b-5p的靶基因。研究表明,双重特异性磷酸酶16(DUSP16,也称为促分裂原活化的蛋白激酶磷酸酶7)通过减弱磷酸丝氨酸/苏氨酸和磷酸酪氨酸残基(例如p38和JNK途径21)的磷酸化可抑制其靶激酶的活化[22]。DUSP16表达调节能够控制ERK和p38 MAP激酶活性,介导化疗诱导的乳腺癌干细胞富集[23]。靶向DUSP16/TAK1信号通过抑制JNK MAPK减轻高脂饮食(HFD)小鼠的肝脏血脂异常和炎症[24]。并发现DUSP16表达上调可促进人类肝癌细胞的增殖[25]。CircDUSP16通过调控miR-497-5p/TKTL1轴促进食管鳞癌细胞发育[26]。CircDUSP16通过海绵化miR-145-5p促进了胃癌的发生和侵袭[27]。而本研究中发现骨肉瘤组织中DUSP16显著高表达,具有明显的高表达预后差特征,与miR-146b-5p存在显著负相关,暗示了DUSP16的癌基因属性。经细胞实验检测证实, DUSP16过表达促进了OS细胞增殖,miR-146b-5p对OS细胞的增殖调控与其对DUSP16表达的调控相关。作用机制方面,既往研究报道在三阴性乳腺癌化疗过程中,DUSP16 表达显著降低导致p38被激活,从而促进了化疗耐药乳腺癌干细胞表型的规范,为临床治疗提供了新靶点[23]。既往HAO等[28]也报道,小鼠模型中DUSP1可抑制p38 MAPK磷酸化,导致HCC细胞中P53磷酸化增强,从而激活p53和p21并诱导其表达,促进肝细胞癌细胞的凋亡。所以本研究进一步探究了miR-146b-5p调控DUSP16对P53信号通路的影响,发现敲低DUSP16可促使P53磷酸化及P53靶基因P21表达显著升高,证实miR-146b-5p调控DUSP16可能通过P53信号通路参与了OS的发生发展。故继续探究miR-146b-5p调控DUSP16对P53信号通路的影响机制可为OS的诊断及治疗提供更加有效的参考靶标。
综上所述,OS中miR-146b-5p低表达通过靶向负调控DUSP16参与OS细胞的增殖及克隆形成,与P53信号通路关系密切。
猜你喜欢
中国现代医生(2022年19期)2022-11-04
中国医学物理学杂志(2022年9期)2022-10-09
环球时报(2022-09-20)2022-09-20
中国现代医生(2022年19期)2022-08-25
江苏农业科学(2019年23期)2019-03-03
中国中医药信息杂志(2018年11期)2018-01-05
三联生活周刊(2017年1期)2017-01-11
医学信息(2016年29期)2016-11-28
小资CHIC!ELEGANCE(2015年14期)2015-09-23
小资CHIC!ELEGANCE(2015年15期)2015-09-01
