
金水缓纤组分方Ⅱ通过调控SETDB1/Snai1表观抑制A549细胞上皮间质转化*
2022-05-30 12:44许朋俐刘田田
中医学报 2022年6期
许朋俐,刘田田
1.河南中医药大学,河南 郑州 450046; 2.呼吸疾病中医药防治省部共建协同创新中心,河南 郑州 450046; 3.河南省中医药防治呼吸病重点实验室,河南 郑州 450046
特发性肺纤维化(idiopathic pulmonary fibrosis,IPF)是慢性纤维化性间质性肺炎的一种特殊类型[1],中医将其归于“肺痹”“肺痿”的病症范畴[2]。李建生教授将其病机概括为“正虚络痹积损”[3],并据多年临床经验拟定了金水缓纤方(ZL.201610877761.4)[4]。课题组在金水缓纤方的基础上,通过系统药理学结合体内外实验验证筛选出成分清楚、与原方疗效相当的金水缓纤组分方Ⅰ(ZL.201811415482.1),并进一步通过实验配比优化筛选出了疗效明显、组分明确的金水缓纤组分方Ⅱ(effective-component compatibility of Jinshui Huanxian formula Ⅱ,ECC-JHF Ⅱ),即淫羊藿苷、川陈皮素、贝母素甲、芍药苷、异甘草素(比例为10026.256.258)[5]。
特发性肺纤维化发病机制复杂,上皮间质转化(epithelial to mesenchymal transition,EMT)是其发生发展的重要病理机制之一[6]。课题组前期动物实验和细胞实验研究表明,金水缓纤方及其组分方能有效改善肺纤维化,缓解上皮间质转化[7-8]。锌指转录因子1(snail family zinc finger 1,Snai1)作为EMT的“主转录因子”,能通过抑制E-钙粘蛋白(E-cadherin)等上皮细胞标志蛋白的表达,激活N-钙粘蛋白(N-cadherin)等间充质细胞标志蛋白的表达,促进EMT的发生[9]。组蛋白赖氨酸N端甲基转移酶SET结构域分支型1(SET domain,bifurcated 1,SETDB1),也称为ESET或KMT1E,是一种对组蛋白H3第9位赖氨酸甲基化修饰的特异性甲基转移酶,参与常染色质基因的转录沉默。在一项乳腺癌的研究中发现,SETDB1通过对组蛋白的甲基化修饰,打破组蛋白甲基化和乙酰化的平衡,抑制Snail1基因表达[10]。基于此,本研究在细胞水平探讨金水缓纤组分方Ⅱ调控SETDB1及Snai1对EMT的表观抑制作用。
1 材料
1.1 细胞人非小细胞肺癌细胞(A549,细胞目录号:TCHu150),购自中国科学院典型培养物保藏委员会细胞库。
1.2 药物与试剂川陈皮素(nobiletin,NBN)、异甘草素(isoliquiritigenin,ISO)、芍药苷(paeoniflorin,Paeo)、淫羊藿苷(icariin,ICA)和贝母素甲(verticine,PEI)(成都曼思特生物科技有限公司,批号:MUST-16070901、MUST-17012503、MUST-16041901、MUST-17051810、MUST-16031101);转化生长因子-β1(transforming growth factor-β1,TGF-β1,美国PeproTech公司,货号:100-21C-10);吡非尼酮(pirfenidone,PFD,美国Sigma公司,货号:P2116);罗斯威尔公园纪念研究所(roswell park memorial institute,RPMI)-1640培养基、PBS缓冲液、乙二胺四乙酸(ethylene diamine tetraacetic acid,EDTA)-胰蛋白酶消化液、特级胎牛血清(以色列Biological Industries公司,货号:01-100-1ACS、02-024-1ACS、03-050-1ACS、04-001-1ACS);青霉素-链霉素溶液(武汉普诺赛生命科技有限公司,货号:PB1801220);Lipofectamine 3000细胞转染试剂、BCA蛋白定量试剂盒(赛默飞世尔公司,货号:L3000015、A53225);聚丙烯酰胺凝胶电脉(polyacrylamide gel electrophoresis,PAGE)试剂盒(上海雅酶生物科技有限公司,货号:PG112);抗GAPDH、纤维连接蛋白(fibronection,FN1)、α-平滑肌肌动蛋白(α-smooth muscle actin,α-SMA)、N-cadherin抗体(武汉三鹰生物技术有限公司,货号:60004-1-IG、15613-1-AP、14395-1-AP、22018-1-AP);抗E-cadherin抗体(美国GeneTex公司,货号:GTX100443);抗波形蛋白(Vimentin)抗体(英国Abcam公司,货号:ab92547);抗SETDB1、组蛋白H3第9位点赖氨酸三甲基化(histone He lysine 9 trimethylation,H3K9me3)抗体(美国Cell Signaling Technology公司,货号:93212S、13969S);抗Snai1抗体(美国Proteintech公司,货号:19099-1-AP);重组蛋白A(美国Millipore公司,货号:IP05);靶向setdb1的小干扰RNA(siRNA-setdb1,基因序列:F:5′-GAUCUAUCGAGGCUCUACA-3′;R:5′-UGUAGAGCGAUGUC-3′)委托上海吉玛制药技术有限公司合成。
1.3 仪器TS100型倒置显微镜(日本Nikon公司);3131型CO2培养箱(美国Thermo公司);1658001型基础电泳仪电源、1703930型小垂直板电泳槽、1645050型小型转印槽、ChemiDoc MP型全能型成像系统(美国BIO-RAD公司)。
2 方法
2.1 细胞培养用含有10%胎牛血清及1%青霉素-链霉素溶液的RPMI-1640完全培养基培养A549细胞,将其置于5%CO2、37 ℃恒温培养箱中培养,定期在倒置显微镜下观察细胞状态,每两天进行一次换液,当细胞生长面积达到细胞培养皿底面积的90%左右时,进行细胞传代处理,以保证细胞在对数生长期内。
2.2 药物制备及给药按比例称量NBN、ISO、Paeo、ICA和PEI并溶解于二甲基亚砜(dimethyl sulfoxide,DMSO)溶液,配成浓度为61.25 g·L-1的母液,过滤除菌后放-20 ℃冰箱备用;用含5% BSA的PBS将TGF-β1溶解至10 mg·L-1,放-80 ℃备用;PFD用DMSO配成0.4 g·L-1的溶液,放 -80 ℃ 备用。使用时用RPMI-1640完全培养基将以上药物稀释至工作浓度,加至细胞培养皿中。
2.3 A549细胞活力的检测将生长状态较好的A549细胞接种于96孔细胞培养板中,每孔接种细胞数为1×104个,设置6个复孔。放入37 ℃、5%CO2恒温细胞培养箱中培养24 h,待其生长密度约为70%时,加入不同浓度(0 mg·L-1、3.83 mg·L-1、7.66 mg·L-1、15.31 mg·L-1、30.63 mg·L-1、61.25 mg·L-1、122.50 mg·L-1)的ECC-JHF Ⅱ培养48 h;每孔加10 μL CCK8溶液(注意不要产生气泡,以防影响酶标仪读数),于 37 ℃ 恒温箱孵育1 h后用酶标仪测定450 nm处的吸光值。
2.4 Western Blot检测A549细胞中EMT相关蛋白表达水平将生长状态较好的A549细胞接种于6孔细胞培养板中,每孔接种1×105个细胞,放入37 ℃、5%CO2恒温细胞培养箱中培养24 h,待其生长密度约为70%时,将细胞分为空白组、TGF-β1组、ECC-JHF Ⅱ组和PFD组,空白组和TGF-β1组加入2 mL含2 μL DMSO的RPMI-1640完全培养基,ECC-JHF Ⅱ和PFD组分别加入含相应药物的培养基,作用12 h后,除空白组外,各组加入5 μg·L-1TGF-β1诱导12 h、24 h或36 h后,用RIPA蛋白裂解液提取蛋白,在冰上裂解30 min后,12 000×g、4 ℃离心15 min。使用BCA蛋白测定试剂盒测定蛋白浓度后,加入5×上样缓冲液,将其调整至相同蛋白浓度,100 ℃煮沸5 min。凝胶电泳将其分离并转至PVDF膜上。5%脱脂奶粉室温封闭2 h后,孵育一抗(E-cadherin、N-cadherin、GAPDH稀释比例为15 000;FN1、α-SMA稀释比例为 12 000;Vimentin稀释比例为12 500;Snai1稀释比例为1500;SETDB1、H3K9me3稀释比例为11 000),4 ℃过夜。次日取出蛋白条带,TBST洗4次,每次10 min,室温孵育二抗(稀释比例为15 000)2 h。经过化学发光法显影,用Image J软件测量灰度值后分析各组差异性。
2.5 siRNA转染A549细胞后EMT相关蛋白表达水平的检测将状态较好的A549细胞以2×105个/孔的密度接种于6孔细胞培养板中,放入恒温培养箱(37 ℃、5%CO2)培养24 h,细胞密度达到60%后分别转染空白质粒和siRNA-setdb1,转染空白质粒和siRNA-setdb1的细胞分别培养6 h后加入ECC-JHF Ⅱ预处理12 h,TGF-β1诱导24 h后记为Nc+ECC-JHF Ⅱ组和si-setdb1+ECC-JHF Ⅱ组。未经处理的转染后细胞分别记为NC组和si-setdb1组。提取蛋白,用Western Blot方法检测EMT相关蛋白的表达。
2.6 染色质免疫沉淀(chromatin immunoprecipitation,ChIP)技术检测ECC-JHF Ⅱ对Snai1基因启动子上H3K9me3水平的影响将状态较好的A549细胞以2×105个/孔的密度接种于6孔细胞培养板中,分为空白组、TGF-β1组和ECC-JHF Ⅱ组,放入37 ℃、5%CO2恒温培养箱培养24 h,细胞贴壁后生长密度达到80%时,ECC-JHF Ⅱ组给予ECC-JHF Ⅱ预处理12 h之后采用TGF-β1处理24 h;TGF-β1组给予TGF-β1处理24 h。通过甲醛交联与超声破碎细胞获取样本,加入与琼脂糖共价结合的重组蛋白A、H3K9me3抗体、IgG抗体,4 ℃ 过夜。次日加入与琼脂糖共价结合的重组蛋白A进行免疫复合物沉淀后,进行解交联并回收DNA,通过qPCR测定目的基因含量。

3 结果
3.1 ECC-JHF Ⅱ对A549细胞活力的影响CCK8结果显示,与0 mg·L-1ECC-JHF Ⅱ组比较,122.5 mg·L-1ECC-JHF Ⅱ组细胞活力显著降低(P<0.01);3.83~61.25 mg·L-1ECC-JHF Ⅱ组细胞活力无显著变化(P>0.05),可选用该浓度范围内的ECC-JHF Ⅱ进行后续实验。见表1。

表1 ECC-JHF Ⅱ对A549细胞活力的影响
3.2 ECC-JHF Ⅱ浓度筛选与空白组相比,TGF-β1 组E-cadherin蛋白表达水平显著下调(P<0.01),N-cadherin、FN1、Vimentin和α-SMA蛋白表达水平显著上调(P<0.01);与TGF-β1组相比,61.25 mg·L-1ECC-JHF Ⅱ组及PFD组E-cadherin蛋白表达水平显著上调(P<0.01),N-cadherin、FN1、Vimentin和α-SMA蛋白表达水平显著下调(P<0.01),所以后续实验ECC-JHF Ⅱ的使用浓度为 61.25 mg·L-1。见图1,表2。
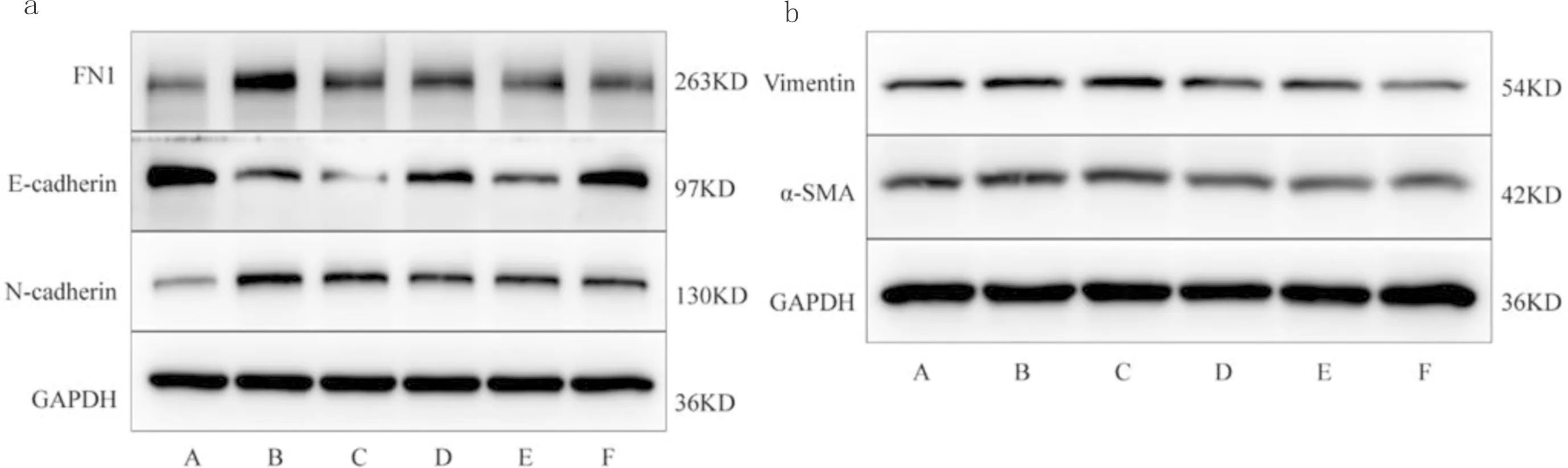
注:n=3;a:FN1、E-cadherin、N-cadherin的蛋白表达图;b:Vimentin、α-SMA的蛋白表达图;A:空白组;B:TGF-β1组;C:30.63 mg·L-1 ECC-JHF Ⅱ组;D:61.25 mg·L-1 ECC-JHF Ⅱ组;E:91.88 mg·L-1 ECC-JHF Ⅱ组;F:PFD组
表2 不同浓度ECC-JHF Ⅱ对FN1、E-cadherin、N-cadherin、Vimentin、α-SMA蛋白表达的影响
3.3 ECC-JHF Ⅱ有效改善EMT相关蛋白及SETDB1、H3K9me3蛋白表达水平与空白组相比,TGF-β1组E-cadherin、SETDB1和H3K9me3蛋白表达显著下调(P<0.01),FN1、α-SMA、N-cadherin、Vimentin、Snai1蛋白表达显著上调(P<0.01);与TGF-β1组相比,ECC-JHF Ⅱ组和PFD组SETDB1、H3K9me3和E-cadherin蛋白表达显著上调(P<0.01),FN1、α-SMA、N-cadherin、Vimentin、Snai1蛋白表达显著下调(P<0.01)。见图2,表3-表4。
表3 ECC-JHF Ⅱ对FN1、α-SMA、Vimentin、N-cadherin蛋白表达的影响

注:a:FN1、α-SMA、E-cadherin、N-cadherin、Vimentin的蛋白表达图;b:Snail、H3K9me3、SETDB1的蛋白表达图;n=3;A:空白组;B:TGF-β1组;C:ECC-JHF Ⅱ组;D:PFD组
表4 ECC-JHF Ⅱ对Snai1、E-cadherin、SETDB1、H3K9me3蛋白表达的影响
3.4 不同时间作用下ECC-JHF Ⅱ对TGF-β1诱导的EMT相关蛋白表达的影响与空白组相比,12 h TGF-β1组、24 h TGF-β1组SETDB1、H3K9me3、E-cadherin蛋白表达显著下调(P<0.01),Snail、FN1、N-cadherin蛋白表达显著上调(P<0.01);36 h TGF-β1组SETDB1、E-cadherin蛋白表达显著下调(P<0.01),FN1、Snai1、N-cadherin蛋白表达显著上调(P<0.01)。与12 h TGF-β1组相比,12 h TGF-β1+ECC-JHF Ⅱ组SETDB1、H3K9me3、E-cadherin蛋白表达显著上调(P<0.01),FN1蛋白表达显著下调(P<0.01);与24 h TGF-β1组相比,24 h TGF-β1+ECC-JHF Ⅱ组SETDB1、H3K9me3、E-cadherin蛋白表达显著上调(P<0.01),Snai1、FN1、N-cadherin蛋白表达显著下调(P<0.01);与36 h TGF-β1组相比,36 h TGF-β1+ECC-JHF Ⅱ组E-cadherin蛋白表达显著上调(P<0.01),Snai1、N-cadherin蛋白表达显著下调(P<0.01)。见图3,表5。
表5 不同时间作用下ECC-JHF Ⅱ对EMT相关蛋白表达的影响

注:a:SETDB1、H3K9me3、Snail的蛋白表达图;b:FN1、N-cadherin、E-cadherin、的蛋白表达图;n=3;A:空白组;B:12 hTGF-β1组;C:12 hTGF-β1+ECC-JHF Ⅱ组;D:24 hTGF-β1组;E:24 hTGF-β1+ECC-JHF Ⅱ组;F:36 hTGF-β1组;G:36 hTGF-β1+ECC-JHF Ⅱ组
3.5 ECC-JHF Ⅱ改善TGF-β1诱导的Snai1启动子上H3K9me3表达下调与空白组相比,TGF-β1 组Snai1启动子上H3K9me3表达减少(P<0.05);与TGF-β1组相比,TGF-β1+ECC-JHF Ⅱ组Snai1启动子上H3K9me3表达增加(P<0.01)。见图4。

注:n=3;与空白组比较,1)P<0.05;与TGF-β1组比较,4)P<0.01
3.6 siRNA敲低SETDB1抑制ECC-JHF Ⅱ对EMT相关蛋白的改善作用与空白组相比,TGF-β1组SETDB1、H3K9me3、E-cadherin蛋白表达显著下调(P<0.01),FN1、N-cadherin、Snai1蛋白表达显著上调(P<0.01);与TGF-β1组相比,ECC-JHF Ⅱ组SETDB1、H3K9me3、E-cadherin蛋白表达显著上调(P<0.01),FN1、N-cadherin、Snai1蛋白表达显著下调(P<0.01);与Nc组相比,si-setdb1组SETDB1、H3K9me3、E-cadherin蛋白表达显著下调(P<0.01),Snai1、FN1、N-cadherin蛋白表达显著上调(P<0.05);与Nc+ECC-JHF Ⅱ组相比,si-setdb1+ECC-JHF Ⅱ组SETDB1、H3K9me3、E-cadherin蛋白表达下调(P<0.01),Snai1、FN1、N-cadherin蛋白表达上调(P<0.01)。见图5,表6。

注:n=3;A:空白组;B:TGF-β1组;C:Nc组;D:si-setdb1组;E:ECC-JHF Ⅱ组;F:Nc+ECC-JHF Ⅱ组;G:si-setdb1+ECC-JHF Ⅱ组
表6 si-setdb1抑制ECC-JHF Ⅱ对EMT相关蛋白表达的改善作用
4 讨论
近年来,特发性肺纤维化发病率呈上升趋势[11]。但目前治疗策略十分有限,临床推荐药物吡非尼酮和尼达尼布也只能起到延缓肺纤维化进展的作用,无法阻止甚或逆转其病理进程[12-13]。肺移植手术是其有效的治疗手段,但存在肺源不足及费用昂贵等问题。由此,亟需加强本病的防治研究。
肺纤维化发病机制复杂,EMT是其关键病理机制之一。肺纤维化发生后,肺内有大量肺纤维化细胞灶形成[12]。肺纤维化灶主要由成纤维细胞和肌成纤维细胞组成,它们分泌的大量胶原成为异常沉积的细胞外基质的重要组成成分。研究发现,肺纤维化中约有1/3的成纤维细胞和肌成纤维细胞都经EMT形成[14]。EMT发生后,细胞骨架重排,上皮细胞由卵圆形变为成长梭状,E-钙粘蛋白等上皮细胞标志蛋白表达下调,N-钙粘蛋白、波形蛋白、α-平滑肌肌动蛋白、纤维连接蛋白等间充质细胞标志蛋白表达增加[15]。
TGF-β1是诱导EMT发生的重要因子。在肺泡上皮细胞受到损伤后,机体为进行损伤修复而产生大量的致纤维化细胞因子,如TGF-β等。TGF-β与TGF-β Ⅱ型受体(TGF-βRⅡ)结合,募集并诱导Ⅰ型受体C端磷酸化,从而激活Smads,激活的Smad2/Smad3与受体分离后,与Smad4形成三聚体,并作为转录因子移位到细胞核内[16-17]。Smad3/4复合物响应TGF-β,靶向Snai1序列,激活Snai1的表达[18]。Snai1通过与Smad3/4协同作用,抑制E-cadherin等上皮细胞标志蛋白的表达,激活N-cadherin等间充质细胞标志蛋白的表达[19-20]。
表观遗传学修饰是指在不改变基因碱基序列的情况下,通过DNA或组蛋白的甲基化和乙酰化等引起可遗传的基因功能改变,进而引起细胞表型变化[21]。目前关于肺纤维化发病过程中表观遗传调控作用的研究甚少。已有研究表明,抗纤维化基因的超甲基化和促纤维化基因的低甲基化是导致成纤维细胞大量生成和肺瘢痕形成的原因[22-23]。在EMT过程中,基因表达的重编程伴随着组蛋白修饰的动态变化[24]。关于信号通路中相关因子的表观遗传学变化,特别是在EMT发生时如何控制EMT驱动基因的组蛋白修饰,研究较少。SETDB1作为转移抑制因子,可通过与Smad2/3形成阻遏复合物抑制肺癌转移,其在高度转移性肺癌细胞中受到强烈抑制,恢复SETDB1的表达则抑制了丝状伪足的形成、迁移和侵入[25]。此外,SETDB1可以抑制乳腺上皮细胞和癌细胞的EMT进程[10]。课题组前期研究显示,SETDB1通过H3K9me3的表观修饰可以直接调节Snai1的表达,驱动EMT基因重编程;TGF-β1诱导的A549细胞通过减弱SETDB1的表达以促进EMT的发生[26]。
中医药在肺纤维化的防治上有明显优势,中医认为肺纤维化常见虚实夹杂或本虚标实之证,治疗以扶正祛邪为要。黄云鉴等通过文献研究发现,肺纤维化的治疗以养肺化痰、益气养阴、养肺清热和活血通络为主,使用清热、活血、益气类中药的频率较高[27]。中医治疗肺纤维化在临床实践中有许多有效方剂,如孔祥文[28]用血府逐瘀汤和丹参饮合二陈汤加减治疗痰瘀互结型肺纤维化,以补肺汤及平喘固本汤加减治疗肺纤维化后期肺肾气虚证。临床观察显示益气活血固本方治疗气虚血瘀型肺纤维化临床有效,能改善临床症状并缓解肺功能下降[29];益气祛痰化瘀汤联合西医治疗能减轻IPF气虚兼痰瘀阻肺型患者胸闷、咳嗽、气短等主要症状,改善肺功能[30];“气阴两补”法治疗肺纤维化临床效果较好,能有效降低炎症因子水平,改善患者肺功能[31]。
李建生教授基于肺纤维化“正虚络痹积损”病机,以补益肺肾、活血化痰为主要治法,拟定了金水缓纤方,该方能明显提高患者生存质量,减缓疾病进展。课题组在此基础上通过组分优化配伍获得了与之疗效相当的金水缓纤组分方Ⅱ[5]。金水缓纤组分方Ⅱ能有效改善肺纤维化并减轻上皮细胞间质化[5];并可通过激活mTOR和自噬,抑制巨噬细胞的极化,从而减缓博来霉素诱导的大鼠肺纤维化[32]。金水缓纤组分方Ⅱ中的单体成分具有抑制上皮间质转化、成纤维细胞活化及抗氧化作用。其中,淫羊藿苷可抑制TGF-β1诱导的人支气管上皮细胞EMT,从而抑制气道重塑[33]。川陈皮素抑制TGF-β1诱导的A549细胞EMT,抑制细胞侵袭转移并抑制Snai1的表达[34]。异甘草素通过激活Nrf2的表达,发挥抗氧化作用[35],还可通过激活MRC-5细胞的自噬作用,抑制TGF-β1诱导的成纤维细胞活化[36]。贝母素甲具有抗炎、抗癌、治疗急性肺损伤的作用[37];芍药苷可通过增加E-cadherin表达,降低Snai1和α-SMA的表达,抑制TGF-β1引起的A549细胞EMT[38],并可减轻博来霉素引起的小鼠肺纤维化[39]。
本实验研究结果显示,金水缓纤组分方Ⅱ可通过增加SETDB1的表达,上调Snai1启动子上的H3K9me3水平,抑制Snai1及N-钙粘蛋白、波形蛋白、纤维连接蛋白、α-平滑肌肌动蛋白等间质细胞标志蛋白的表达、促进E-钙粘蛋白的表达,从而缓解EMT。为验证SETDB1在其中是否起关键作用,用siRNA沉默SETDB1基因的表达,观察金水缓纤组分方Ⅱ对EMT的抑制作用是否受到影响。结果显示,敲低SETDB1表达后,金水缓纤组分方Ⅱ缓解TGF-β1诱导的上皮间质转化作用受到抑制,这表明SETDB1对Snai1的表观调控作用在金水缓纤组分方Ⅱ抑制TGF-β1诱导的A549细胞EMT的过程中起了关键作用。本研究为从细胞层面进一步阐释金水缓纤组分方Ⅱ治疗肺纤维化的作用机制提供了实验依据。
综上,金水缓纤组分方Ⅱ可通过调控SETDB1/Snai1表现抑制A549细胞的上皮间质转化。本研究从表观遗传学角度阐释了金水缓纤组分方Ⅱ通过组蛋白甲基化修饰抑制Snai1表达,进而抑制上皮细胞间质转化的机制。由于研究中使用的A549细胞不能完全代表正常的人肺泡上皮细胞,后续研究可在其他细胞系或基因敲除小鼠中进行进一步验证。
猜你喜欢
上海师范大学学报·自然科学版(2022年3期)2022-07-11
家庭医学(2022年5期)2022-04-27
海燕(2022年2期)2022-02-18
三农资讯半月报(2020年11期)2020-06-21
健康之友(2020年1期)2020-03-24
分析化学(2019年3期)2019-03-30
东坡赤壁诗词(2018年1期)2018-03-31
饮食科学(2016年2期)2016-07-04
中国民族民间医药·下半月(2011年10期)2011-12-27
家庭医药(2009年8期)2009-09-30
