
新疆地区番茄潜叶蛾遗传多样性的ISSR分析
2022-05-30 03:15李爱梅付开赟丁新华吐尔逊阿合买提冯宏祖郭文超
生物安全学报 2022年2期
李爱梅, 付开赟, 丁新华, 何 江, 吐尔逊·阿合买提, 冯宏祖*, 郭文超
1塔里木大学植物科学学院,新疆 阿拉尔 843300; 2新疆农业科学院植物保护研究所/农业农村部 西北荒漠绿洲作物有害生物综合治理重点实验室,新疆 乌鲁木齐 830091; 3新疆农业科学院 微生物应用研究所/新疆特殊环境微生物重点实验室,新疆 乌鲁木齐 830091
南美番茄潜叶蛾Tutaabsoluta(Meyrick),别名番茄潜叶蛾、番茄潜麦蛾、番茄麦蛾,属鳞翅目Lepidoptera麦蛾科Gelechiidae,原产于南美洲的秘鲁,是世界性的检疫入侵害虫(李栋等,2019)。该虫的传播速度快,截至2021年2月11日,已入侵105个国家(南美洲10个、北美洲3个、欧洲31个、非洲35个、亚洲26个),成为世界性番茄SolanumlycopersicumL.的重要害虫。在我国,新疆伊犁、云南临沧、贵州、四川、广西、湖南及江西等地,相继发现该虫危害(陆永跃,2021)。在云南省弥渡县,大棚里的番茄植株被害率高达100%,果实被害率达19.33%,最高可达40%(尹艳琼等,2021)。番茄潜叶蛾的寄主范围较广,可危害包括茄科、豆科、锦葵科、苋科、旋花科、藜科、菊科、十字花科以及禾本科在内的近40种植物,但主要危害茄科植物,特别嗜食番茄(包括鲜食番茄、加工番茄和樱桃番茄/圣女果)(Camposetal.,1976; Cisneros & Mujica,1998; Pereyra & Sánchez,2006)。番茄潜叶蛾幼虫潜入叶片并取食叶肉,还会钻入果实内危害,成虫喜在植株幼嫩的叶片上产卵,但随着该植株的危害程度加剧,产卵偏好有所减弱(阿米热·牙生江等,2021)。番茄潜叶蛾具有很高的生物潜能,对不同气候条件的适应性等方面存在差异(Vargas,1970),可能存在着更深层次的分化。但关于我国番茄潜叶蛾的种群分化以及遗传多样性的研究还未见报道。
近些年,随着分子生物学技术、分子标记种类及其监测技术的不断发展,ISSR(简单重复序列区间 inter simple sequence repeat)分子标记方法由于操作简单、成本较低、耗时短等,已成为物种遗传变异监测的重要方法之一(Camposetal.,2017),被广泛应用于鉴定种资资源、筛选稀有品种以及探索各种群遗传多样性等(Essadkietal.,2006; Guptaetal.,2008; Reddyetal.,2002)。
番茄潜叶蛾是农业上的重要害虫,该虫的入侵直接导致在一个生长季节内番茄地块杀虫剂的使用次数由12次突增到30次以上(张桂芬等,2018; Gontijoetal.,2013),杀虫剂的过度使用,导致该虫对多种杀虫剂产生了抗药性。研究发现,杀虫剂的使用可引起昆虫遗传多样性及遗传分化(褚栋等,2008)。新疆地区各地理种群的番茄潜叶蛾对不同类型的杀虫剂产生了抗药性,因此,对新疆地区不同地理种群的番茄潜叶蛾进行遗传多样性的研究有利于了解不同区域的番茄潜叶蛾基因及遗传分化情况对不同地理种群的影响。然而,迄今为止有关新疆地区番茄潜叶蛾的基因流及遗传分化情况的研究较少。本研究拟采用ISSR分子标记技术对所采集到的20个不同地理种群的番茄潜叶蛾进行遗传多样性分析,旨在明确不同地理环境下番茄潜叶蛾对其适应性引起的遗传变化,为番茄潜叶蛾的综合治理提供参考。
1 材料与方法
1.1 实验材料
番茄潜叶蛾样品采自新疆喀什地区喀什市、伽师县、疏附县、巴楚县、疏勒县、叶城县、莎车县,伊犁特克斯县、霍城县、霍尔果斯市、伊宁市、新源县、察布查尔县,阿克苏温宿县,阿拉尔市,阿克苏市等地区及贵州省清镇市,云南省昆明市、玉溪市,哈萨克斯坦阿拉木图州等地,样品采集后立即放入75%酒精中,-20 ℃保存。记录20个地理种群的编号、采样时间、地理位置、经纬度、海拔高度及样本数量大小。
1.2 DNA的提取
对采集的番茄潜叶蛾进行基因组DNA提取,提取方法按TaKaRa裂解液说明书进行操作。用浓度为2%的琼脂糖凝胶进行电泳检测,使用B-500 BIOPHOTO METER(METASH)测定其浓度及纯度,选取D260 nm/D280 nm值在1.7~1.9、浓度大于30 ng·mL-1的DNA样品进行ISSR遗传多样性分析。
1.3 ISSR预实验分析
1.3.1 引物筛选 根据预实验,从哥伦比亚大学公布的第9套100条ISSR引物序列中共筛选出15条条带清晰、多态性好的引物进行ISSR-PCR扩增反应实验。
1.3.2 ISSR-PCR扩增 PCR反应体系(25 μL):2×Easy Taq PCR Super Mix 12.5μL、引物2 μL、模板DNA 1 μL、ddH2O 9.5 μL。PCR扩增条件:94 ℃预变性3 min;94 ℃变性30 s,33~55 ℃(不同引物的退火温度不一致)退火30 s,72 ℃延伸1 min,30个循环;72 ℃延伸5 min;4 ℃下保存。
1.4 数据统计与分析
电泳图谱中的每一条带均视为一个分子标记,代表一个等位基因位点。按凝胶同一位置上DNA条带的有无进行统计,有条带记为“1”,无条带记为“0”,将所有个体在全部位点上构成1个(0或1)矩阵模型。在得到ISSR 原始数据矩阵后,将矩阵导入软件中,利用POPGENE 32软件计算各种群的遗传参数,计算各群体的有效等位基因数Ne、Nei′s基因多样性指数H、Shannon信息多样性指数I、多态位点比率、群体内基因多样度Hs、总基因多样度Ht、2种群或亚种群间的遗传分化系数Gst(Gst=Ht-Hs/Ht),以及基因流Nm=(1-Gst)/Gst等参数(张桂芬等,2020)。根据Nei′s遗传距离的分析结果,利用NTSYS 2.1软件对其进行UPGMA聚类分析,根据所读取的数据分析相应的结果。
2 结果与分析
2.1 番茄潜叶蛾种群DNA的ISSR扩增结果
根据预实验结果,得出各引物的最佳退火温度(表1),再用各引物对不同地理种群进行ISSR扩增(图1)。利用15条ISSR引物对20个种群297头番茄潜叶蛾的DNA进行扩增,共得到137条可统计的条带,平均每条引物扩增了9.133条条带,其中引物857、885扩增的条带数最高,为13条,引物818扩增出的条带数最少,为6条;各引物扩增出的多态性条带及其占比存在差异,多态性条带占比最大的是引物807、808、811、812、826、827、856、857、868、872、886,为100%。最小的为引物885,为84.62%。不同种群的每个个体都呈现出唯一的ISSR基因型,说明ISSR分子标记对番茄潜叶蛾有较高的鉴别效率,也说明番茄潜叶蛾的个体间存在较大的遗传变异。这些位点在各个地理种群中分布不平衡,从番茄潜叶蛾20个种群的多态位点占比中可看出,番茄潜叶蛾的遗传多样性较丰富。
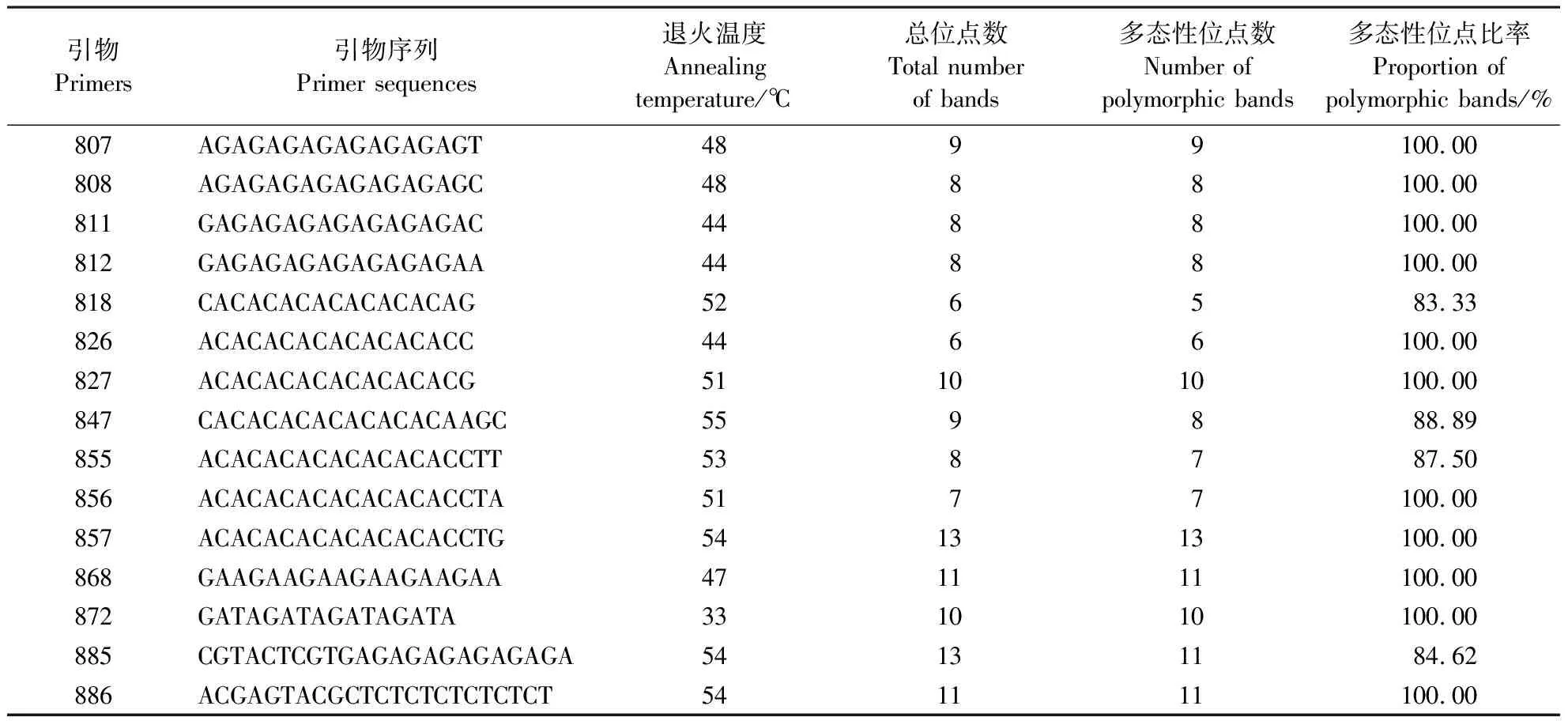
表1 ISSR扩增中引物退火温度及多态性条带Table 1 Primer annealing temperature and polymorphic bands in ISSR amplification

图1 引物827对察布查尔县种群DNA的ISSR扩增图Fig.1 ISSR amplification map of primer 827 on the DNA of Qapqal County population M: DL2000 plus DNA marker;1~24:均为察布查尔县样品。 M: DL2000 plus DNA marker; 1-24: Samples from Qapqal County.
2.2 番茄潜叶蛾种群的遗传多样性分析
从表2可知,20个番茄潜叶蛾种群的多态性位点比率在31.30%~93.13%,平均多态性位点比率为58.28%。察布查尔县(CBCE)地理种群多态性位点占比最高(93.13%),多态性条带多达13条,特克斯县(TKS)地理种群多态位点占比最低(31.30%)。从地理空间分布上看,中国南方地区部分番茄潜叶蛾种群(GZ、YNYX、YNKM)的多态性位点占比低于北方地区种群(KS、JS、SF、CBCE)。特克斯县(TKS)的Nei′s基因多样性指数(0.1153)最小,察布查尔县(CBCE)的Nei′s基因多样性指数(0.2531)最大,说明中国南北方的各个地理种群Nei′s基因多样性指数并无规律。特克斯县(TKS)的番茄潜叶蛾种群的I(0.1719)最小,察布查尔县(CBCE)种群的I(0.3976)最大,说明南北地区的番茄潜叶蛾种群遗传多样性较高。
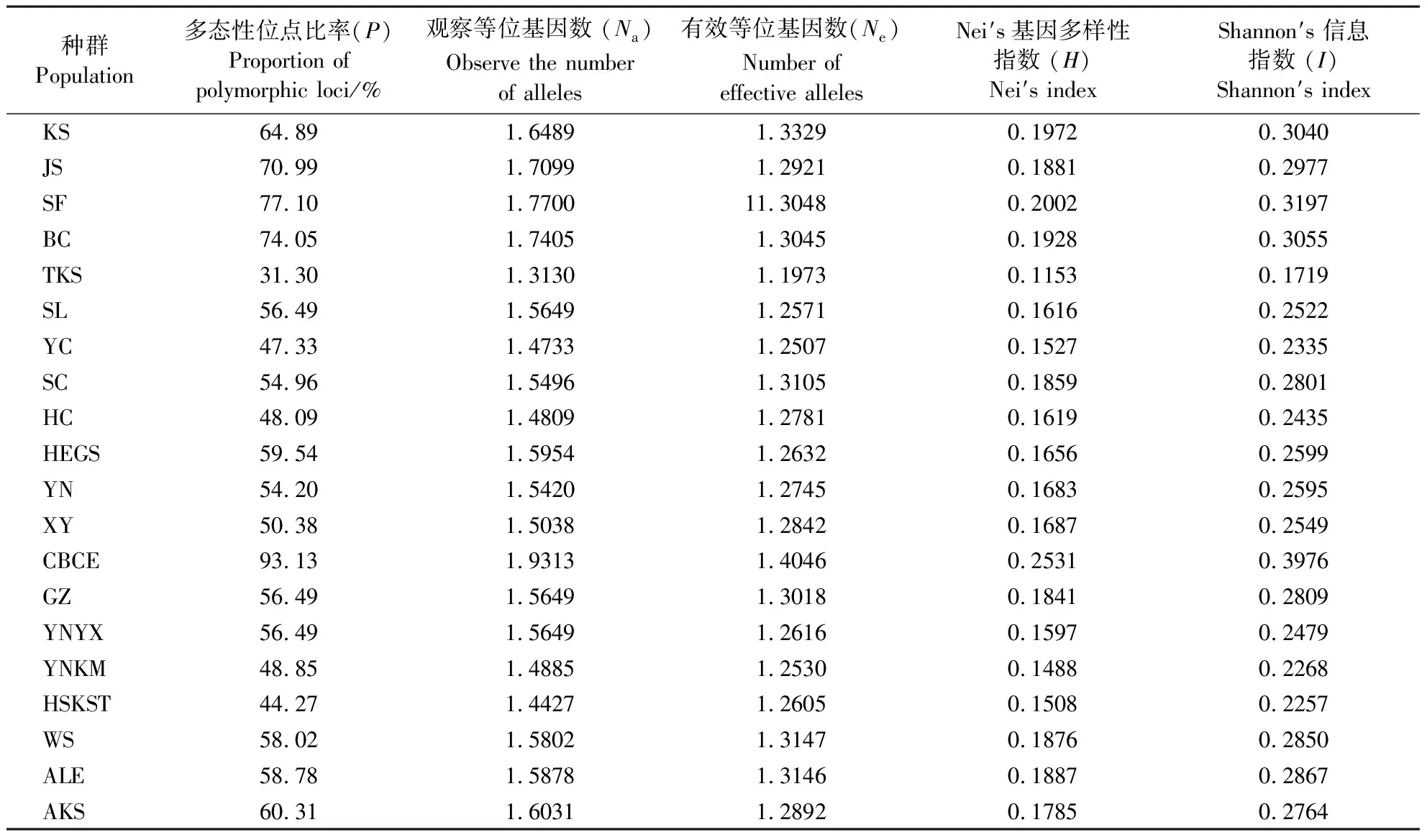
表2 番茄潜叶蛾种群间的遗传多样性指数Table 2 Genetic diversity index among T. absoluta populations
2.3 番茄潜叶蛾种群遗传分化及基因流
对番茄潜叶蛾种群内遗传分化进行分析,物种水平上的总基因多样性(Ht)为0.2351,种群内的遗传分化系数(Hs)为0.1755。根据Nei′s基因多样度算法,20个番茄潜叶蛾地理种群之间的的遗传分化系数(Gst)为0.2536,说明变异来自不同的种群之间及种群内部的个体间,占总变异的25.36%,有74.64%的变异来自于种群内的个体之间。本研究检测出番茄潜叶蛾物种水平上的基因流(Nm)为1.4716,表明番茄潜叶蛾各种群间的基因流存在一定的流动性。
2.4 遗传一致度、遗传距离及聚类分析
由表3可知,番茄潜叶蛾20个地理种群的遗传一致性系数在0.8502~0.9935,其平均遗传一致性系数为0.9329,察布查尔县(CBCE)种群与特克斯县(TKS)种群的遗传一致性系数(0.8502)最小,疏附县(SF)种群与伽师县(JS)种群的遗传一致性系数(0.9935)最大。通过对遗传一致性的分析,说明所采集的各个地理种群间的遗传一致性较高。
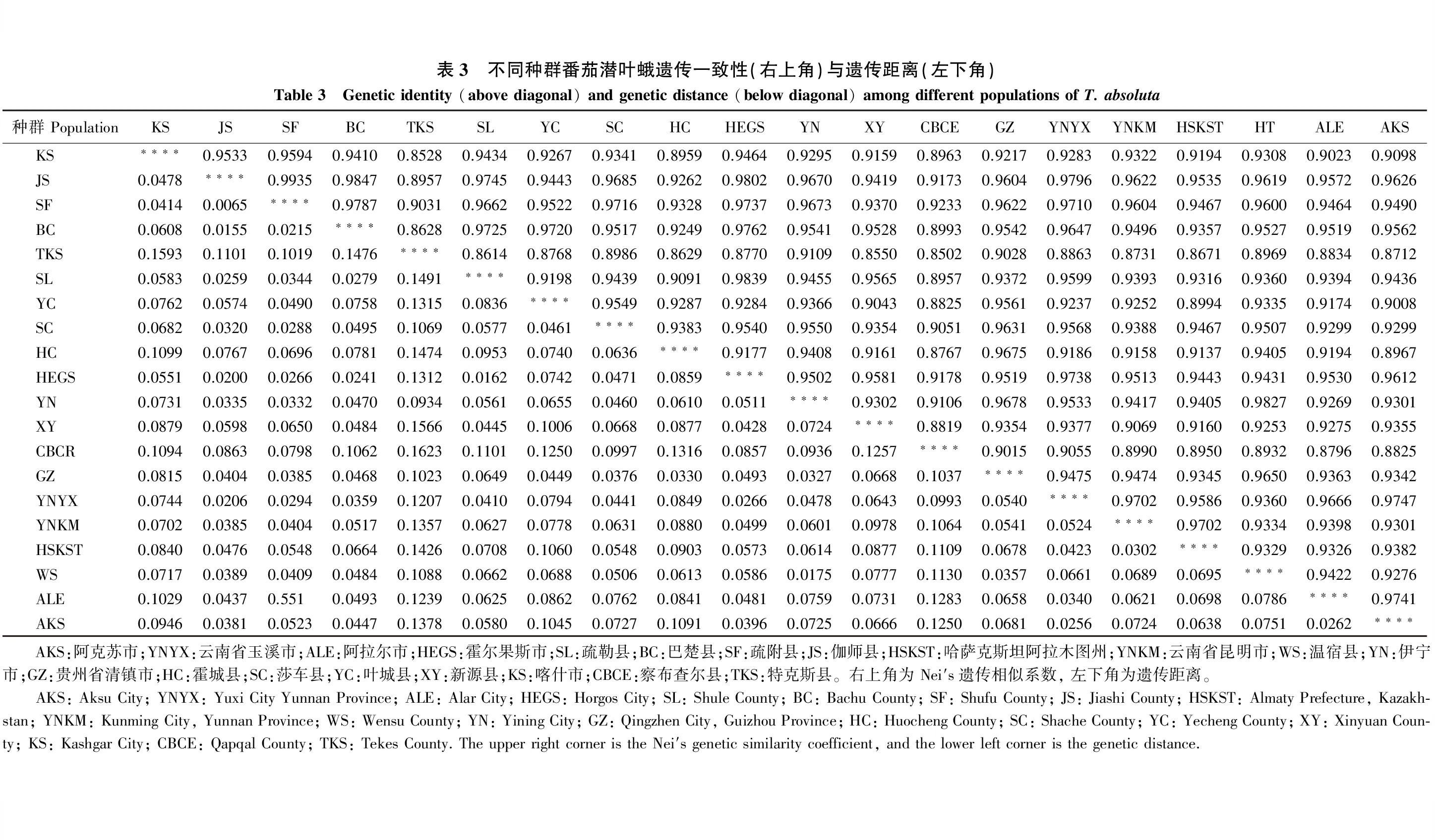
遗传距离的大小反映了每个地理种群间彼此亲缘关系的远近,为确定番茄潜叶蛾20个地理种群间的遗传关系,利用Nei′s无偏估计遗传距离进行了计算(表 3)。20个番茄潜叶蛾种群的遗传距离大小在0.0065~0.1623,其平均遗传距离为0.0726,疏附县(SF)种群与伽师县(JS)种群间的遗传距离最小,为0.0065,察布查尔县(CBCE)种群与特克斯县(TKS)种群的遗传距离最大,为0.1623。表明番茄潜叶蛾不同地理种群之间,遗传分化存在着一定程度上的差异,但这种差异比较小。
使用UPGMA方法对各个番茄潜叶蛾地理种群进行聚类分析,结果如图2所示,当遗传一致性系数为0.94时,20个地理种群的聚类分为5组,最先分出的是特克斯县(TKS)、察布查尔县(CBCE)、喀什市(KS)、新源县(XY),其余各地理为一组。其中:云南玉溪(YNYX)种群夹杂在新疆阿克苏地区的地理种群中;哈萨克斯坦阿拉木图州、云南、贵州、伊犁地区与喀什地区的部分地理种群交叉在一个分支里。在本研究中,番茄潜叶蛾的遗传一致性较高,并未形成明显的遗传结构,同时各个地理种群间与地理空间并未体现出一致的关系。
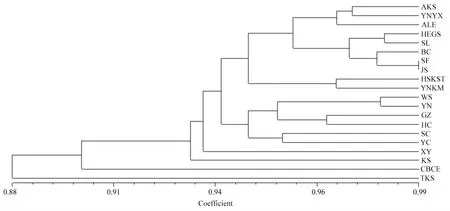
图2 20个番茄潜叶蛾地理种群间的聚类分析图Fig.2 Cluster analysis of 20 geographical populations of T. absoluta KS:阿克苏市;YNYX:云南省玉溪市;ALE:阿拉尔市;HEGS:霍尔果斯市;SL:疏勒县;BC:巴楚县;SF:疏附县;JS:伽师县;HSKST: 哈萨克斯坦阿拉木图州;YNKM:云南省昆明市;WS:温宿县;YN:伊宁市;GZ:贵州省清镇市;HC:霍城县;SC:莎车县; YC:叶城县;XY:新源县;KS:喀什市;CBCE:察布查尔县;TKS:特克斯县。 AKS: Aksu City; YNYX: Yuxi City, Yunnan Province; ALE: Alar City; HEGS: Horgos City; SL: Shule County; BC: Bachu County; SF: Shufu County; JS: Jiashi County; HSKST: Almaty Prefecture, Kazakhstan; YNKM: Kunming City, Yunnan Province; WS: Wensu County; YN: Yining City; GZ: Qingzhen City, Guizhou Province; HC: Huocheng County; SC: Shache County; YC: Yecheng County; XY: Xinyuan County; KS: Kashgar City; CBCE: Qapqal County; TKS: Tekes County.
3 讨论与结论
一个物种目前现有的遗传多样性及遗传结构是根据自身原因与所在的环境因素共同作用的结果,如地理分布、基因漂变、基因流、物种进化的历史及交配系统等。就番茄潜叶蛾而言,该虫于2006年入侵欧洲,目前已入侵世界上105个国家。该虫繁殖力强、世代重叠严重、适应性强,可借助人类活动进行远扩散,这些特征有助于番茄潜叶蛾各种群间的基因交流,减小了遗传漂变的影响。
本研究所选取的番茄潜叶蛾种群来自于哈萨克斯坦阿拉木图州、云南、贵州以及新疆喀什、阿克苏、伊犁地区,其各个不同地理种群间的遗传多样性丰富及遗传结构变异不大,造成这种差异的主要原因可能是番茄潜叶蛾入侵我国时间尚短,可能是由同一种群入侵,通过苗木运输、番茄产品的输送等在多个地点扩散。
本文初步分析了番茄潜叶蛾的遗传多样性,为番茄潜叶蛾部分种群遗传结构的研究提供了分子水平的基础资料及试验证据。但是,ISSR分子标记在估算种群遗传的变异上还存在一些缺点,如对于标记的显性基因,不能区分纯合体及杂合体等,为各种群遗传变异的分析所呈现的统计信息有局限(张民照和康乐,2002)。此外,本试验所采集的地理种群及个体数量有限,难以全面反映番茄潜叶蛾在我国各入侵地的具体情况,还需更深入研究。
ISSR研究结果表明,番茄潜叶蛾尚处在入侵早期阶段,且具备频繁入侵和多点的特征。防控上要注意加强检疫,阻绝多点入侵来源。
猜你喜欢
分子催化(2022年1期)2022-11-02
今日农业(2022年15期)2022-09-20
世界科学技术-中医药现代化(2022年3期)2022-08-22
昆明医科大学学报(2022年2期)2022-03-29
昆明医科大学学报(2021年6期)2021-07-31
昆明医科大学学报(2021年3期)2021-07-22
烟草科技(2021年6期)2021-06-24
湖南电力(2021年1期)2021-04-13
电脑知识与技术(2018年19期)2018-11-01
中学生物学(2018年8期)2018-03-01
