
青藏高原高寒草甸六种杂类草植物种子萌发期抗旱性研究
2022-06-02 02:02:02张春辉任彦梅姚步青黄小涛周华坤
草地学报 2022年5期
张春辉,马 真,任彦梅,武 林,姚步青,黄小涛,马 丽,郭 婧,周华坤
(1.青海大学省部共建三江源生态与高原农牧业国家重点实验室,青海 西宁 810016; 2. 中国科学院西北高原生物研究所青海省寒区恢复生态学重点实验室,青海 西宁 810008; 3. 中国科学院高原生物适应与进化重点实验室,青海 西宁 810008;4. 青海省社会科学院,生态环境研究所,青海 西宁 810000)
种子的产生代表着一段生活史的结束,也预示着新的生活史的开始。对于很多植物来说,有机会萌发的种子只是自身产生种子中很少的一部分,而这仅有的一部分种子在萌发过程中会受到各种环境因素的影响(光照、水分、温度等)[1-3]。种子在面对这些不确定环境因子时的萌发策略,决定着它后续的成功与否。
干旱是植物整个生活史中都必须要面对的一个重要问题[4-5],而种子萌发阶段作为生活史的开始,对干旱的响应尤其敏感而关键。该阶段的抗旱性直接影响着物种的建植、种群的扩张,以及群落结构。研究认为,植物种子的萌发率通常会随着干旱胁迫程度的加剧而降低[6-7]。聚乙二醇(Polyethylene glycol,PEG)是一种亲水性物质,因无法透过细胞壁而对种子没有伤害,通常被用来模拟自然干旱胁迫从而研究植物萌发期抗旱性[8-9]。
种子大小是植物的关键性状之一,也是它的生活史中第一个起决定性作用的性状。不同的种子大小代表了种子内部储存营养物质的含量,而种子储存营养物质的含量决定了它在逆境条件下的耐受力[10]。对植物种群内不同大小种子的萌发研究中发现,逆境条件下,大种子比小种子具有更强的耐受性[11]。在对干旱胁迫下文冠果种子大小和萌发关系的研究中发现,高浓度PEG胁迫下,大种子萌发率比小种子高[12]。对干旱胁迫下的侧柏种子萌发研究中同样发现,大种子比小种子更具有优势[13]。种子大小和萌发干旱耐受性关系的研究多基于物种种内的种子大小变异,而在不同物种间种子大小和萌发干旱耐受性的关系研究较少。研究物种间种子大小和萌发干旱耐受性的关系对于抗旱物种的筛选具有重要的意义。种子相关的其它性状和干旱耐受性关系的相关研究鲜有报道。
高寒草甸广泛分布于青藏高原,是青藏高原最为主要的植被类型[14]。青藏高原高寒草甸生态系统有着较高的生物多样性[15]。但在土地利用和气候变化的选择压力下,高寒草甸面临着前所未有的挑战[16]。种子资源是退化草地恢复的重要保障[17]。对干旱胁迫下种子抗逆性的研究,可以为退化草地恢复的种质资源利用和保护提供理论依据。
杂类草植物作为高寒草甸植物群落中不可或缺的一部分,是高寒草甸主要的功能群之一[18]。杂类草植物分布较广,且种子产量较高,是构建人工群落时物种收集和选取的优良材料。由此我们选择了高寒草甸常见的六种杂类草植物种子,对其不同干旱胁迫强度下的抗旱性进行了研究,以期明晰干旱胁迫下种子相关性状和干旱耐受性的关系,为退化草地恢复中抗旱物种的筛选提供理论支持。
1 材料与方法
1.1 研究区域概况
试验所需植物种子采集于青藏高原东北部(37°29′~37°45′ N,101°12′~101°23′ E,海拔3 200 m)的青海省海北州门源县境内的中国科学院海北高寒草甸生态系统定位站。该区年降水量为561 mm,主要分布于6—9月;年均温0.8℃,属于典型的高原大陆性气候。
1.2 试验方法
1.2.1试验种子采集及前处理 试验材料于种子成熟期(8—9月)采集,每一物种选取3个以上种群,每一种群随机选取 20株以上的植株。将采集后的种子带回实验室进行去皮去杂等处理,将处理完成的种子放于野外(种子采集地),次年春季取回后进行试验。具体物种见表1。

表1 六种杂类草植物物种名及种子大小Table 1 Species names and seed sizes of six weed species
1.2.2PEG干旱胁迫试验 试验使用不同浓度的聚乙二醇溶液模拟不同程度的干旱。选择饱满、健康种子进行干旱胁迫试验,每个物种四个重复,每个重复25粒种子。根据前期研究经验,并结合当前的研究方法,选取了0%(对照),10%,20%三个PEG浓度。种子放于铺有两层滤纸的培养皿内,分别加入适量PEG溶液。萌发试验在培养箱(温度15~20℃,黑暗/光照:10 h/14 h)内进行,每3天更换一次滤纸和溶液,以维持溶液水势不变。以胚芽长度为种子长度的1/2作为萌发标准,从种子置于培养皿第二日开始每天记录发芽种子数,连续7天培养皿内没有发芽种子视为萌发结束。
1.2.3种子大小测定 随机选取饱满健康的种子进行试验,并用万分之一的电子天平称量每种植物种子的百粒重,重复3次,最后换算成单粒种。每一物种的单粒种子重量作为种子大小。
1.3 数据分析
1.3.1萌发指标及计算方法: 萌发率(Germination proportion)=(试验结束时萌发的种子总数/试验处理种子总数)×100%;
萌发干旱耐受性(Drought tolerance during seed germination)=1-(对照下的萌发率-PEG处理的萌发率)/PEG处理的萌发率。
萌发干旱耐受性的取值范围为0~1,其取值越接近1表明萌发干旱耐受性越高,其取值越接近0表明萌发干旱耐受性越低。
1.3.2数据统计与分析方法 使用R软件计算物种间系统发育距离。使用SPSS 22.0 软件进行后续数据统计与分析,用T检验(T test)检验显著性水平,应用一般线性回归检验种子大小和萌发率与萌发干旱耐受性的回归关系以及物种间系统发育距离和萌发干旱耐受性差异的回归关系。
2 结果与分析
2.1 六种杂类草植物在不同干旱胁迫强度下的萌发率
2.1.1六种杂类草植物种间萌发率 六种杂类草植物萌发率在对照条件下,10% PEG干旱胁迫下,20% PEG干旱胁迫下差异均显著(P<0.05,图1)。
对照条件下,萌发率在75%~100%的物种有三个,萌发率最高的物种为尼泊尔蝇子草(Silenenepalensis),对照条件下萌发率为97.96%;其次为天蓝韭(Alliumcyaneum),萌发率为83.50%,两者萌发率差异不显著;蓝翠雀花(Delphiniumcaeruleum)萌发率为75.56%,显著低于尼泊尔蝇子草(P<0.05)。萌发率在50%~75%的物种为蓬子菜(Galiumverum)(萌发率为68.35%)。萌发率在25%~50%的物种为青海当归(Angelicanitida)(萌发率为40.38%)。萌发率在0~25%的物种为半抱茎葶苈(Drabasubamplexicaulis)(萌发率为20.10%),显著低于其它5个物种(P<0.05,图1a)。
10% PEG干旱胁迫下,萌发率在50%~75%的物种有天蓝韭(萌发率为72.62%),尼泊尔蝇子草(萌发率为63.37%),蓝翠雀花(萌发率为59.83%)。萌发率在25%~50%的物种为蓬子菜(萌发率为40.83%)。萌发率在0~25%之间的物种为青海当归(萌发率为24.00%)和半抱茎葶苈(萌发率为3.13%),在该处理条件下半抱茎葶苈萌发率显著低于其他5个物种(P<0.05,图1b)。
20% PEG干旱胁迫下,各物种萌发率均在0~25%之间。萌发率最高的物种为尼泊尔蝇子草,萌发率为20.12%,显著高于其它5个物种(P<0.05)。其余物种萌发率均低于10%,其中天蓝韭萌发率为6.94%,蓝翠雀花萌发率为6.25%,蓬子菜萌发率1.14%,3个物种萌发率差异不显著。完全不萌发的物种有青海当归和半抱茎葶苈,萌发率均为0(图1c)。
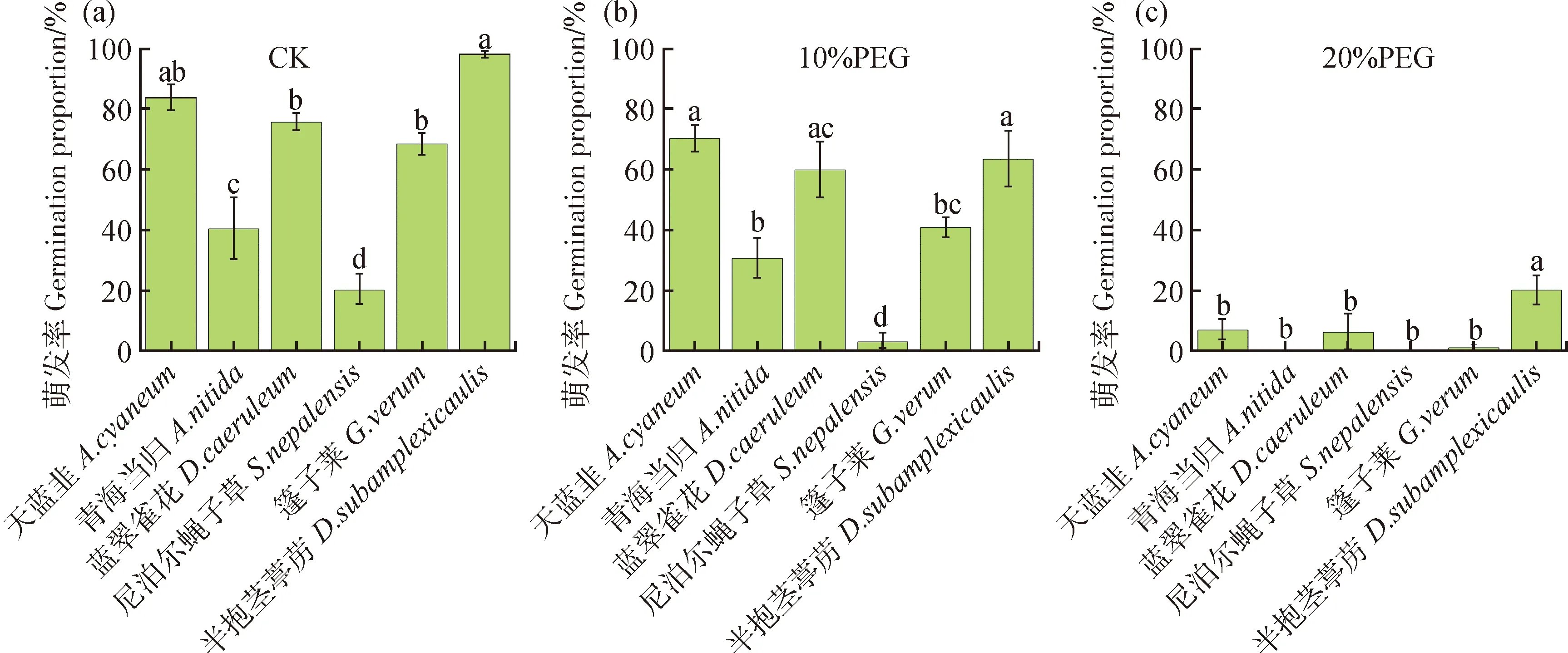
图1 六种杂类草植物种间萌发率比较Fig.1 Comparison of germination rates of six forbs species注:不同小写字母代表不同物种间萌发率差异显著(P<0.05)Note:Different lowercase letters showed significant differences among different species at the 0.05 level
2.1.2六种杂类草植物种子萌发率对PEG干旱胁迫的响应 六种杂类草植物萌发率在三种处理间均差异显著的物种为尼泊尔蝇子草和蓬子菜。尼泊尔蝇子草为对照条件下萌发率最高的物种,萌发率为97.96%;10%PEG干旱胁迫下萌发率为63.37%;20%PEG干旱胁迫下萌发率为20.12%,三种处理间差异显著(P<0.01)(图2f)。蓬子菜对照条件下萌发率为68.35%;10%PEG干旱胁迫下萌发率为40.83%;20%PEG干旱胁迫下萌发率为1.14%,三种处理间差异显著(P<0.01)(图2e)。
三种杂类草植物在10%PEG浓度下的萌发率显著低于对照。除了蓬子菜和尼泊尔蝇子草,还有半抱茎葶苈。半抱茎葶苈为对照条件下萌发率最低的物种,对照条件下萌发率为20.10%;10%PEG干旱胁迫下萌发率为3.13%;20%PEG干旱胁迫下萌发率为0,三种处理间差异显著(P<0.01)(图2d)。
所有物种在20%PEG浓度下的萌发率都显著低于对照(图2)。除半抱茎葶苈,其它5个物种在20%PEG浓度下的萌发率都显著低于10%PEG浓度(P<0.05)。

图2 六种杂类草植物种子萌发率对PEG干旱胁迫的响应Fig.2 Response of seed germination proportion of six forbs to PEG treatment注:不同小写字母代表不同PEG浓度萌发率差异显著(P<0.05)Note:Different lowercase letters showed significant differences of seed germination among PEG treatment at the 0.05 level
2.2 种子性状和萌发干旱耐受性关系研究
种子大小和萌发干旱耐受性呈现显著正相关关系(P=0.015;R2=0.809),随着种子质量的增加,种子的抗旱性增加(图3a),半抱茎葶苈种子对干旱胁迫的耐受性最低,种子质量为0.04 mg;青海当归种子对干旱胁迫的耐受性最高,种子重量为1.71 mg。
六种不同大小的杂类草植物种子初始萌发率差异显著(图1a),尼泊尔蝇子草对照条件下萌发率为97.96%,而半抱茎葶苈萌发率仅为20.10%,反映了这些物种具有不同程度的休眠。然而种子萌发率(即休眠程度)和萌发干旱耐受性没有显著回归关系(P=0.148,图3b)。

图3 种子性状和萌发干旱耐受性回归图Fig.3 Relationship between seed traits and drought tolerance during germination
2.3 系统发育和萌发干旱耐受性关系研究
结果表明物种间系统发育距离和萌发干旱耐受性没有显著回归关系(P=0.887,图4),这说明系统进化对六种物种间萌发干旱耐受性分化没有影响。

图4 物种间系统发育距离和萌发干旱耐受性差异的回归图Fig.4 Relationship between phylogenetic distance and differences of drought tolerance during seed germination between species
3 讨论
种子萌发阶段是植物生活史的重要阶段,而种子萌发阶段的抗旱性强弱是影响该阶段成功与否的重要因子。研究结果表明,不同浓度的PEG干旱胁迫对六种高寒草甸杂类草植物种子的萌发都具有一定的抑制作用。种子的萌发率随着干旱胁迫强度的增加而下降,这与相关的研究结果一致[19-20]。较低浓度PEG(10%)干旱胁迫处理下,天蓝韭、蓝翠雀花、青海当归虽然有下降趋势,但与对照差异不显著,这三个物种可能对该程度的干旱胁迫有一定的耐受性;而半抱茎葶苈、蓬子菜、尼泊尔蝇子草与对照下的萌发率差异显著,这几个物种的萌发率在较低PEG干旱胁迫下就发生了显著的变化。较高浓度PEG(20%)干旱胁迫处理下,六种杂类草植物种子萌发率都与对照差异显著。其中半抱茎葶苈种子萌发率下降最多,10%PEG干旱胁迫下降至对照处理下的1/4,20%PEG干旱胁迫下降至0,该物种对干旱胁迫最为敏感。
种子大小是植物的关键性状,决定着种子内部储藏物质以及相关的耐受性。通常来说,质量大的种子会具有更强的耐受性和存活能力[21-22],而小种子物种在数量和传播方式方面更具有优势[23]。本研究结果表明,种子萌发阶段对干旱胁迫的耐受性随着种子质量的增加而增加,大种子物种在萌发阶段具有较高的干旱耐受性,而小种子物种的萌发更容易受到干旱胁迫的影响。半抱茎葶苈种子质量为0.04 mg,对干旱胁迫的耐受性最低,青海当归种子重量为1.71 mg,对干旱胁迫的耐受性最高。郭有燕和付锦雪对种群内不同大小种子抗旱性的研究中有相似的发现[12-13]。大种子物种萌发阶段在干旱胁迫或其它逆境条件下的耐受性,部分抵消了小种子的数量优势,为逆境条件下大种子物种的建植成功提供了机会[24]。
六种不同大小的杂类草植物种子初始萌发率差异显著,尼泊尔蝇子草对照条件下萌发率为97.96%,而半抱茎葶苈萌发率仅为20.10%,反映了这些物种具有不同程度的休眠。但在不同程度的干旱胁迫下,即使这些物种具有不同程度的休眠,种子大小和萌发干旱耐受性之间仍保持着显著正相关关系。
一般而言,系统进化历史可以影响物种特性的分化[25]。然而,我们的研究中没有发现系统进化历史影响物种萌发耐旱性的分化。这可能是因为萌发耐旱性像其他萌发特性一样,有着一定的本地适应性或者环境可塑性[26],从而导致我们研究的物种间萌发耐旱性可能出现了趋同化,最终掩盖了系统进化历史的影响。
4 结论
不同浓度PEG处理对六种杂类草植物种子萌发都有不同程度的抑制作用。随着PEG浓度的增加,种子的萌发率呈降低趋势。其中较低浓度PEG(10%)胁迫处理对部分植物的种子萌发具有显著的抑制作用,而较高浓度的PEG(20%)胁迫处理对六种杂类草植物种子萌发均有显著的抑制作用。种子大小和干旱耐受性正相关,随着种子质量的增加,种子萌发阶段的抗旱性增加;而萌发率和系统发育距离与萌发干旱耐受性没有显著相关性。综上,可以优先考虑大种子物种作为退化草地恢复中抗旱物种。
猜你喜欢
园林科技(2020年2期)2020-01-18 03:28:18
小学生学习指导(低年级)(2018年9期)2018-09-26 05:59:28
读者·校园版(2018年1期)2017-12-16 18:19:11
浙江农业学报(2017年1期)2017-05-17 06:13:36
西南农业学报(2016年4期)2016-05-17 05:42:13
天津科技大学学报(2016年1期)2016-02-28 16:59:44
浙江大学学报(农业与生命科学版)(2015年4期)2015-12-15 12:47:44
少年科学(2015年2期)2015-08-12 04:13:36
山西农经(2015年7期)2015-07-10 15:00:57
中国医学科学院学报(2015年5期)2015-03-01 04:03:37
