
人为活动干扰对甘南高寒草甸喜马拉雅旱獭食性的影响
2022-06-02 01:37:50张彩军王小燕刘倩倩蒲强胜王志成苏军虎
草地学报 2022年5期
张彩军,王小燕,刘倩倩,安 康,蒲强胜,王志成,苏军虎
(甘肃农业大学草业学院/草业生态系统教育部重点实验室/甘肃省草业工程实验室/中美草地畜牧业可持续发展研究中心,甘肃农业大学-新西兰梅西大学草地生物多样性研究中心,国家林业草原高寒草地鼠害防控工程技术研究中心,甘肃 兰州 730070)
野生动物是草地生态系统的重要组成部分,具有重要的生态作用,对系统的结构、过程和功能的发挥会产生极大影响[1]。长期以来,人类经济社会发展的同时,人类活动对自然生态环境产生了影响,给野生动物的生存也带来了严峻挑战[2]。通常,野生动物会采取相应策略来应对人为活动的影响,如黑嘴鸥(Larussaundersi)[3]和中华竹鼠(Rhizomyssinensis)[4]会选择在远离人类活动的地方栖息。而一些动物则会被迫与人类共存,野生动物协同其他生存策略,会将人为活动干扰带来的不利影响降到最低。如东美松鼠(Sciuruscarolinensis)在人类活动水平较高的生境下,其警戒距离要显著小于人类活动水平较低生境的警戒距离,通过调整它们对人类的行为反应,以最大限度地减少非捕食性生物干扰的影响[5]。也有研究显示当受到人为活动干扰时,砺鹬(Haematopusostralegus)用于警戒的时间增加,而取食时间减少,其通过加大啄食速率、牺牲停息时间来补偿这种损失[6]。雪雁(Chencaeruluscens)则采用夜间觅食的方式来应对白天人为活动干扰带来的影响[7]。野生动物响应人为活动干扰的策略有利于自身及其种群的存活,以往的研究都侧重于野生动物应对人为干扰的行为适应,而较少从食性角度出发探讨野生动物在人为活动干扰下的响应机制。
食物是动物生存和繁殖所需要营养的来源,动物在其长期的进化过程中形成自身特有的食性[8]。动物的食性受栖息地类型、食物资源的分布和质量、被捕食风险以及人为活动干扰等因素的影响,当动物处于不利环境时,会通过调整采食策略来满足自身的需求[9]。对南非隐鼠(Cryptomyshottentotus)在不同环境下的研究表明,其在食物资源贫乏的干旱地区的食物可获得性远远低于湿度适宜且食物资源充足地区,南非隐鼠通过拓宽其食物生态位宽度来弥补食物的不足[10]。研究发现人为活动的干扰会引起动物惊吓逃窜或提高自身警惕水平,进一步影响到动物的觅食行为,从而使影响其食性[11]。准确获取动物在人为活动干扰前后的食性及其营养生态位特征,对于认识动物与环境的相互关系有着重要意义。
喜马拉雅旱獭(Marmotahimalayana)是甘南高寒草甸重要的野生草食动物之一,其体型较大,也较为常见。通常于5—10月活动,主要以草本植物地上部分为食,营家族式生活,通常数个家族形成一个群体[12]。研究发现,当受到家畜或牧民干扰时,喜马拉雅旱獭会减少觅食活动而提高警惕水平,而家畜和牧民离开后,其大多都恢复正常活动,但仍存在持续的警戒行为。喜马拉雅旱獭以牺牲觅食时间为代价,提高警惕的行为可能会对满足冬季冬眠的能量预算产生潜在的后果[13]。旅游作为甘南草原一项特色产业,近年来有了极大的发展。频繁的人为活动对栖息于草原的喜马拉雅旱獭带来了更大的生存挑战,了解喜马拉雅旱獭在人为活动干扰后的食性及营养生态位特征,能为其有效管理提供重要参考。本研究利用粪便显微组织分析法,分析人为活动干扰后喜马拉雅旱獭的食性及其营养生态位变化,进一步探讨喜马拉雅旱獭应对人为活动干扰的食性响应特征,为喜马拉雅旱獭的管理提供参考。
1 材料与方法
1.1 试验区概况
研究区位于甘肃省甘南藏族自治州碌曲县(34°14′07″~34°48′48″ N,102°10′80″~102°58′15″ E),海拔3 550 m,年平均气温2.3℃,年降水量为632~781 mm,降水主要集中于5—9月,年蒸发量1 000~1 500 mm,年均日照时间长2 200~2 400 h。土壤以亚高山草甸土和高山草甸土为主。植被类型为高寒灌丛草甸和高寒草甸[14]。喜马拉雅旱獭是当地常见的野生动物,主要分布在碌曲县西南部[15]。
1.2 试验方法
1.2.1样地设置 在研究区域内选择远离人为活动的自然生境作为对照,距离观景台600~950 m,距离公路1 000~1 500 m,面积约为7 hm2。另选两块有人为活动的草地,三块样地植被状况一致。所选干扰生境1位于213国道两侧,距离国道3~50 m,面积约为5.8 hm2,其干扰主要为来往车辆;干扰生境2的样地为郭莽湿地观景台走廊两侧草地,距离走廊5~80 m面积约为6.2 hm2。观景台走廊位于草原中央,是一个开放式免费景点,夏季前往观光游客络绎不绝,喜马拉雅旱獭栖息于走廊两侧(图1)。

图1 研究区域及样地分布示意图Fig.1 Distribution diagram of study area and plot
1.2.2样品采集 于2018年8月,在研究区域内收集喜马拉雅旱獭新鲜粪便,根据其不同便厕、不同家群收集,不同家群里面收集不同大小、形状的粪便,以确保所收集尽可能多个体的粪便。自然生境下共采集25份喜马拉雅旱獭粪便;干扰生境1共收集到21份;干扰生境2共收集到20份。对采集研究区内出现的每种植物,鉴定植物种属并记录,带回实验室用于制作显微鉴定的植物对照样本。
1.2.3植物资源的调查 在每个样地下随机设置5个0.5 m×0.5 m的样方,样方等份为100个小格,用刺针垂直刺每个小格中央,接触到的植物记录其名称,针刺100次,统计每种植物出现的次数,即每种植物出现的累计次数占针刺总次数的百分比为该物种植物的相对比例[16]。
1.2.4显微装片的制备和镜检 将所采集的每种植物,分根、茎和叶烘干后分别进行粉碎、研磨,研磨后依次经60目和100目网筛筛选,留取100目筛上物作为试验样品。参考相关文献[17]制作显微鉴定的对照样本,在100倍显微镜下观察并拍照保存(图2)。

图2 植物显微照片(部分)Fig.2 Plant photomicrograph (part)
参考相关文献制作喜马拉雅旱獭粪便的显微装片,每份样品制作5张装片[18]。参照沈广爽[19]镜检方法:每张装片在100 倍的显微镜下检查20个视野(图3),对照植物显微装片,根据视野中不同的细胞形态类别和特点,将植物可辨认角质碎片鉴定到属种,并记录其在视野中的出现频次。
图3 粪便显微照片Fig.3 Fecal micrograph
1.3 数据分析与统计
参考相关文献[20],将统计所得的各植物角质碎片的出现频次(F),依据公式:
D=ln(1-F/100)
将其转换为每个视野中各植物可辨认碎片的平均密度(D)。
再根据公式:
相对密度(RD)=各植物可辨认碎片平均密度(D)/各植物可辨认碎片平均密度之和×100%,将D转换为RD,RD即为该种植物占食物组成的百分比,并根据RD值排列各物种可食植物的取食序位。
通过Shannon-Weiner多样性指数(H)、最大Shannon-Weiner多样性指数(Hmax)、Pielou均匀度指数(J)和食物生态位宽度指数(B)分析喜马拉雅旱獭营养生态位特征[21]。
式中:Pi为食物i在取食者食物中出现的频率,S代表粪样中存在的植物物种数。
将喜马拉雅旱獭在3种生境下的食物按功能群划分,采用SPSS 23软件进行单因素方差分析,所有数据处理在Excel 2016中完成。
2 结果与分析
2.1 植物资源分布及其相对比例
3块样地的主要植物种类及其相对比例的调查结果显示(表1)。自然生境下共调查到有17科31属33种植物,按科统计,蔷薇科(Rosaceae)、禾本科(Poaceae)和菊科(Asteraceae)植物所占的相对比例最高,分别占22.59%,17.04%和15.08%。按种统计,鹅绒委陵菜(Potentillaanserina)、垂穗披碱草(Elymusnutans)和草地早熟禾(Poapratensis)所占的相对比例最高,分别占19.56%,11.17%和5.87%。干扰生境1共调查到18科35属39种植物,按科统计菊科、蔷薇科和禾本科植物所占的相对比例最高,分别占25.40%,14.95%和14.03%。按种统计,鹅绒委陵菜、垂穗披碱草和矮莎草(Cyperuspygmaeus)所占的相对比例最高,分别占13.13%,9.22%和8.1%。干扰生境2共调查到19科35属37种植物,按科统计,菊科、唇形科(Lamiaceae)、蔷薇科和禾本科植物所占的相对比例最高,分别占31.22%,19.21%,13.62%和12.66%。按种统计,细叶亚菊(Ajaniatenuifolia)、密花香薷(Elsholtziadensa)、鹅绒委陵菜和垂穗披碱草所占的相对比例最高,分别占19.50%,18.20%,13.40%和8.88%。
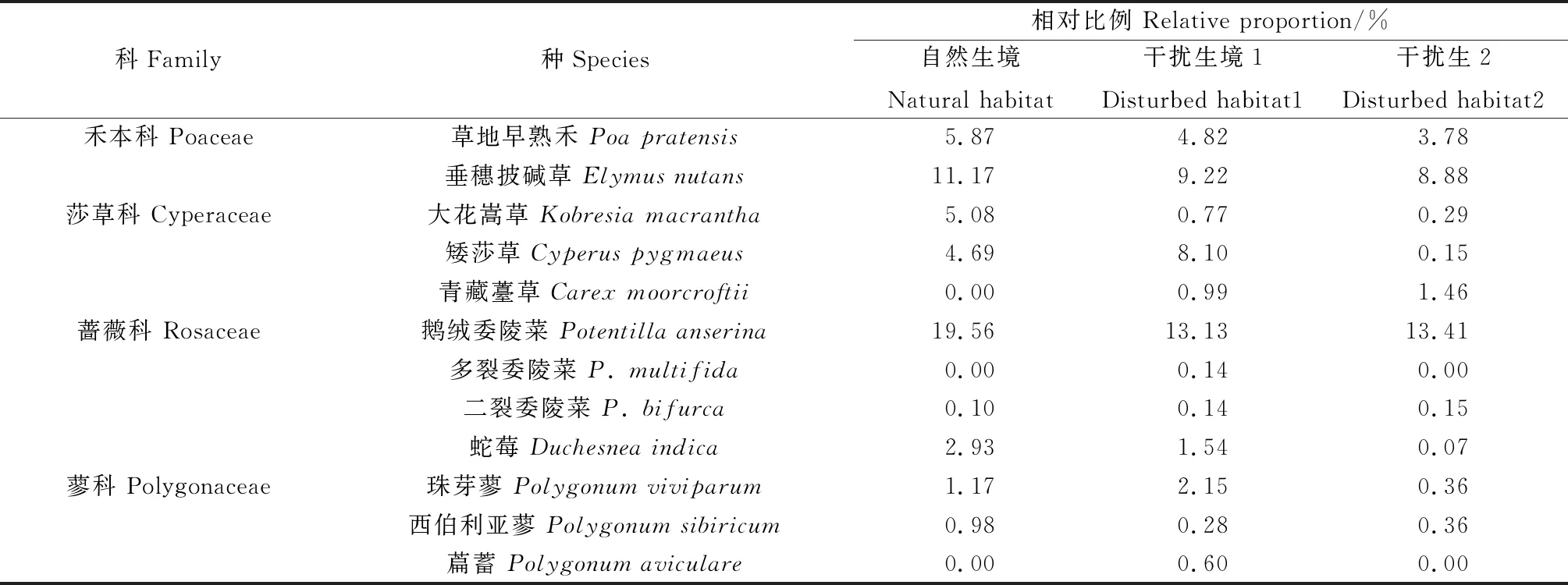
表1 不同样地的主要植物种类及相对比例Table 1 Main plant species and their relative proportions in different study areas

续表1
2.2 食物组成及比例
从收集的喜马拉雅旱獭粪便的显微组织观察结果发现,喜马拉雅旱獭在3种生境下采食的植物种类大致相同,但其比例各异。在自然生境下,喜马拉雅旱獭共采食14科28属28种植物,其中垂穗披碱草、鹅绒委陵菜和草地早熟禾是其食谱的前3种植物,分别占食谱的46.67%,30.17%和6.66%。按科统计,禾本科、蔷薇科植物的相对比例最高,分别占食谱的53.33%,30.17%。在干扰生境1中共采食14科27属28种植物,其中采食序位前3的植物为垂穗披碱草、兰石草(Lanceatibetica)和白苞筋骨草(Ajugalupulina),分别占食谱的24.59%,16.02%和14.21%。按科统计,禾本科、菊科、玄参科(Mazaceae)和唇形科植物的相对比例最高,分别占食谱的25.59%,17.16%,16.02%和14.71%。在干扰生境2中采食14科24属25种植物,其中采食序位前三的植物为垂穗披碱草、伞花繁缕(Stellariaumbellata)和箭叶橐吾(Ligulariasagitta)分别占食谱的19.00%,11.94%和11.84%。按科统计,菊科、禾本科、藜科(Chenopodiaceae)和唇形科植物的相对比例最高,分别占食谱的31.34%,19.32%,13%和11.24%(表2)。按植物功能群统计,喜马拉雅旱獭主要以杂类草和禾本科植物为食,相对于自然生境,喜马拉雅旱獭在干扰生境1和干扰生境2对莎草科和禾本科植物的采食比例显著减少(P<0.05);而对杂类草和豆科的采食比例显著增加(P<0.05);干扰生境2下对豆科的采食量显著高于自然生境(P<0.05,图4)。
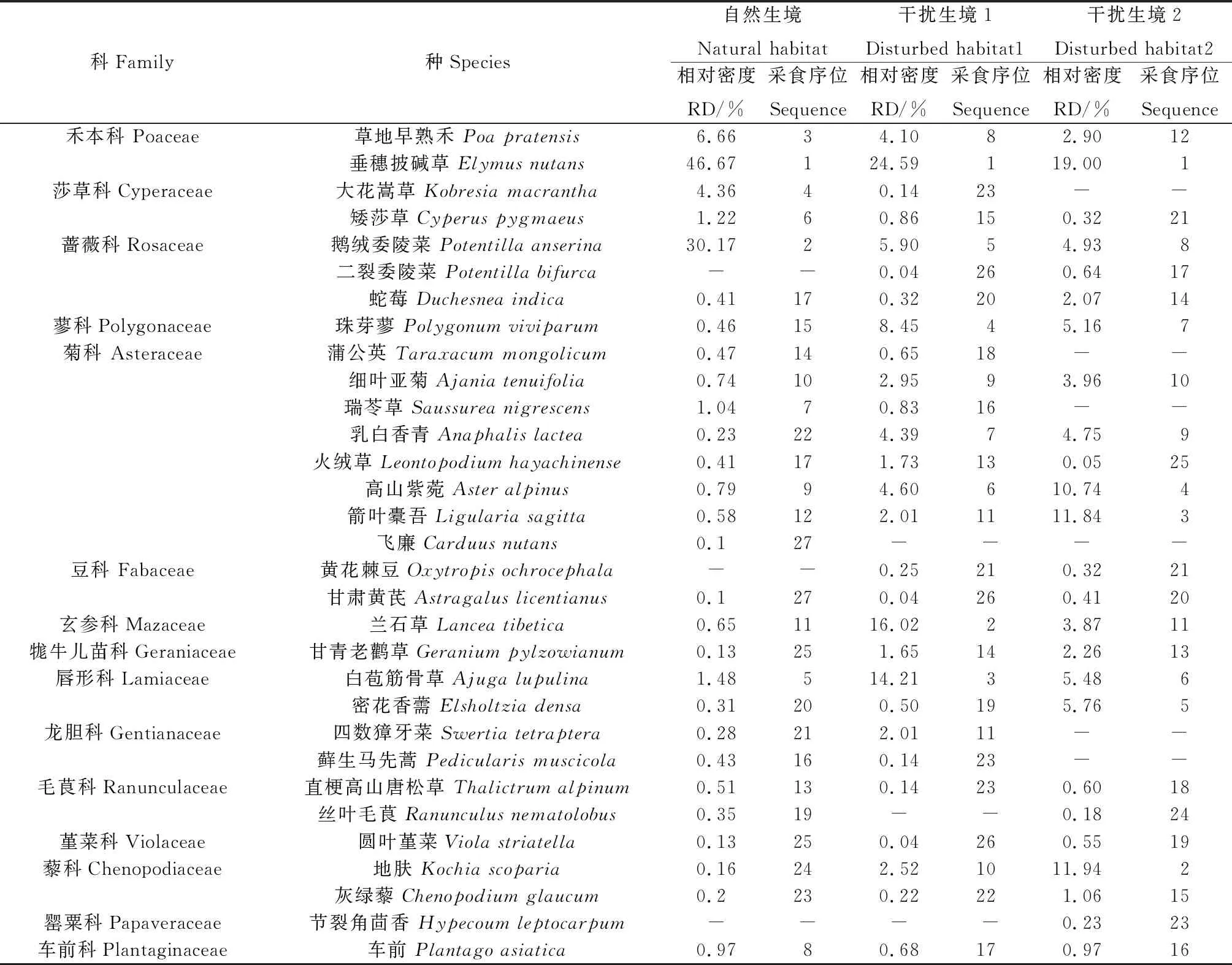
表2 喜马拉雅旱獭的食物组成及比例Table 2 Food composition and ratio of Himalayan marmot

图4 喜马拉雅旱獭食物主要类群的相对密度Fig.4 The relative density of the main food groups of the Himalayan marmot注:不同小写字母表示同一功能群植物在不同生境间差异显著(P<0.05)Note:Different lowercase letters indicate significant difference between plants of the same functional group in different habitats at the 0.05 level
2.3 营养生态位特征
喜马拉雅旱獭在自然生境和干扰生境1所采食的物种数相同,在干扰生境2所采食的物种数略少于其他生境。而干扰生境2下食物的Shannon-Weiner多样性指数H、Pielou均匀度指数J和食物生态位宽度B的值最大,干扰生境1次之,而自然生境下的各指标值最小(表3)。

表3 喜马拉雅旱獭的营养生态位参数Table 3 Nutritional niche parameters of Himalayan marmot
3 讨论
3.1 喜马拉雅旱獭的食性
本研究发现喜马拉雅旱獭食性较广,以杂类草和禾本科植物为主要的食物来源,且喜食禾本科的垂穗披碱草。这与热娜古丽等在阿尔金山对喜马拉雅旱獭食性的研究结果相似[22]。
研究区域的植被组成以杂类草和禾本科植物为主,可见栖息地食物的可利用性决定了喜马拉雅旱獭的食物选择模式。喜马拉雅旱獭在自然生境下采食28种植物,在人为活动干扰生境下分别采食28和25种植物,不同生境下调查发现的植物种类大多都出现喜马拉雅旱獭食谱中,表明喜马拉雅旱獭食性泛化。高寒草甸生活的喜马拉雅旱獭于5月下旬出蛰,9月下旬入蛰,且体内需储存大量脂肪以备冬眠,因此需要采食较多的食物来满足自身所需[14]。食性泛化有利于减少动物觅食时间,提高进食效率,进而增加动物的生存概率[23]。
3.2 人为活动干扰对喜马拉雅旱獭食性的影响
本研究发现,人为活动干扰对喜马拉雅旱獭的食性有一定的影响,尤其对不同功能群植物的影响显著。相对自然生境,喜马拉雅旱獭在干扰生境1和干扰生境2对莎草科和禾本科植物采食的相对比例显著减少(P<0.05);而对杂类草和豆科采食的相对比例显著增加(P<0.05);干扰生境2下对豆科采食的相对比例显著高于自然生境(P<0.05)。受食物资源可利用性限制,野生动物在挑选食物时,不仅受植物资源的分布和适口性的影响,还要考虑捕食者的威胁[24]。研究发现,当放牧地牧草资源丰富时,放牧家畜能够选择喜食的牧草,反之,家畜就被迫采食先前不喜食的植物[25]。3种生境下其食谱中垂穗披碱草所占的比例较大,可见喜马拉雅旱獭喜食禾本科植物。人为活动干扰对喜马拉雅旱獭生境的植被组成产生了一定的影响,特别在人为活动干扰生境2中,由于游客的踩踏等干扰,其植物优势种变为细叶亚菊和密花香薷,其喜食植物垂穗披碱草等植物比例降低。当受到人为活动干扰时,喜马拉雅旱獭受到惊吓后驻足警戒或逃窜[9],其采食的范围缩小,为满足日常的营养需求,提高了对环境中其他可利用食物的采食率,具体表现为两种不同干扰生境下其食物的Shannon-Weiner多样性指数和Pielou均匀度指数均有所增加。此外,受觅食范围的限制,其采食的植物种类也减少。
3.3 人为活动干扰对喜马拉雅旱獭营养生态位的影响
本研究中,喜马拉雅旱獭在两种人为活动干扰生境下的食物生态位宽度均大于自然生境。生态位宽度是一个种群在一个群落中所利用的各种不同资源的总和[26]。当食物资源充足时,食草动物利用最适于其生存和最方便利用的食物资源,导致营养生态位变窄,当食物资源短缺时,它们则尽量发挥其资源利用潜力,促使生态位变宽[27]。相比于夏季,赤腹松鼠(Callosciuruserythraeus)在秋季的生态位宽度会变窄,因为秋季是大多数果实成熟的季节,赤腹松鼠选择容易获得且能量高的食物以减少能量消耗[28]。有研究发现,喜马拉雅旱獭种群随着人为活动的增强,其生境内洞穴密度变大,各洞穴之间的距离变短,这样就有利于躲避人为活动潜在的捕食风险[29],但会消耗一定的能量和时间,亦有研究表明当受到人为活动干扰时,喜马拉雅旱獭提高警惕水平,减少觅食时间来应对外界干扰[11],而觅食和休息的时间对于冬眠动物越冬存活率至关重要。经典觅食理论认为,动物会通过利用资源获得高效长期的能量增益率,从而达到最大化适应性[30]。为保证日常所需的能量供应,喜马拉雅旱獭通过拓宽自身的营养生态位宽度,提高对食物资源的利用率来适应人为活动干扰对其产生的不利影响。
4 结论
人为活动干扰对喜马拉雅旱獭所采食的植物种类影响较小,其食物均以禾本科和杂类草植物为主,垂穗披碱草是其最喜食的植物。但人为活动干扰影响了喜马拉雅旱獭的食物组成,干扰生境下其采食莎草科和禾本科植物的相对比例减少,采食杂类草和豆科植物的相对比例增加,人为活动干扰增加了喜马拉雅旱獭的食物多样性及食物生态位宽度。拓宽自身食物生态位宽度是喜马拉雅旱獭应对人为活动干扰的重要策略。
猜你喜欢
小哥白尼(野生动物)(2022年8期)2022-09-20 03:48:22
小哥白尼·野生动物画报(2022年8期)2022-05-30 10:48:04
当代水产(2021年10期)2022-01-12 06:21:02
小哥白尼(野生动物)(2021年10期)2021-02-12 03:10:10
上海包装(2019年2期)2019-05-20 09:10:56
广东石油化工学院学报(2016年3期)2016-05-17 05:16:24
湖南城市学院学报(自然科学版)(2016年4期)2016-02-27 14:02:41
中国粮油学报(2016年5期)2016-01-23 02:44:49
农产品市场周刊(2015年39期)2015-10-31 19:12:27
草食家畜(2012年2期)2012-03-20 13:22:34
