
双乾肉羊BMP3 基因多态性及其与背最长肌脂肪酸含量的相关性研究
2022-06-13 12:36梁应嘉杨少滢金海国
中国畜牧杂志 2022年6期
崔 超,马 骏,张 鑫,梁应嘉,杨少滢,3,肖 成,刘 宇,金海国,曹 阳*
(1.吉林省农业科学院,吉林长春 130033;2.吉林农业大学动物科学技术学院,吉林长春 130118;3.延边大学农学院,吉林延吉 136200)
随着生活质量的提高,高品质羊肉越来越受到消费者欢迎,如何提高羊肉品质以及选育高品质肉羊品种是目前需要解决的一大难题。家畜的肉质与脂肪的含量密切相关,适量的肌内脂肪含量可增加大理石花纹,降低剪切力,增加肉质风味及适口性。影响肌内脂肪的关键因素是遗传因素。
骨形态发生蛋白3(Bone Morphogenetic Protein 3,)是转化生长因子-(TGF-)超家族的成员。目前关于基因的研究主要在调控心脏、骨骼、肾脏和胚胎发育以及母畜的生殖过程等方面,基因在成骨组织的形成和损伤后的修复过程中也起着重要作用,同时有部分报道证明基因在脂代谢中扮演着重要角色。本课题以双乾肉羊为研究对象,通过Sanger 测序检测基因第一外显子的多态性,并综合分析其与绵羊背最长肌脂肪酸相对含量之间的相关性,以期为后续探索影响肉质性状的候选基因提供素材。
1 材料与方法
1.1 实验动物 实验所用双乾肉羊均来自吉林省农业科学院试验羊场,随机选取健康且体况良好的双乾肉羊49 只,断奶后饲养6 个月屠宰,采集背最长肌进行脂肪酸含量检测,采集肝组织,-80℃保存备用。
1.2 主要仪器及试剂 动物组织DNA 快速提取试剂盒购自Axygen 公司;2×ES Taq Master Mix 购自北京康为世纪生物科技有限公司;Q5000 超微量分光光度计购自美国Quawell 公司;PCR 仪购自上海伯乐生命医学产品有限公司。
1.3 组织样DNA 提取 从肝组织中提取绵羊基因组DNA,并使用超微量分光光度计测定提取的DNA 浓度和纯度,将DNA 样品进行-80℃保存。
1.4 引物设计及合成 根据NCBI 网站所预测的羊基因(XM_027971127.2)外显子序列,采用Primer 5.0 软件设计基因第一外显子引物,并由苏州金唯智生物科技有限公司合成。引物序列为F:5'-ACATCAGCCAGGGAACCGA-3';R:5'-GCGACTGTC TAAAGGGCTAAGG-3',扩增目的片段长度为1 494 bp。
1.5 绵羊基因PCR 产物扩增 PCR 反应体系如下:2×ES Taq Master Mix 10 μL(1×),上、下游引物(0.5 μmol/L)各0.5 μL,DNA(<250 ng/50 μL)1 μL,ddHO 补足至20 μL。PCR 反应过程如下:95℃预变性3 min,95℃变性30 s,适宜退火温度退火30 s,72℃延伸45 s,总共进行34 个循环,72℃延伸5 min,产物4℃保存。将PCR 产物取5 μL 进行1.5% 琼脂糖凝胶电泳检测引物特异性。
1.6 绵羊基因多态性测序及分析 把扩增出的、条带明亮的PCR 产物送到苏州金唯智生物科技有限公司测序。使用DNAMAN 和Chromas 软件通过对测序结果的序列及序列图谱进行比对分析,进一步筛选SNP位点,统计并计算不同基因型频率。
1.7 肌肉脂肪酸含量的检测 用气相色谱法进行肌肉脂肪酸的测量,方法参照“肉与肉制品脂肪酸测定”(GB/T9695.2-2008)。
1.8 数据统计与分析 利用家养动物遗传分析工具计算群体遗传参数、检验,并分析判断基因型是否处于哈代-温伯格平衡状态。采用SPSS 23.0 软件中One-Way ANOVA 对实验数据进行单因素方差分析并用LSD 法进行多重比较,以明确不同基因型与双乾肉羊肌肉脂肪酸含量的相关性,实验数据用平均值±标准差表示,差异显著性标准以<0.05 来判断。
2 结果
2.1 PCR 扩增产物 由图1 可知,扩增片段长度约为1 494 bp,与预期大小一致,条带明亮、单一、无杂带,符合后续实验要求。
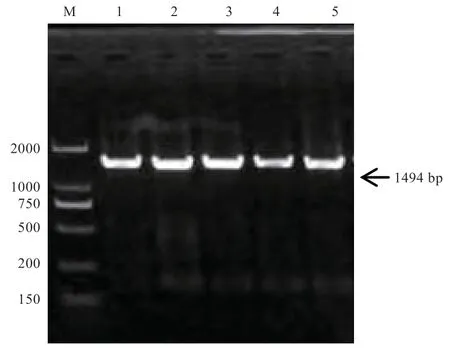
图1 BMP3 基因第1 外显子的PCR 扩增电泳图
2.2 绵羊基因PCR 产物测序及序列分析 测序结果通过DNAMAN 软件比对分析发现SNPs 位点,在基因第1 外显子276 bp 处发现G →A 突变(图2),然后使用下游引物将49 个样单独测序,并用Chromas软件对测序峰图逐一比对,发现其有A 和G 2 个等位基因,AA、AG 和GG 3 种基因型(图3)。

图2 绵羊BMP3 基因外显子1 核苷酸序列比对
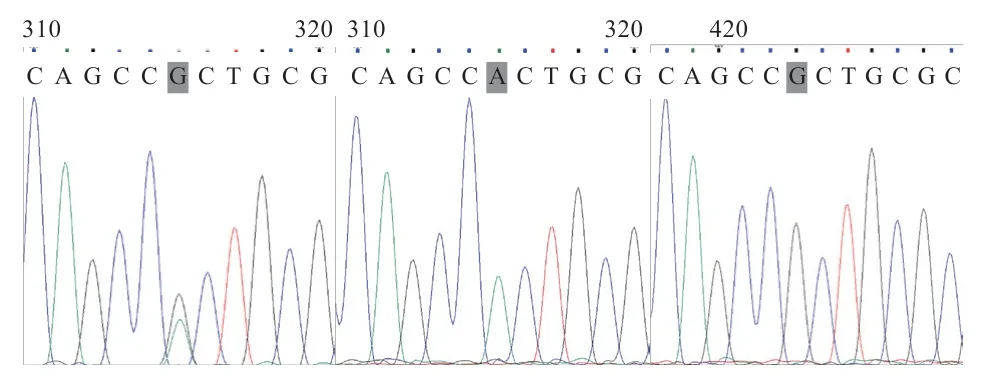
图3 绵羊BMP3 基因外显子1 AA、AG 和GG 基因型的测序峰图
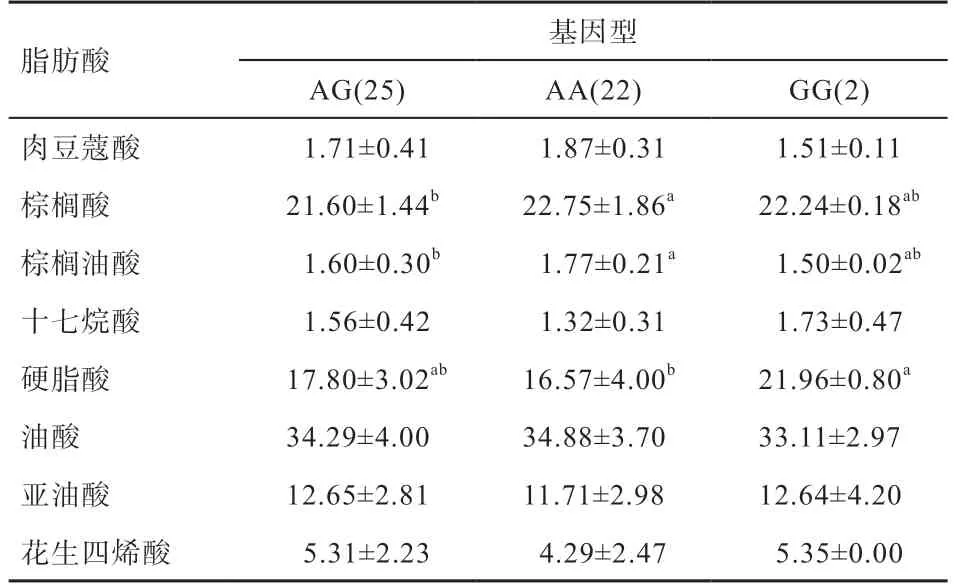
2.3 绵羊基因多态位点遗传多样性分析 由表1 可知,绵羊群体基因第1 外显子AA、AG 和GG 基因型频率分别为44.9%、51.0% 和4.1%,因此AG 型为优势基因型。A、G 基因频率分别为70.4% 和29.6%,因此A 为优势等位基因。根据卡方适合性检验结果并对照值表,判断该位点在双乾肉羊群体中处于哈代-温伯格平衡状态(>0.05)。经计算,杂合度(He)、纯合度(Ho)和有效等位基因数(Ne)分别为0.420 0、0.580 0 和1.724,该位点属于中度多态(0.25 表1 绵羊BMP3 基因不同突变位点基因型频率及基因频率 2.4基因不同基因型与绵羊肌肉脂肪酸含量的相关性 由表2 可知,基因外显子1 的不同基因型个体间肌肉脂肪酸含量存在显著性差异,绵羊基因A/G 位点3 种不同基因型对棕榈油酸、棕榈酸和硬脂酸含量均有显著性影响。其中AA 型的棕榈酸和棕榈油酸含量在该群体中均显著高于AG 型,与GG 型个体间不存在显著差异。GG 基因型的硬脂酸含量在该群体中显著高于AA 型,与AG 型个体间不存在显著差异。其余脂肪酸含量在3 种基因型间不存在显著性差异。 表2 绵羊BMP3 基因多态性与脂肪酸关联分析 骨形态发生蛋白基因属于TGF-超家族,是一类都具有骨及软骨诱导活性但理化性质、分子结构及功能特点不尽相同的蛋白质,参与骨和软骨形成、胚胎发生、肌肉生长、脂肪形成、生殖系统发育甚至癌症的发生。基因是一个多功能基因,在调控心脏、骨骼及成骨组织的形成和损伤后的修复过程、肾脏和胚胎发育以及母畜的生殖过程中起重要作用,并对生殖、牙根的发育以及胎儿发育起着重要的调控作用。刘建等研究表明,在骨折修复的早期诱导成骨中起重要作用。此外,也是一种肿瘤抑制因子,其活性丧失与甲状腺乳头状癌、胆管癌的发生发展有关。张小辉等发现在牛卵巢、肝等12 个组织中均有表达,并且脂肪组织中表达相对较高,这也说明可能参与脂肪代谢过程。进一步研究发现,BMP3/WISP1 信号通路在骨髓间充质干细胞增殖和脂肪生成中起着关键作用。脂肪含量是肉质性状的重要指标之一,与口感、风味密切相关。 本研究选取49 只双乾肉羊作为研究对象,根据测序结果统计发现基因第1 外显子276 bp 处存在G/A 突变,优势基因型为AG 型,该位点在所测的双乾肉羊群体中处于哈代-温伯格平衡,说明该群体基因型几乎不受人工选择干涉的影响,随着群体中随机交配的进行,遗传组成将保持不变,从而容易达到遗传平衡状态。群体遗传学分析显示,该位点属于中度多态(0.25 肌肉中脂肪酸组成与肉品质紧密相关,如肌内脂肪含量与棕榈酸呈明显正相关,与亚油酸呈明显负相关;部分脂肪酸之间也相互影响,如肉豆蔻酸和棕榈酸呈明显正相关,亚油酸与肉豆蔻酸和棕榈酸均呈明显负相关。棕榈油酸(9-十六碳烯酸)含量与肉的风味之间存在着较高的正相关性,硬脂酸(十八碳酸)含量和口味之间有较高的负相关性,可能影响肉的膻味。本研究在绵羊群体中检测不同基因型在肉中脂肪酸含量的差异,结果显示,AA 基因型双乾肉羊背最长肌中棕榈酸和棕榈油酸显著高于AG 基因型个体,GG 基因型个体硬脂酸显著高于AA 型,说明AA 基因型个体肉的风味品质更高。本实验初步推断基因多态性对绵羊肉质性状有一定影响,下一步实验应扩大样本群体数进一步分析和验证。 双乾肉羊基因第1 外显子276 bp 处发现一个A/G 突变属同义突变;AA 基因型个体背最长肌棕榈油酸和棕榈酸含量显著高于AG 基因型,GG 基因型个体硬脂酸含量在双乾肉羊群体中显著高于AA 型,其余脂肪酸含量在3 种不同基因型间不存在显著性差异。综上,基因多态性对绵羊肌肉脂肪酸含量有一定影响,AA 型个体肉的风味品质更高,在生产中具有优势,而GG 型个体可能与肉的膻味有关,相关结果可为后续绵羊肉质性状分子选育提供可参考的遗传标记。
3 讨论
4 结论
猜你喜欢
作物学报(2022年12期)2022-10-14中国农业科学(2022年16期)2022-09-19中国农业科学(2022年15期)2022-08-09安徽农学通报(2022年6期)2022-04-07电脑报(2020年40期)2020-11-06农民致富之友(2020年2期)2020-02-12农民致富之友(2019年9期)2019-05-22新课程·下旬(2018年8期)2018-11-10电脑知识与技术(2018年19期)2018-11-01农村百事通(2018年3期)2018-03-14
