
辣椒FLA 基因家族的生物信息学分析
2022-05-30 13:36杨晶晶
中国蔬菜 2022年5期
李 彤 杨晶晶
(1 兰州财经大学农林经济管理学院,甘肃兰州 730101;2 甘肃农业大学园艺学院,甘肃兰州 730070)
阿拉伯半乳糖蛋白(arabinogalactan proteins,AGPs)是高等植物中重要的一类糖蛋白,广泛存在于细胞壁、质膜和细胞外基质中,在细胞增殖与生长、细胞分化、细胞程序性死亡、体细胞胚胎发生、花药发育与花粉管生长、合子分裂和胚胎发育等过程中发挥重要作用(Gaspar et al.,2001;马浩力 等,2015)。类成束阿拉伯半乳糖蛋白(fasciclinlike arabinogalactan proteins,FLAs)家族是AGPs家族的一个亚家族,除了含有类AGP 的糖基化区域,还含有1~2 个具有粘附作用的类成束蛋白(fasciclin-like,FAS)结构域(Johnson et al.,2003;钟静 等,2017)。
FLA 基因家族成员参与植物多种生命活动,在植物的生长发育过程中发挥着重要的调控作用(卞坤 等,2017)。在拟南芥中,突变体植株表现出侧根数目增加,根和叶片的再生能力减弱,表明在根的发育和细胞分化过程起重要作用(Johnson et al.,2011);基因在小孢子发育过程中,尤其是在花粉细胞壁发育过程中起重要作用(Li et al.,2010);具有FLA4 H2 区氨基酸替换的盐过度敏感突变体植株在一定盐离子浓度下表现出根表皮细胞异常膨胀,根长变短,表明与盐离子耐受性和细胞膨胀有关,并且能与ABA 协同作用参与ABA 信号通路调控,从而影响根的发育(Johnson et al.,2003);与与次生细胞壁发育有关,与的双突变体茎机械强度下降,具体表现为:细胞壁中纤维素、阿拉伯糖和半乳糖含量减少(MacMillan et al.,2010)。在棉花的纤维细胞中,基因大量表达,参与棉纤维的发生和伸长,且过量表达可导致棉花纤维的长度增加(Huang et al.,2008,2013)。
FLA基因家族广泛分布在植物组织和细胞中,目前已在拟南芥()(Johnson et al.,2003)、水稻()(Ma &Zhao,2010)、小 麦()(Faik et al.,2006)等植物中鉴定出基因。现已发现拟南芥中有21 个基因,水稻中有27 个基因,棉花中有19 个基因,小麦中有34 个基因。植物中FLA 基因家族的挖掘,对深入了解基因的生物学功能及促进植物生产具有重要意义,但在辣椒(L.)中还鲜见关于FLA 基因家族成员的报道。因此,本试验采用生物信息学分析的方法,对辣椒基因组中FLA 基因家族成员进行全面鉴定,包括对其蛋白质理化性质、系统进化树、保守基序、基因结构、染色体定位及启动子顺式作用元件等进行分析,旨在揭示辣椒FLA 基因家族成员的数量、结构和进化特点,进一步为该家族成员的功能研究提供理论依据。
1 材料与方法
1.1 辣椒FLA 基因家族成员的鉴定
根据已报道的拟南芥FLA 基因家族的序列号,在拟南芥基因组数据库TAIR(http://www.arabidopsis.org)下载得到21 条FLAs 蛋白序列作为探针序列,并在茄科植物基因组数据库(sol genomics network,SGN)中的辣椒品种遵辣1 号基因组(Qin et al.,2014)数据库进行BLASTP(evalue <l),获得同源的辣椒候选基因(Schultz et al.,2002;Fernandez-Pozo et al.,2015)。然后利用Pfam〔Pfam:Home page(xfam.org)〕 和SMART〔SMART:Main page(emblheidelberg.de)〕数据库鉴定候选基因是否含有FAS结构域,删除冗余的基因,进而确定辣椒FLA 基因家族成员(El-Gebali et al.,2019;Letunic et al.,2021)。最后利用在线网站ProtParam(http://web.expasy.org/protparam/)分析辣椒FLA 基因家族成员编码的蛋白质序列长度、分子量及等电点等理化性质(Wilkins et al.,1999),利用Euk-mPLoc 2.0(http://www.csbio.sjtu.edu.cn/bioinf/euk-multi-2/)进行亚细胞定位预测(Chou &Shen,2010)。
1.2 辣椒FLA 基因家族的蛋白系统进化、保守基序、基因结构和染色体定位分析
利用MEGA-X 构建辣椒和拟南芥FLAs 蛋白序列的系统进化树,序列比对方法为MUSCLE,构建方法为邻接法(neighbor-joining,NJ),Bootstrap的重复次数设置为1 000,剩余设置选择默认选项,然后利用在线网站Evolview(https://evolgenius.info/evolview-v2)对进化树进行可视化(Saitou &Nei,1987;He et al.,2016;Kumar et al.,2018)。利用MEME 5.2.0〔MEME-Submission form(memesuite.org)〕软件分析辣椒FLAs 蛋白保守基序,参数设置模体基序长度最小为15 aa,最大为150 aa,保守序列数目最大为10(Bailey et al.,2015)。于在 线 网 站GSDS 2.0(http://gsds.gao-lab.org/index.php)中输入辣椒FLAs 蛋白的基因序列和编码序列(coding sequence,CDS),获得辣椒的基因结构(Hu et al.,2015)。通过TBtools 提取辣椒各条染色的总长,再依据辣椒FLA 基因家族的起始位置,利用Mapchart 2.3 进行染色体定位分析及作图(Voorrips,2002;Chen et al.,2020)。
1.3 辣椒FLA 基因家族启动子顺式作用元件分析
利用TBtools v1.082 工具提取辣椒编码序列的起始密码子上游2 000 bp 序列,利用PlantCARE 预测辣椒顺式作用元件,结果用TBtools v1.082进行可视化(Rombauts et al.,1999;Chen et al.,2020)。
2 结果与分析
2.1 辣椒FLA 基因家族成员的鉴定
通过BLASTP 从辣椒全基因组中获得25 个辣椒FLAs基因家族成员,命名为~。理化性质分析结果表明(表1),25 个都含有1~2 个FLA 基因家族特有的FAS 结构域,其中、、、、、、包含2 个FAS 结构域,表明25 个基因均为辣椒的FLA 基因家族成员;CaFLAs 的蛋白长度在201~449 aa 之间,平均长度为328 aa;CaFLAs 蛋白分子量在20.73~49.14 kDa 之间;蛋白理论等电点在4.44~8.74 之间,大多数为酸性蛋白。亚细胞定位分析结果显示,辣椒FLA 基因家族中除编码的蛋白既定位在质膜又定位在细胞核外,其余均定位于质膜。
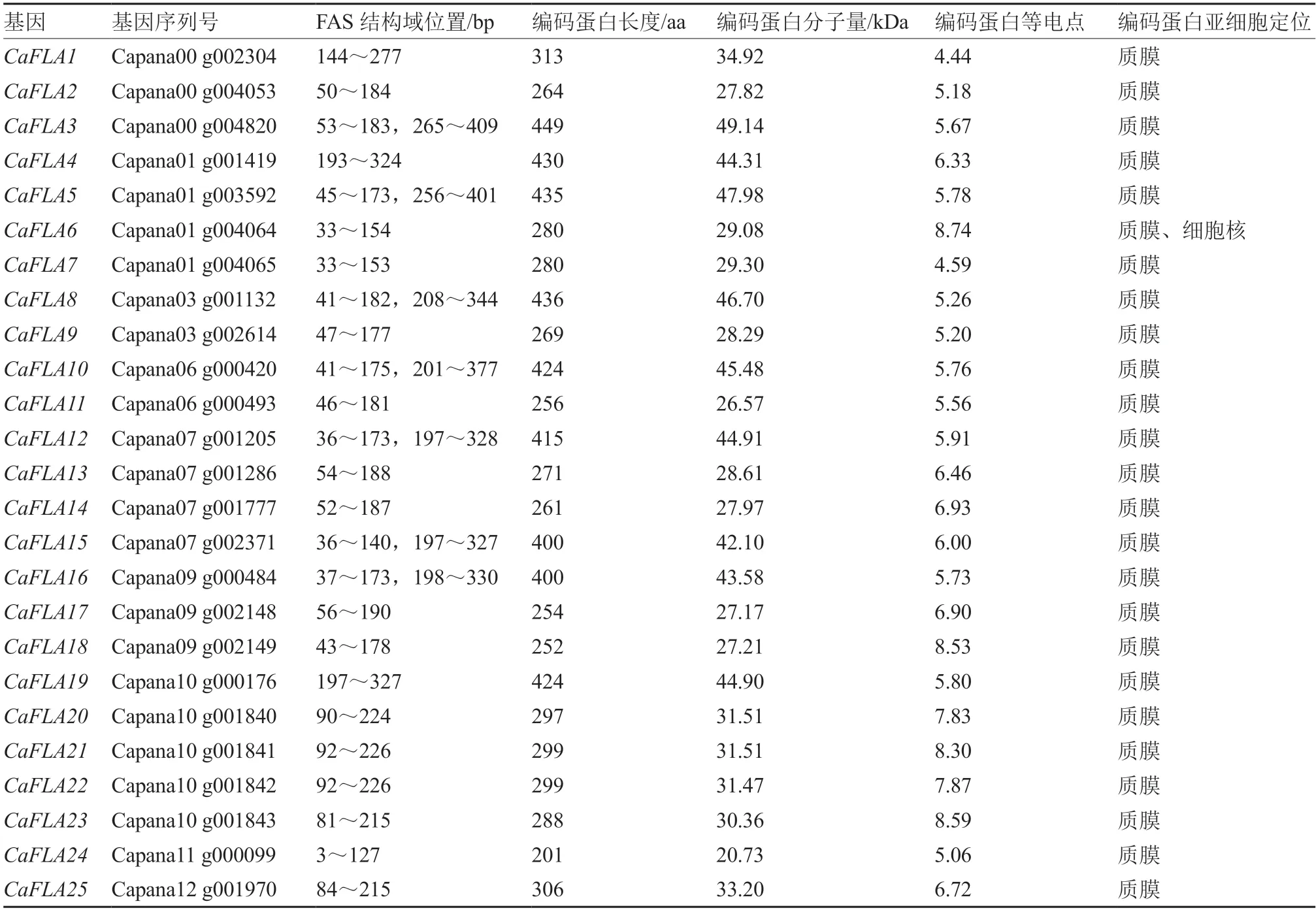
表1 辣椒FLA 基因家族成员基本特征
2.2 辣椒FLAs 蛋白系统进化树分析
如图1 所示,辣椒和拟南芥的FLAs 蛋白在蛋白序列上相对保守,且大多数Boostrap 大于70,表明FLAs 在辣椒和拟南芥进化过程中相对保守。25 个CaFLAs 蛋白被分为3 个亚家族,其中CaFLA4、CaFLA6、CaFLA7、CaFLA8、CaFLA9、CaFLA10、CaFLA12、CaFLA15、CaFLA16 和CaFLA19 进化关系较近,聚为第一亚族(Group1);CaFLA2、CaFLA11、CaFLA13、CaFLA14、CaFLA17、CaFLA18、CaFLA20、CaFLA21、CaFLA22、CaFLA23 和CaFLA24 聚为第二亚族(Group2);CaFLA1、CaFLA3、CaFLA5和CaFLA25 聚为第三亚族(Group3)。CaFLA3/CaFLA5、AtFLA3/AtFLA7、AtFLA5/AtFLA14、CaFLA2/CaFLA13、CaFLA8/CaFLA10、CaFLA12/CaFLA16、CaFLA15/CaFLA19、AtFLA4/CaFLA4、AtFLA2/AtFLA11、CaFLA6/CaFLA7是直系同源对,且Boostrap 为100,表明两者在蛋白结构方面相似。

图1 辣椒和拟南芥FLAs 蛋白的系统进化树
2.3 辣椒FLA 基因家族保守基序及基因结构分析
利用MEME 5.2.0 在线软件对辣椒25 个CaFLAs 蛋白进行保守基序分析,结果如图2 所示。位于同一亚族的CaFLAs 蛋白具有相同的保守基序,如位于第一亚族的CaFLA4、CaFLA6、CaFLA7、CaFLA12、CaFLA15、CaFLA16、CaFLA19 都 含 有motif3、motif4、motif7、motif9基序,位于第二亚族的CaFLA2、CaFLA11、CaFLA13、CaFLA14、CaFLA17、CaFLA18、CaFLA20、CaFLA21、CaFLA22、CaFLA23 都含有motif1、motif2、motif3、motif5、motif10 基序,位于第三亚族的CaFLA1、CaFLA3、CaFLA5 都含有motif5 基序。经Pfam 比对可知,motif1、motif2、motif3、motif4、motif5、motif7、motif8、motif9、motif10 是FAS 结构域的一部分。同一亚族包含相同的保守基序,表明辣椒FLA 基因家族在遗传进化上是相对保守的。
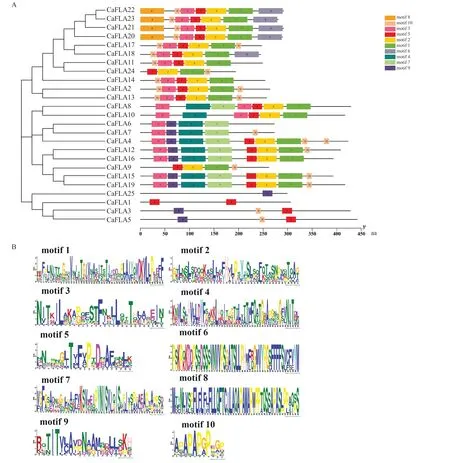
图2 辣椒FLAs 蛋白的保守基序分析
利用GSDS 2.0 在线网站对25 个基因进行基因结构分析,结果如图3 所示。有20 个辣椒FLA 基因家族成员不含内含子,其余成员(、、、和)只含有1 个内含子,表明位于同一亚族的基因具有相似的基因结构。

图3 辣椒FLA 基因家族成员基因结构分析
2.4 辣椒FLA 基因家族成员染色体定位分析
利用Mapchart 2.3 对25 个基因进行染色体定位,结果如图4 所示。25 个基因分别定位在0、1、3、6、7、9、10、11 号和12 号染色体上,在9 条染色体上不均匀分布,其中10 号染色体分布密度最高,分布了5 个基因;11 号和12 号染色体分布密度最低,仅存在1 个基因,其他染色体分布了2~4 个基因。

图4 辣椒FLA 基因家族成员染色体定位
2.5 辣椒FLAs 基因启动子顺式作用元件分析
辣椒启动子区域的顺式作用元件分析结果如图5 所示,25 个均拥有响应逆境胁迫的顺式作用元件,共含有17 种,其中分布最广的是光响应元件(light responsiveness),在25个中均有发现;分布较广的是厌氧胁迫响应元件(anaerobic induction),在20 个中被发现;其次是赤霉素响应元件(gibberellin responsiveness),在14 个中被发现。脱落酸响应元件(abscisic acid responsiveness)、生长素响应元件(auxin responsiveness)、防御与胁迫响应元件(defense and stress responsiveness)、低温响应元件(low temperature responsiveness)、茉莉酸甲酯响应元件(MeJA responsiveness)、分生组织表达元件(meristem expression)、干旱诱导的MYB 结合位点元件(MYB in drought)、水杨酸响应元件(salicylic acid responsiveness)、玉米醇溶蛋白代谢调节元件(zein metabolism regulation)、昼夜节律元件(circadian control)、缺氧特异性诱导元件(anoxic specific inducibility)、损伤响应元件(wound responsiveess)、胚乳表达元件(endosperm expression)、类黄酮生物合成的MYB 结合位点元件(MYB in flavonoid biosynthesis)不均一地分布在各个亚族的少数启动子上。表明辣椒基因参与辣椒的激素应答、胁迫响应(干旱、低温)、光响应等生物学过程。

图5 辣椒FLAs 基因启动子顺式作用元件分析结果
3 讨论与结论
本试验在辣椒中共鉴定出25 个基因,与拟南芥(21 个)(Johnson et al.,2003)、水稻(27个)(Ma &Zhao,2010)相比,FLA 基因家族成员相差不大,表明在物种进化过程中是相对保守的。对其理化性质的分析结果显示,辣椒FLA基因家族成员都含有1~2 个FLA 基因家族特有的FAS 结构域;CaFLAs 蛋白等电点在4.44~8.74 之间,多数为酸性蛋白;绝大多数CaFLAs 蛋白定位在质膜上,这可能与FLAs 参与细胞间信号转导有关(Wang et al.,2015;钟静 等,2017)。系统进化分析结果显示,辣椒和拟南芥FLAs 蛋白被聚类在3 个独立的进化分支中,每个进化分支中都有辣椒和拟南芥FLAs 的分布,表明该家族成员可能拥有共同的祖先。
利用MEME 5.2.0 在线软件分析得出,辣椒FLA 基因家族共有10 个保守基序,各基因中都有分布,属于高保守基序;研究发现不同亚族之间保守基序的相似性与亚族间的系谱进化密切相关,辣椒FLA 基因家族各亚族保守基序基本相同,表明辣椒FLA 基因家族在进化过程中具有高度保守性(黎帮勇 等,2015)。通过启动子顺式作用元件分析可知,辣椒启动子含有多种激素响应、逆境胁迫响应、光响应和植物生长发育相关的顺式作用元件,表明其可能具有较为广泛的生物学功能,可能参与多种生物代谢过程(郝小聪 等,2019)。值得注意的是,与激素响应有关的顺式作用元件几乎分布在所有的辣椒基因启动子中,表明辣椒FLA 基因家族可能参与辣椒生长和发育的许多过程,类似于其在拟南芥中的作用(马浩力 等,2015)。
猜你喜欢
快乐作文(1.2年级)(2020年8期)2020-09-10
中学生数理化·中考版(2016年7期)2016-12-07
中学物理·高中(2016年8期)2016-08-08
飞碟探索(2015年9期)2015-11-05
红领巾·探索(2015年9期)2015-09-10
风流一代·经典文摘(2011年1期)2011-07-15
恋爱婚姻家庭·养生版(2011年8期)2011-05-14
现代计算机(2009年9期)2009-12-02
职业·下旬(2009年8期)2009-10-12
金点子生意(2009年8期)2009-09-29
