
穗肥施氮量对不同穗型超级稻品种产量的影响及其机制
2022-05-29 05:42:44周沈琪张伟杨顾骏飞刘立军杨建昌
作物学报 2022年8期
刘 昆 黄 健 周沈琪 张伟杨 张 耗 顾骏飞 刘立军 杨建昌
穗肥施氮量对不同穗型超级稻品种产量的影响及其机制
刘 昆 黄 健 周沈琪 张伟杨 张 耗 顾骏飞 刘立军*杨建昌
扬州大学江苏省作物遗传生理重点实验室 / 江苏省粮食作物现代产业技术协同创新中心 / 江苏省作物基因组学和分子育种重点实验室, 江苏扬州 225009
施用氮素穗肥是水稻增产的一项重要管理措施, 但其用量对不同穗型超级稻品种的增产效应及其机制尚不明确。本研究选用3个穗型(以每穗粒数表示)差异较大的超级稻品种南粳9108 (小穗型)、扬两优6号(中穗型)和甬优1540 (大穗型)为材料, 在基蘖肥用量相同(162 kg N hm–2)的情况下, 研究了0、54、108、162和216 kg km–2五种穗肥施氮量对上述水稻品种产量的影响, 并观察了其对颖花分化退化及抽穗后相关形态生理指标的调控效应。结果表明: (1) 在0~216 kg hm–2穗肥施氮量范围内, 随施氮量增加, 水稻每穗粒数均逐渐增加, 结实率和粒重逐渐降低, 且穗肥施氮量越高结实率和千粒重下降越明显。南粳9108、扬两优6号和甬优1540三个水稻品种在穗肥施氮量分别为162~216、108~162和54~108 kg hm–2时产量最高。依据产量与穗肥施氮量曲线方程计算出上述3个品种高产最适穗肥施氮量分别为177.6~182.0、134.3~136.3和109.9~125.7 kg hm–2。(2) 总体而言, 穗型大的品种产量高, 穗型小的品种穗肥增产效应大。施用穗肥后小穗型品种二次颖花分化数和现存数增加幅度大是其增产效应高于中、大穗型品种的主要原因。(3) 3个水稻品种在高产的穗肥施氮量条件下, 高效叶面积率、粒叶比(颖花/叶、实粒/叶和粒重/叶)、非结构性碳水化合物(non-structural carbohydrate, NSC)转运量、糖花比、抽穗后0~40 d根系氧化力、颖花根活量和籽粒与根系玉米素(zeatin, Z)和玉米素核苷(zeatin riboside, ZR)含量高。相关分析表明, 不同穗型超级稻品种产量与以上指标基本呈显著或极显著正相关关系。上述结果表明, 穗肥施氮量应根据穗型大小进行调节。适宜的穗肥施氮量有助于在较高总颖花量的前提下, 保持抽穗后较高的高效叶面积率、粒叶比、NSC转运量、糖花比、根系氧化力、颖花根活量和籽粒与根系中Z + ZR含量, 这有利于维持较高的结实率和粒重, 从而提高水稻产量。
水稻; 穗肥施氮量; 穗型; 高产; 形态生理
超级稻品种对提高水稻产量和保证我国粮食安全有重要作用。超级稻品种不仅表现出较高的光合速率、较强的根系活力和“源–库–流”协调等生理功能, 同时还具有分蘖适中、剑叶直立、株高高、茎秆结实、抗倒伏、穗大、每穗粒数多等形态特征[1-3]。穗型与水稻产量和群体结构密切相关, 是水稻理想株型育种和高产栽培研究关注的热点问题[4]。稻穗为圆锥花序, 由穗轴、一次(一级)枝梗、二次(二级)枝梗、小穗梗和小穗(颖花)组成[5]。而常根据水稻每穗粒数的多少将其划分为: 小穗型、中穗型和大穗型[6]。小穗型品种单个颖花同化物供应充足, 籽粒灌浆充实从而有较高的结实率和千粒重, 但是每穗颖花数和总颖花数少限制了其产量的提高。中穗型品种源库较为协调, 产量构成因素间较为适中。大穗型品种具有库容大、产量潜力高等特点, 但其籽粒间灌浆充实不均匀而影响产量的进一步提高[7]。而水稻穗型大小及产量与细胞分裂素(cytokinin, CTK)含量密切相关[8]。而地上部干物重、叶面积、茎鞘非结构性碳水化合物(non-structural carbohydrate, NSC)积累和转运、叶片光合速率、根干重、根系氧化力和颖花根活量同样是影响水稻产量的重要形态生理指标。
氮是限制作物生长和谷物产量潜力的关键养分之一[9-11]。近60年来, 世界水稻产量不断增加, 部分原因是氮肥投入量的增加[12-13]。据报道, 目前我国稻田施氮量普遍超过180 kg hm–2, 超出世界平均水平的75%, 江苏省一些地区的平均施氮量更是高达300 kg hm–2 [14-15]。不合理的氮肥施用不但增加了生产成本, 还造成稻田氮素利用率急剧降低、稻米品质下降、土壤养分失衡, 并由此引发了一系列的环境问题[16]。此外, 过量施用氮肥还会增加水稻对倒伏、病虫害的敏感性, 最终导致水稻减产[17-18]。而合理的氮肥管理不仅可以提高粮食产量, 而且有利于降低稻田温室气体排放[19-21]。穗肥是稻田氮肥管理的重要组成部分。前人研究发现, 增加穗肥施氮量能够增强油菜素甾醇的生物合成和信号转导, 促进水稻颖花分化、减少其退化, 进而形成较高的每穗粒数[22]。穗肥施氮量还会影响籽粒蔗糖-淀粉代谢关键酶活性, 进而对结实率和千粒重产生影响, 最终影响产量[23]。而穗肥施氮量对于不同穗型超级稻品种产量的响应是否一致?其高产的生理机制是什么?迄今鲜有报道。
针对以上问题, 本研究选用3种不同穗型超级稻品种为材料, 研究了穗肥施氮量对其产量的影响, 并观察了抽穗后形态生理指标与产量形成的关系。旨在进一步揭示不同穗型超级稻品种产量形成的生理机制, 为超级稻品种选育以及栽培调控提供理论与实践依据。
1 材料与方法
1.1 供试材料与试验设计
试验于2018—2019年在扬州大学江苏省作物栽培生理重点实验室试验农场(32°30′N, 119°25′E)进行。以多年试验筛选出的大面积推广应用的不同每穗颖花数的超级稻品种为材料, 分别为南粳9108 (小穗型: 每穗粒数130粒左右)、扬两优6号(中穗型: 每穗粒数在220粒左右)和甬优1540 (大穗型: 每穗粒数为300粒左右)。上述3个水稻品种生育期均在150~155 d。5月12日至13日播种, 6月13日至14日人工插秧。大田前茬作物为小麦, 土壤类型为沙壤土, 耕作层含有机质21.8 g kg–1、有效氮104.5 mg kg–1、速效磷33.2 mg kg–1、速效钾88.4 mg kg–1。株行距为25.0 cm × 13.5 cm, 南粳9108为双本栽插, 扬两优6号和甬优1540为单本栽插。采用双因素裂区设计, 穗肥施氮量为主区, 品种为裂区。于穗分化期设置5种穗肥施氮量处理(1) 0N: 0 kg hm–2, 不施用穗肥; (2) 54N: 施用氮素54 kg hm–2; (3) 108N: 施用氮素108 kg hm–2; (4) 162N: 施用氮素162 kg hm–2; (5) 216N: 施用氮素216 kg hm–2。穗肥分促花肥、保花肥两次等量施用。各处理基、蘖肥用量均分别为135 kg hm–2和27 kg hm–2。基施过磷酸钙(含P2O513.5%) 300 kg hm–2和氯化钾(含K2O 62.5%) 200 kg hm–2。小区面积为20 m2, 随机区组排列, 重复3次, 计45个小区。主区间分筑田埂, 以防肥水串灌。水稻全生育期严格控制病虫草害, 以常规高产栽培方式进行田间管理。
1.2 取样与测定
1.2.1 颖花分化与退化 于水稻抽穗期(稻穗抽出1/3~1/2时)选取长势一致的挂牌主茎10根用于颖花分化、退化的测定, 参照Zhang等[24]方法观察并记录每穗一次颖花和二次颖花退化数和现存数。退化的颖花为具有白色的小颖花状薄膜痕迹, 部分可用肉眼观察计数, 部分颖花退化痕迹较小的在体视显微镜下观察计数。每穗颖花的分化数为退化数与现存数的总和。
1.2.2 干物质重与叶面积指数(leaf area index, LAI) 分别于抽穗后0、10、20、30、40 d和成熟期取代表性植株5穴(按群体平均茎蘖数取样), 将植株根、茎、叶、穗分离, 采用美国LI-COR公司生产的Li-Cor 3050型叶面积仪测定抽穗期总叶面积、有效叶面积(有效分蘖的叶面积)和高效叶面积(有效分蘖顶部3张叶片的叶面积)。随后分别放入烘箱, 在105℃杀青30 min, 后75℃烘干至恒重后称重。
1.2.3 茎鞘NSC积累量 参照Li等[25]方法, 使用上述抽穗期和成熟期烘干样品测定茎鞘NSC积累量。可溶性总糖的测定: 将粉碎的干样先过100目筛, 然后放入15 mL离心管中并向其中加入10 mL 80%乙醇, 80℃水浴20 min后冷却, 在经过2000转 min–1离心15 min, 取上清液, 重复提取3次。将合并的上清液定容至50 mL测定可溶性总糖(带有残渣的离心管放入80℃烘箱烘干后用于提取淀粉), 吸取0.5 mL上清液至10 mL离心管, 之后再加入1.5 mL超纯水和4 mL 0.2%蒽酮试剂, 沸水浴15 min, 冷却至室温测定其在620 nm处的吸光度值。淀粉含量测定: 向烘干的样品残渣中加入2 mL超纯水, 沸水浴15 min, 并不断搅动, 冷却后加入9.2 mol mL–1HClO42 mL, 振荡10 min, 再加入6 mL蒸馏水, 2000转 min–1离心20 min, 留取上清液, 再向残渣中加入2 mL 4.6 mol mL–1HClO4, 重复上述提取步骤, 最后合并上清液, 定容至50 mL, 最终淀粉含量测定方法与可溶性糖测定方法相同。NSC积累量为可溶性糖与淀粉积累量之和。
1.2.4 玉米素(zeatin, Z)和玉米素核苷(zeatin riboside, ZR)含量 分别于抽穗后0、10、20、30和40 d剥取各处理5株穗中部籽粒与根系液氮速冻后于−80℃冰箱中保存, 参照Liu等[26]方法测定籽粒及根系Z + ZR含量。
1.2.5 根系氧化力 于上述相同时期, 各小区取代表性植株挖根3穴(每穴以稻株基部为中心, 挖取 20 cm × 20 cm × 20 cm 的土块), 装于70目的筛网袋中, 先用流水再用农用压缩喷雾器将根冲洗干净, 称取根鲜重, 取部分根参照Meng等[27]方法测定根系氧化力。
1.2.6 考种与计产 成熟期各小区调查100株穗数, 并按平均穗数取样10穴手工脱粒, 考查水稻每穗粒数、结实率和千粒重等产量构成因素。结实率采用水漂法测定。各小区实收5 m2机器脱粒晒干后测定重量和含水率, 按13.5%的含水率折算为实收产量。
1.3 数据计算与统计分析
高效叶面积率(%) = 高效LAI/抽穗期LAI × 100;
颖花/叶(cm–2) = 总颖花数/抽穗期叶面积;
实粒/叶(cm–2) = 总实粒数/抽穗期叶面积;
粒重/叶(mg cm–2) = 籽粒产量/抽穗期叶面积;
NSC转运量(g stem–1) = 抽穗期茎鞘NSC积累量 – 成熟期茎鞘NSC积累量
NSC转运率(%) = (抽穗期茎鞘NSC积累量 – 成熟期茎鞘NSC积累量)/抽穗期茎鞘NSC积累量 × 100
糖花比(mg spikelet–1) = 抽穗期茎鞘NSC累积量/颖花量;
颖花根活量(μg α-napthyamine h–1spikelet–1) = 水稻结实期根量 × 单位干重根系氧化力/颖花量;
用Microsoft Excel 2003与SPSS软件统计分析数据, 用= 0.05最小显著极差法(LSD0.05)进行平均数显著性检验, SigmaPlot 11.0和R语言绘图。
2 结果与分析
2.1 方差分析
方差分析(表1)表明, 产量、每穗粒数、糖花比和颖花根活量在穗肥施氮量和品种间存在差异显著,而上述指标在年份、年份×穗肥施氮量、年份×品种、穗肥施氮量×品种(产量除外)和年份×穗肥施氮量×品种间差异均不显著。因此, 文中除产量外, 其余指标均用2019年试验数据表示。
2.2 产量
在0~216 kg hm–2穗肥施氮量范围内, 随施氮量的增加, 水稻每穗粒数均逐渐增加, 结实率和千粒重逐渐降低, 且穗肥施氮量越高结实率和千粒重下降幅度越明显。南粳9108 (小穗型)、扬两优6号(中穗型)和甬优1540 (大穗型) 3个水稻品种在穗肥施氮量分别为162~216、108~162和54~108 kg hm–2产量最高。相同穗肥条件下, 穗型越大的水稻品种产量越高。与不施穗肥相比, 施用穗肥使穗型较小的南粳9108产量提高11.9%~25.3%, 明显高于中、大穗型的扬两优6号和甬优1540的产量增幅, 分别为8.33%~16.50%和2.05%~11.90%。南粳9108 (小穗型)每穗粒数大幅增加(达13.2%~39.2%)以及结实率、千粒重下降幅度小(分别为1.42%~10.90%和1.16%~ 4.63%)是其产量增幅大的主要原因(表2)。
通过穗肥施氮量与产量的曲线方程, 计算出南粳9108、扬两优6号和甬优1540最适穗肥施氮量分别为177.6~182.0、134.3~136.3和109.9~125.7 kg hm–2。表明适宜穗肥施氮量应根据品种穗型大小进行调节。穗型较小的品种适当增加穗肥施氮量, 而穗型较大的品种可以适当减少穗肥施氮量, 这样有利于提高不同穗型超级稻品种产量(表3)。
2.3 颖花分化与退化
随穗肥施氮量增加, 不同程度提高了水稻一、二次颖花分化数、退化数和现存数, 且主要以二次颖花增加量为主。一次颖花退化率随穗肥施氮量增加而增加, 而二次颖花颖花退化率随穗肥施氮量增加呈先升后降的变化趋势。相同穗肥处理下, 不同穗型超级稻品种二次颖花分化数和现存数明

表1 穗肥施氮量对不同穗型超级稻品种产量和主要形态生理指标的方差分析
*、**分别表示在0.05和0.01水平上显著, ns表示差异不显著(> 0.05)。
*and**mean significant differences at the 0.05 and 0.01 probability levels, respectively. ns: not significant at> 0.05.
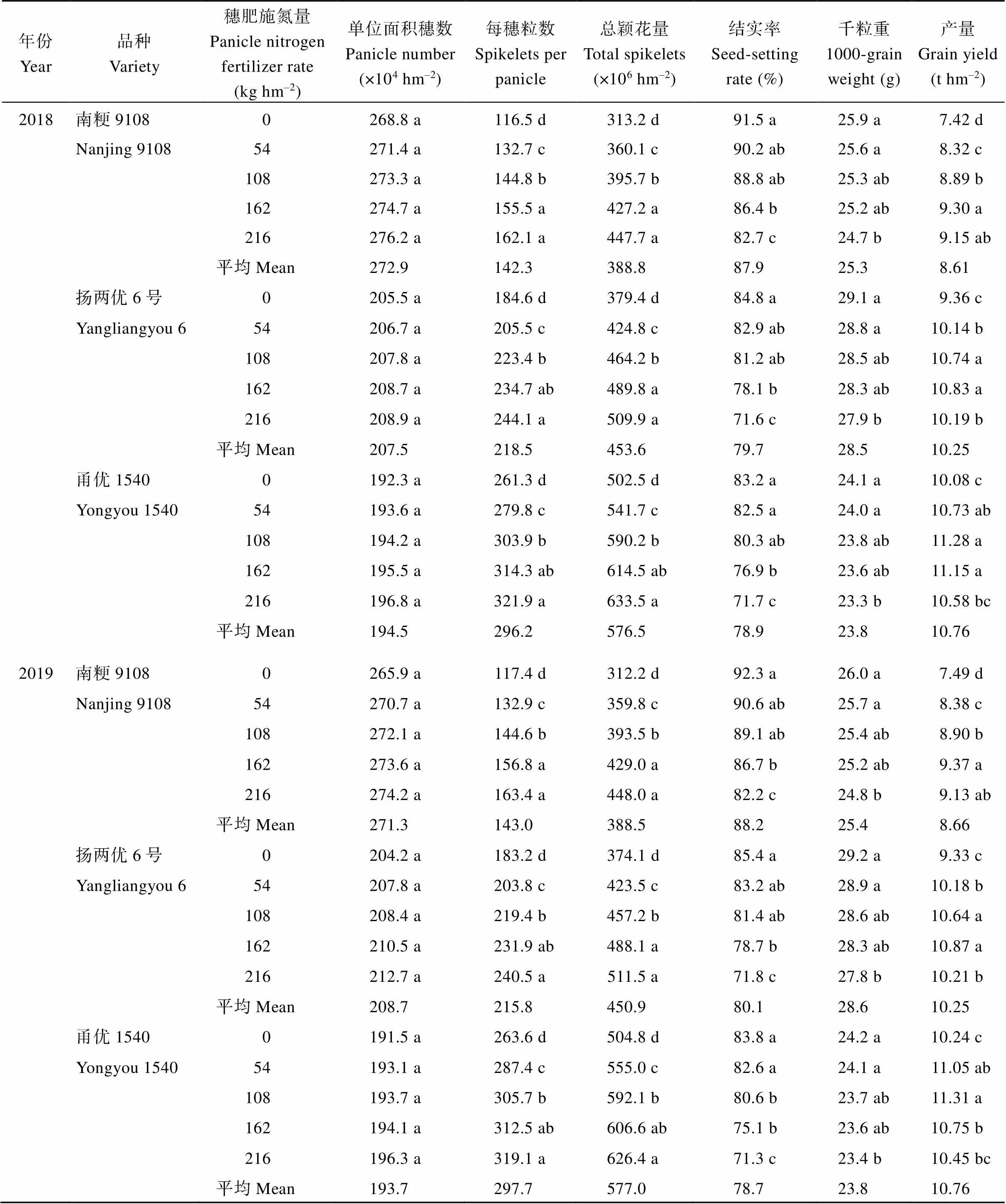
表2 穗肥施氮量对不同穗型超级稻品种产量及其构成因素的影响
标以不同字母表示在0.05水平上差异显著, 同栏内同一品种间比较。
Different lowercase letters within the same column indicate significance differences at the= 0.05 within the same variety.
2.4 LAI和粒叶比
在0~216 kg hm–2穗肥施氮量范围内, 随施氮量增加, 不同穗型超级稻品种抽穗期总LAI与高效LAI均提高。但南粳9108 (小穗型)、扬两优6号(中穗型)和甬优1540 (大穗型)高效叶面积率和粒叶比(颖花/叶、实粒/叶和粒重/叶)则分别在穗肥施氮量为162~216、108~162和108 kg hm–2条件下较高。与扬两优6号和甬优1540相比, 施用穗肥使南粳9108高效叶面积率和粒叶比增幅更大, 分别达5.69%~ 14.00%和8.34%~23.00% (表5)。

表3 不同穗型超级稻品种获得最高产量时的穗肥施氮量
opt: 获得最高产量时的穗肥施氮量;max: 最高产量。
opt: the optimal panicle nitrogen fertilizer rate for maximum grain yield;max: maximum grain yield.
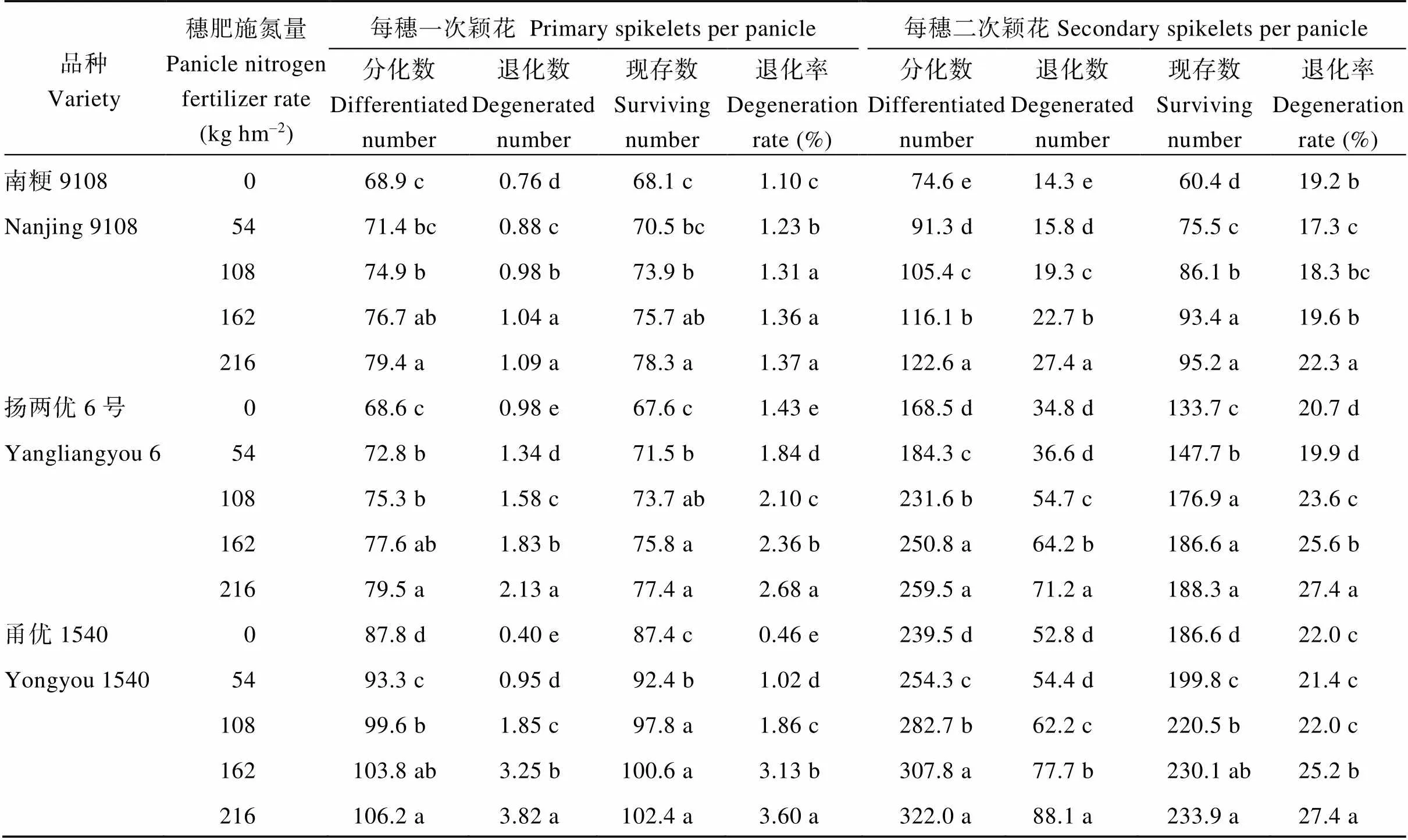
表4 穗肥施氮量对不同穗型超级稻品种一、二次颖花分化和退化的影响
标以不同字母表示在0.05水平上差异显著, 同栏内同一品种间比较。
Different lowercase letters within the same column indicate significant differences at the= 0.05 within the same variety.

表5 穗肥施氮量对不同穗型超级稻品种抽穗期叶面积指数(LAI)和粒叶比的影响
标以不同字母表示在0.05水平上差异显著, 同栏内同一品种间比较。LAI: 叶面积指数。
Different lowercase letters within the same column indicate significant differences at the= 0.05 within the same variety. LAI: leaf area index.
2.5 NSC转运量、NSC转运率和糖花比
南粳9108、扬两优6号和甬优1540分别在穗肥施氮量为162~216、108~162和54~108 kg hm–2条件下NSC转运量和糖花比高, 且相同穗肥条件下穗型越小的水稻品种NSC转运量越低、糖花比越高。不同穗型水稻品种NSC转运率均随穗肥用量增加而降低, 且穗型越大的水稻品种降幅越大(图1)。
2.6 根系氧化力
在抽穗后0~40 d范围内, 不同水稻品种根系氧化力均随抽穗后天数的增加而降低, 且穗型越大的品种根系活力越强。南粳9108、扬两优6号和甬优1540分别在穗肥施氮量为162~216、108~162和54~108 kg hm−2条件下根系氧化力高, 较不施穗肥处理增幅分别达16.1%~152.5%、5.69%~67.50%和4.12%~47.20% (图2)。

图1 穗肥施氮量对不同穗型超级稻品种NSC转运量、NSC转运率和糖花比的影响
0N、54N、108N、162N和216N分别代表穗肥施氮量为0、54、108、162和216 kg hm–2。标以不同字母表示在0.05水平上差异显著, 同一品种内比较。
0N, 54N, 108N, 162N, and 216N represent the panicle nitrogen fertilizer rate of 0, 54, 108, 162, and 216 kg hm–2, respectively. Different lowercase letters indicate significant differences at the= 0.05 within the same variety. NSC: non-structural carbohydrate.
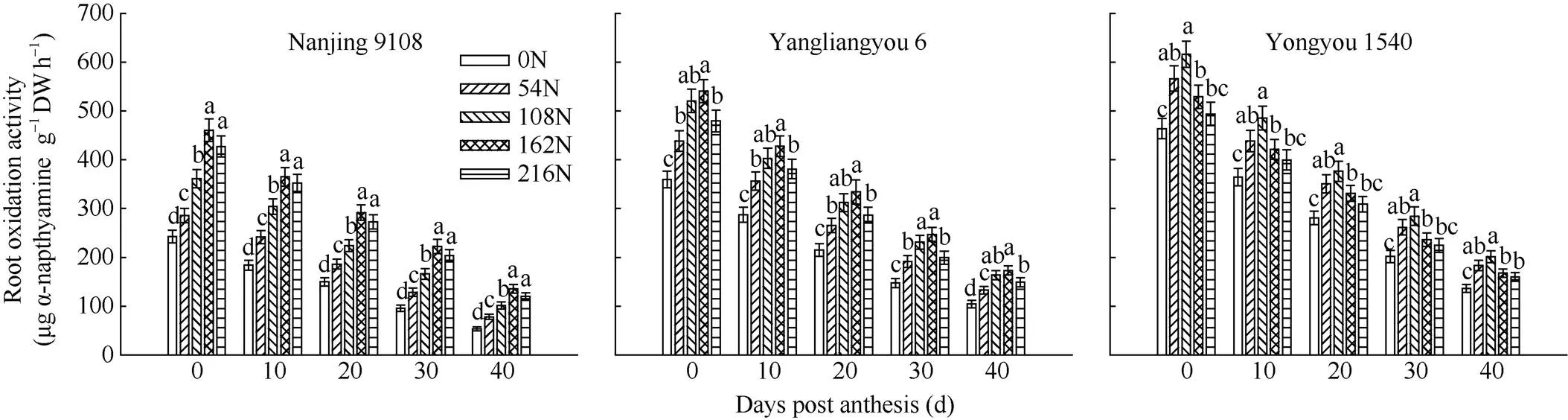
图2 穗肥施氮量对不同穗型超级稻品种根系氧化力的影响
0N、54N、108N、162N和216N分别代表穗肥施氮量为0、54、108、162和216 kg hm–2。标以不同字母表示在0.05水平上差异显著, 同一生育期内比较。
0N, 54N, 108N, 162N, and 216N represent the panicle nitrogen fertilizer rate of 0, 54, 108, 162, and 216 kg hm–2, respectively. Different lowercase letters indicate significant differences at the= 0.05 within the same growth stage.
2.7 根干重与颖花根活量
不同穗型超级稻品种单茎根干重于抽穗后0~40 d均随穗肥施氮量增加而增加。与不施穗肥相比, 施用穗肥显著提高了南粳9108 (小穗型)颖花根活量, 且在抽穗后0~20 d范围内较为明显, 达27.6%~168.9%, 增幅基本高于扬两优6号和甬优1540。穗型越大的品种单茎根干重较高、颖花根活量较低(图3)。
2.8 籽粒与根系Z + ZR含量
南粳9108、扬两优6号和甬优1540分别在穗肥施氮量为162~216、108~162和54~108 kg hm–2条件下籽粒Z + ZR含量高。且穗型越小的品种增幅越大, 而穗型越大的品种籽粒Z + ZR含量越高。根系Z + ZR含量变化趋势与籽粒Z + ZR含量变化趋势基本一致, 且根系Z + ZR含量明显高于籽粒(图4)。
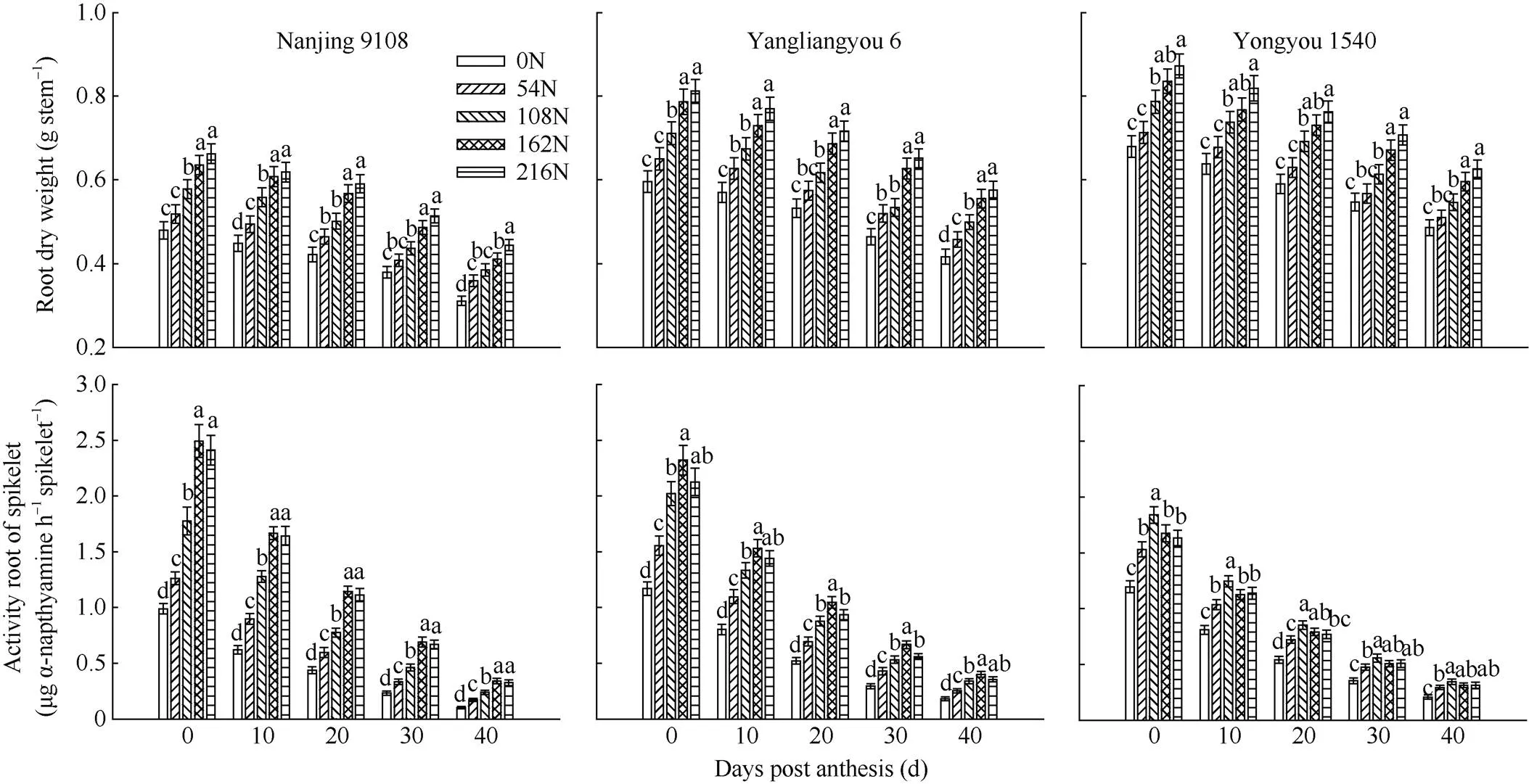
图3 穗肥施氮量对不同穗型超级稻品种根干重和颖花根活量的影响
0N、54N、108N、162N和216N分别代表穗肥施氮量为0、54、108、162和216 kg hm–2。标以不同字母表示在0.05水平上差异显著, 同一生育期内比较。
0N, 54N, 108N, 162N, and 216N represent the panicle nitrogen fertilizer rate of 0, 54, 108, 162, and 216 kg hm–2, respectively. Different lowercase letters indicate significant differences at the= 0.05 within the same growth stage.

图4 穗肥施氮量对不同穗型超级稻品种籽粒与根系Z + ZR含量的影响
0N、54N、108N、162N和216N分别代表穗肥施氮量为0、54、108、162和216 kg hm–2。标以不同字母表示在0.05水平上差异显著, 同一生育期内比较。
0N, 54N, 108N, 162N, and 216N represent the panicle nitrogen fertilizer rate of 0, 54, 108, 162, and 216 kg hm–2, respectively. Different lowercase letters indicate statistical significance at the= 0.05 within the same growth stage.
2.9 相关分析
相关分析表明, 南粳9108、扬两优6号和甬优1540产量基本均与高效叶面积率、粒叶比(颖花/叶、实粒/叶和粒重/叶)、NSC转运量、糖花比、抽穗后0~40 d根系氧化力、颖花根活量和幼穗与根系Z + ZR含量呈显著正相关关系。且根系和地上部主要生理指标对产量的贡献率最大, 表明根系和地上部主要生理指标的改善是不同穗型超级稻品种产量提高的关键因素(图5)。
3 讨论
3.1 穗肥施氮量对不同穗型超级稻品种产量的影响
水稻产量取决于库容的大小和籽粒灌浆充实的程度, 库大、源强、流畅是水稻高产群体的重要特征[6,28]。合理施用氮素穗肥是水稻增产的一项重要管理措施。随穗肥施氮量的增加, 不同穗型超级稻品种的每穗粒数均逐渐增加, 结实率和千粒重逐渐降低, 且穗肥施氮量越高结实率和千粒重下降幅度越明显,而产量呈先升后降的变化趋势。表明每穗粒数与结实率之间呈负相关关系, 穗肥施氮量过高不利于提高水稻产量。而二次颖花分化数和现存数大幅度增加是不同穗型超级稻品种每穗粒数显著增长的主要原因。氮肥是影响水稻产量的重要因素。前人通过Meta分析研究发现, 常规稻较杂交稻施肥增产效果显著, 且杂交稻适当减少氮肥施用仍能获得较高的水稻产量[18]。而对于不同穗型超级稻品种来说, 南粳9108、扬两优6号和甬优1540三个水稻品种在穗肥施氮量分别为162~216、108~162和54~108 kg hm–2产量最高。且穗型大的品种产量高, 穗型小的品种穗肥增产效应大。通过穗肥施氮量与产量的曲线方程, 计算出南粳9108、扬两优6号和甬优1540最适穗肥施氮量分别为177.6~182.0、134.3~136.3和109.9~125.7 kg hm–2。表明适宜穗肥施氮量应根据品种穗型大小进行调节。穗型较小的品种适当增加穗肥施氮量, 而穗型较大的品种可以适当减少穗肥施氮量, 这样有利于提高不同穗型超级稻品种产量。而施用穗肥后小穗型品种二次颖花分化数和现存数增幅大、结实率和千粒重降幅小是其增产效应高于中、大穗型品种的主要原因。
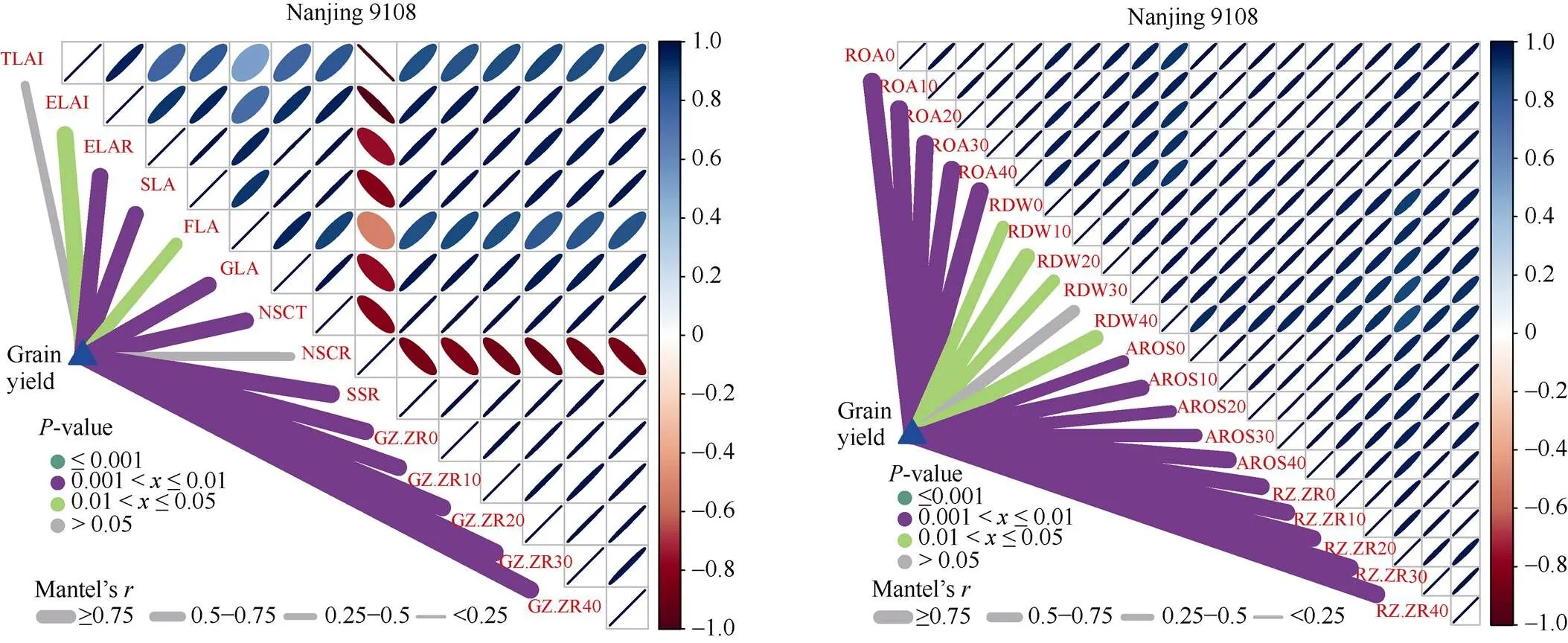
(图5)
TLAI、ELAI、ELAR、SLA、FLA、GLA、NSCT、NSCR、和SSR分别代表抽穗期总LAI、高效LAI、高效叶面积率、颖花/叶、实粒/叶、粒重/叶、NSC转运量、NSC转运率和糖花比。PZ.ZR0、PZ.ZR10、PZ.ZR20、PZ.ZR30和PZ.ZR40分别代表抽穗后0、10、20、30和40 d籽粒Z + ZR含量。ROA0、ROA10、ROA20、ROA30和ROA40分别代表抽穗后0、10、20、30和40 d根系氧化力。RDW0、RDW10、RDW20、RDW30、RDW40分别代表抽穗后0、10、20、30和40 d根干重。AROS0、AROS10、AROS20、AROS30和AROS40分别代表抽穗后0、10、20、30和40 d颖花根活量。RZ.ZR0、RZ.ZR10、RZ.ZR20、RZ.ZR30和RZ.ZR40分别代表抽穗后0、10、20、30和40 d根系Z + ZR含量。
TLAI, ELAI, ELAR, SLA, FLA, GLA, NSCT, NSCR, and SSR represent total LAI, effective LAI, high effective leaf area rate, spikelet/leaf area, filled grain/leaf area, grain weight/leaf area, NSC translocation amount, NSC remobilization, and sugar-spikelet ratio at heading stage, respectively. PZ.ZR0, PZ.ZR10, PZ.ZR20, PZ.ZR30, and PZ.ZR40 represent Z + ZR content in grains at 0, 10, 20, 30, and 40 days after heading, respectively. ROA0, ROA10, ROA20, ROA30, and ROA40 represent root oxidation activity at 0, 10, 20, 30, and 40 days after heading, respectively. RDW0, RDW10, RDW20, RDW30, and RDW40 represent root dry weight at 0, 10, 20, 30, and 40 days after heading, respectively. AROS0, AROS10, AROS 20, AROS 30, and AROS 40 represent activity root of spikelet at 0, 10, 20, 30, and 40 days after heading, respectively. RZ.ZR0, RZ.ZR10, RZ.ZR20, RZ.ZR30, and RZ.ZR40 represent Z + ZR content in roots at 0, 10, 20, 30, and 40 days after heading, respectively. NSC: non-structural carbohydrate.
穗肥通常分为促花肥和保花肥, 前者能促进颖花分化, 而后者有利于减少颖花退化[26]。高产栽培穗肥施用策略中, 大穗型水稻品种通常以施用保花肥为主, 而多穗型品种则以施用促花肥为主[28]。Zhang等[28]的研究表明, 在施氮总量相同的情况下, 小穗型品种单施促花肥、大穗型品种单施保花肥或粒肥有利于提高水稻产量和氮肥利用效率, 而中穗型品种单施促花肥或保花肥或分促花肥、保花肥和粒肥等量施用的产量和氮肥利用效率相近。本试验中, 3个不同穗型品种促花肥和保花肥均为等量施用,这样可以避免因二者施用比例不同对适宜施氮量造成的干扰, 从而更加有利于明确不同穗型水稻品种适宜穗肥用量的差异。未来仍有必要加强不同穗型水稻品种促花肥和保花肥二者施用比例对产量的影响及其机制的研究。
本研究中3个供试超级稻品种除了穗型大小有明显差异外, 品种类型也存在不同。如南粳9108为常规粳稻, 扬两优6号是籼型杂交稻, 而甬优1540为偏粳的亚种间籼粳杂交稻。本文只关注了穗型大小与穗肥用量的关系, 但品种类型不同是否也会造成穗肥适宜用量的不同?将来有必要针对此问题进一步开展研究。
3.2 穗肥施氮量对不同穗型超级稻品种产量影响的生理机理
根系是水稻的重要组成部分, 为水稻生长发育提供养分和水分, 是合成激素的重要场所[29]。CTK是水稻生长发育过程中重要的植物激素, CTK活性增强能够促进水稻每穗粒数增加, 提高水稻产量[26,30]。而氮肥能够促进CTK的生物合成和跨膜运输, 增强水稻根系活力, 提高水稻产量[11,31]。但穗肥过量施用均会导致不同穗型超级稻品种抽穗后0~40 d籽粒与根系Z + ZR含量、根系氧化力和颖花根活量降低, 不利于水稻产量的形成。
根干重是根系最重要的形态性状之一, 与根系生长发育密切相关, 同时根系生物量的增加对水稻产量的提高至关重要[27]。因为它增加了根毛对养分和水分的吸收和利用, 活跃的根系活力保证了营养、水分和植物激素的充足供应, 从而提高水稻产量[27]。但近年来, 一些研究者得到了与上述不同的结果。有研究者认为, 根系既是水分养分的吸收器官, 但同时又需要消耗地上部提供的光合产物用以根系的建成和维持生长[32]。与地上部相比, 用于生产单位根干物质量的能量是地上部的2倍。基于这种认识, 有人提出了“根系冗余生长”的观点, 即根冠比过大或根量过大会造成无效消耗而对产量产生不利的影响[33]。本研究结果表明, 与不施穗肥相比, 不同穗型超级稻品种单茎根干重于抽穗后0~40 d均随穗肥施氮量增加而增加, 而产量则呈现先升后降的变化趋势。表明根系生物量与产量存在二次曲线关系, 根干重过大并不利于提高不同穗型超级稻品种产量。而抽穗后0~40 d主要根系生理指标(根系Z + ZR含量、根系氧化力和颖花根活量)的改善是不同穗型超级稻品种在高总颖花量条件下, 维持较高的结实率和粒重, 进而提高产量的重要根系生理原因。
根系与地上部是相互依存的关系, 即良好的地上部生长有利于向根系提供光合产物维持根系活力,而较好的根系活力可向地上部提供充足的养分促进地上部生长, 从而保证水稻高产、超高产[34-35]。抽穗后较强的根系活力有利于水稻保持较高的剑叶光合速率。碳水化合物是植物体光合作用的主要产物, 其中NSC如淀粉、果聚糖、蔗糖等是参与植株生命代谢的重要物质[36]。前人常用抽穗期NSC积累量与颖花数之比, 即糖花比表示灌浆初期每朵颖花积累物质的多少[37]。抽穗期糖花比高, 表明抽穗前茎鞘中NSC累积量大, 每朵颖花获得的NSC多, 不仅有利于抽穗前花粉粒的充实完成, 而且可以增加抽穗至成熟期茎中同花物向籽粒的运转量, 促进花后胚乳细胞发育和籽粒充实[37-38]。在高施氮量条件下, 小穗型品种仍具有较高的茎鞘干物质积累和NSC浓度, 而高氮下加剧了大穗型品种氮素积累, 干物质积累增加, 但茎鞘NSC浓度大幅降低[6,36]。这是大穗型品种在高氮处理下糖花比较明显下降, 而小穗型品种的糖花比随穗肥施氮量的增加而增大的主要原因。因此, 抽穗后0~40 d糖花比和抽穗–成熟期NSC转运量高, 是不同穗型超级稻品种在高总颖花量条件下保持较高结实率和千粒重进而提高水稻产量的重要生理基础。
适宜的氮肥管理有利于维持水稻光合生产能力,促进水稻干物质积累, 从而提高水稻产量[12,39]。但穗肥施氮量过高会产生无效生长, 群体结构恶化, 不利于产量形成[40-41]。抽穗期叶面积组成及其配置直接关系到水稻群体质量的优劣, 影响水稻光合作用和产量形成[20]。有研究指出, 抽穗期较高的LAI有利于提高籼粳杂交稻产量[42]。而抽穗期较高的高效叶面积率和粒叶比(颖花/叶、实粒/叶和粒重/叶)有利于提高不同穗型超级稻品种产量。因此, 抽穗期较高的高效叶面积率和粒叶比是不同穗型超级稻品种高产的重要农艺性状。
4 结论
小穗型品种适当增加穗肥施氮量, 而中、大穗型品种适当减少穗肥施氮量, 有利于提高不同穗型超级稻品种产量。适宜的穗肥施氮量有助于在较高总颖花量的前提下, 保持抽穗后较高的高效叶面积率、粒叶比、糖花比、根系氧化力、颖花根活量和籽粒与根系Z + ZR含量, 这有利于维持较高的结实率和粒重, 从而提高产量。
[1] Huang M, Tang Q, Ao H, Zou Y. Yield potential and stability in super hybrid rice and its production strategies., 2017, 16: 1009–1017.
[2] Long J R, Ma G H, Wan Y Z, Song C F, Sun J, Qin R J. Effects of nitrogen fertilizer level on chlorophyll fluorescence characteristics in flag leaf of super hybrid rice at late growth stage., 2013, 20: 220–228
[3] 吴俊, 邓启云, 袁定阳, 齐绍武. 超级杂交稻研究进展. 科学通报, 2016, 61: 3787–3796.
Wu j, Deng Q Y, Yuan D Y, Qi S W. Progress of super hybrid rice research in China., 2016, 61: 3787–3796 (in Chinese with English abstract).
[4] Ke S, Liu X J, Luan X, Yang W, Zhu H, Liu G, Zhang G, Wang S. Genome-wide transcriptome profiling provides insights into panicle development of rice (L.)., 2018, 675: 285–300.
[5] Jang S, Lee Y, Lee G, Seo J, Lee D, Yu Y, Chin J H, Koh H J. Association between sequence variants in panicle development genes and the number of spikelets per panicle in rice., 2018, 19: 5.
[6] Liu K, Chen Y, Huang J, Qiu Y, Li S, Zhuo X, Yu F, Gao J, Li G, Zhang W, Zhang H, Gu J, Liu L, Yang J. Spikelet differentiation and degeneration in rice varieties with different panicle sizes., 2021, 11: e320.
[7] 顾俊荣, 韩立宇, 董明辉, 陈培峰, 乔中英. 不同穗型粳稻干物质运转与颖花形成及籽粒灌浆结实的差异研究. 扬州大学学报(农业与生命科学版), 2017, 38(4): 68–73.
Gu J R, Han L Y, Dong M H, Chen P F, Qiao Z Y. Studies on the difference of dry matter accumulation and transportation, spikelets formation and the grain filling ofrice varieties with different panicle types.(Agric Life Sci Edn), 2017, 38(4): 68–73 (in Chinese with English abstract).
[8] Han Y, Yang H, Jiao Y. Regulation of inflorescence architecture by cytokinins., 2014, 5: 669.
[9] Xue Y, Duan H, Liu L, Wang Z, Yang J, Zhang J. An improved crop management increases grain yield and nitrogen and water use efficiency in rice., 2013, 53: 271–284.
[10] Zhou W, Lyu T, Chen Y, Hu J, Zhang Q, Ren W. Late nitrogen application enhances spikelet number inhybrid rice (L.)., 2017, 74: 127–133.
[11] Gu J, Li Z, Mao Y, Struik P C, Zhang H, Liu L, Wang Z, Yang J. Roles of nitrogen and cytokinin signals in root and shoot communications in maximizing of plant productivity and their agronomic applications., 2018, 274: 320–331
[12] Wang Z, Zhang W, Beebout S S, Zhang H, Liu L, Yang J, Zhang J. Grain yield, water and nitrogen use efficiencies of rice as influenced by irrigation regimes and their interaction with nitrogen rates., 2016, 193: 54–69.
[13] Ju C, Zhu Y, Liu T, Sun C. The effect of nitrogen reduction at different stages on grain yield and nitrogen use efficiency for nitrogen efficient rice varieties., 2021, 11: 462.
[14] 白志刚, 张均华, 黄洁, 朱练峰, 曹小闯, 朱春权, 钟楚, 金千瑜. 氮肥运筹对水稻氮代谢及稻田土壤氮素迁移转化的影响. 生态学杂志, 2018, 37: 3440–3448.
Bai Z G, Zhang J H, Huang J, Zhu L F, Cao X C, Zhu C Q, Zhong C, Jin Q Y. Effects of nitrogen regime on nitrogen metabolism of rice and nitrogen transformation and translocation in paddy soils., 2018, 37: 3440–3448 (in Chinese with English abstract).
[15] Peng S, Buresh R J, Huang J, Zhong X, Zou Y, Yang J, Wang G, Liu Y, Hu R, Tang Q, Cui K, Zhang F, Dobermann A. Improving nitrogen fertilization in rice by site-specific N management: a review., 2010, 30: 649–656.
[16] Zhou C, Huang Y, Jia B, Wang S, Dou F, Samonte S O P, Chen K, Wang Y. Optimization of nitrogen rate and planting density for improving the grain yield of different rice genotypes in Northeast China., 2019, 9: 555.
[17] Zhang W J, Li G H, Yang Y M, Li Q, Zhang J, Liu J Y, Wang S H, Tang S, Ding Y F. Effect of nitrogen application rate and ratio on lodging resistance of super rice with different genotypes., 2013, 13: 63–72.
[18] Xu L, Yuan S, Wang X, Yu X, Peng S. High yields of hybrid rice do not require more nitrogen fertilizer than inbred rice: a meta-analysis., 2021, 10: e276.
[19] Wang Z, Gu D, Beebout S S, Zhang H, Liu L, Yang J, Zhang J. Effect of irrigation regime on grain yield, water productivity, and methane emissions in dry direct-seeded rice grown in raised beds with wheat straw incorporation., 2018, 6: 495–508.
[20] Wei H, Zhang H, Blumwald E, Li H, Cheng J, Dai Q, Huo Z, Xu K, Guo B. Different characteristics of high yield formation between inbredsuper rice and inter-sub-specific hybrid super rice., 2016, 198: 179–187.
[21] Chen Y, Li S, Zhang Y, Li T, Ge H, Xia S, Gu J, Zhang H, Lü B, Wu X, Wang Z, Yang J, Zhang J, Liu L. Rice root morphological and physiological traits interaction with rhizosphere soil and its effect on methane emissions in paddy fields., 2019, 129: 191–200.
[22] Zhang W, Zhu K, Wang Z, Zhang H, Gu J, Liu L, Yang J, Zhang J. Brassinosteroids function in spikelet differentiation and degeneration in rice., 2019, 61: 943–963.
[23] Xiong Q, Tang G, Zhong L, He H, Chen X. Response to nitrogen deficiency and compensation on physiological characteristics, yield formation, and nitrogen utilization of rice., 2018, 9: 1075.
[24] Zhang W, Fu L, Men C, Yu J, Yao J, Sheng J, Xu Y, Wang Z, Liu L, Yang J, Zhang J. Response of brassinosteroids to nitrogen rates and their regulation on rice spikelet degeneration during meiosis., 2020, 9: e201.
[25] Li G H, Pan J F, Cui K H, Yuan M, Hu Q Q, Wang W, Mohapatra P K, Nie L, Huang J, Peng S. Limitation of unloading in the developing grains is a possible cause responsible for low stem non-structural carbohydrate translocation and poor grain yield formation in rice through verification of recombinant inbred lines., 2017, 8: 1369.
[26] Liu K, Li T, Chen Y, Huang J, Qiu Y, Li S, Wang H, Zhu A, Zhuo X, Yu F, Zhang H, Gu J, Liu L, Yang J. Effects of root morphology and physiology on the formation and regulation of large panicles in rice., 2020, 258: 107946
[27] Meng T, Wei H, Li X, Dai Q, Huo Z. A better root morpho-physiology after heading contributing to yield superiority of/hybrid rice., 2018, 228: 135–146.
[28] Zhang Z, Chu G, Liu L, Wang Z, Wang X, Zhang H, Yang J, Zhang J. Mid-season nitrogen application strategies for rice varieties differing in panicle size., 2013, 150: 9–18.
[29] Bianco M D, Giustini L, Sabatini S. Spatiotemporal changes in the role of cytokinin during root development., 2013, 199, 324–338.
[30] Zheng C, Zhu Y, Wang C, Guo T. Wheat grain yield increase in response to pre-anthesis foliar application of 6-benzylaminopurine is dependent on floret development., 2016, 11: e0156627.
[31] Krouk G. Hormones and nitrate: a two-way connection., 2016, 91: 599–606.
[32] Chu G, Chen S, Xu C, Wang D, Zhang X. Agronomic and physiological performance of/hybrid rice cultivar under low nitrogen conditions., 2019, 243: 107625.
[33] Duque L O, Villordon A. Root branching and nutrient efficiency: status and way forward in root and tuber crops., 2019, 10: 237.
[34] Ning P, Li S, Li X, Li C. New maize hybrids had larger and deeper post-silking root than old ones., 2014, 166: 66–71.
[35] Santiago-Arenas R, Hadi S N, Fanshuri B A, Ullah H, Datta A. Effect of nitrogen fertilizer and cultivation method on root systems of rice subjected to alternate wetting and drying irrigation., 2019, 175: 388–399.
[36] Xu G W, Lu D K, Wang H Z, Li Y. Morphological and physiological traits of rice roots and their relationships to yield and nitrogen utilization as influenced by irrigation regime and nitrogen rate., 2018, 203, 385–394.
[37] Fu J, Huang Z H, Wang Z Q, Yang J C, Zhang J H. Pre-anthesis non-structural carbohydrate reserve in the stem enhances the sink strength of inferior spikelets during grain filling of rice., 2011, 123: 170−182.
[38] Zhen F, Zhou J, Mahmood A, Wang W, Chang X, Liu B, Liu L, Cao W, Zhu Y, Tang L. Quantifying the effects of short-term heat stress at booting stage on nonstructural carbohydrates remobilization in rice., 2020, 8: 194–212.
[39] Zhang J, Tong T, Potcho P M, Huang S, Ma L, Tang X. Nitrogen effects on yield, quality and physiological characteristics of giant rice., 2020, 10: 1816.
[40] Meng T, Wei H, Li C, Dai Q, Xu K, Huo Z, Wei H, Guo B, Zhang H. Morphological and physiological traits of large-panicle rice varieties with high filled-grain percentage., 2016, 15: 1751–1762.
[41] Huang L, Yang D, Li X, Peng S, Wang F. Coordination of high grain yield and high nitrogen use efficiency through large sink size and high post-heading source capacity in rice., 2019, 233: 49–58.
[42] Xu D, Zhu Y, Chen Z, Han C, Hu L, Qiu S, Wu P, Liu G, Wei H, Zhang H. Yield characteristics of/hybrids rice in the middle and lower reaches of the Yangtze River in China., 2020, 19: 2394–2406.
Effects of panicle nitrogen fertilizer rates on grain yield in super rice varieties with different panicle sizes and their mechanism
LIU Kun, HUANG Jian, ZHOU Shen-Qi, ZHANG Wei-Yang, ZHANG Hao, GU Jun-Fei, LIU Li-Jun*,and YANG Jian-Chang
Jiangsu Key Laboratory of Crop Genetics and Physiology / Jiangsu Co-innovation Centre for Modern Production Technology of Grain Crops / Jiangsu Key Laboratory of Crop Genomics and Molecular Breeding, Yangzhou University, Yangzhou 225009, Jiangsu, China
The application of panicle nitrogen fertilizer is an important management measure to increase rice yield, but its effect and mechanism of application rates on the yield increase of super rice varieties with different panicle sizes are still unclear. In this study, three super rice varieties with large differences in panicle sizes (indicated by the spikelet number per panicle) of Nanjing 9108 (small panicle size), Yangliangyou 6 (medium panicle size), and Yongyou 1540 (large panicle size) were selected as materials. Under the condition of the same amount of base-tiller fertilizer (162 kg N hm–2), the effects of five panicle nitrogen fertilizer rates (PNR) of 0, 54, 108, 162, and 216 kg hm–2on the yield of the above rice varieties were studied. And its regulatory effects on the differentiation and degeneration of spikelets and related morphophysiological indices after heading were observed. The results were as follows: (1) In the PNR range of 0–216 kg hm–2, the spikelet number per panicle gradually increased but the seed-setting rate and 1000-grain weight gradually decreased with the increase of PNR. And the higher the PNR, the more obvious the decrease of seed-setting rate and 1000-grain weight. Three rice varieties, Nanjing 9108, Yangliangyou 6, and Yongyou 1540, had the highest yields in the PNR of 162–216, 108–162, and 54–108 kg hm–2, respectively. According to the curve equation of grain yield and PNR, the optimal PNR for high yields of the above three varieties were calculated to be 177.6–182.0, 134.3–136.3, and 109.9–125.7 kg hm–2, respectively. (2) In general, rice varieties with large panicle sizes had higher yields, while rice varieties with small panicle sizes had greater yield-increasing effects of PNR. The number of differentiated and surviving secondary spikelets in rice varieties with small panicle size increased greatly after application of panicle nitrogen fertilizer, which was the main reason that the yield-increasing effect was higher than that of rice varieties with medium and large panicle size. (3) Under the condition of high-yield PNR, high effective leaf area ratio, grain-leaf ratio (spikelet/leaf area, filled grain/leaf area, and grain weight/leaf area), non-structural carbohydrate (NSC) translocation amount, sugar-spikelet ratio, root oxidation activity, activity root of spikelet and zeatin (Z) + zeatin riboside (ZR) content in grains and roots from 0–40 day(s) after heading of three rice varieties were high. Correlation analysis showed that the yields of super rice varieties with different panicle sizes and the above indicators basically had a significant or extremely significant positive correlation. These results indicated that the PNR should be adjusted according to the panicle size. The appropriate PNR was beneficial to maintain a high effective leaf area ratio, grain-leaf ratio, NSC translocation amount, sugar-spikelet ratio, root oxidation activity, activity root of spikelet, and Z + ZR content in grains and roots after heading under the premise of higher total spikelets. This helped to maintain a high seed-setting rate and grain weight, thereby ultimately increasing the grain yield.
rice; panicle nitrogen fertilizer rate; panicle size; high yields; morphology and physiology
通信作者(Corresponding author):刘立军, E-mail: ljliu@yzu.edu.cn, Tel: 0514-87972133
E-mail: 940057576@qq.com
2021-09-20;
2021-11-29;
2021-12-16.
10.3724/SP.J.1006.2022.12068
本研究由国家自然科学基金项目(32071947, 31871557), 江苏省农业科技自主创新项目(cx(18)3007), 江苏省作物遗传生理重点实验室开放课题(YSCL201807), 国家重点研发计划项目(2016YFD0300502, 2017YFD0301206), 江苏高校优势学科建设工程项目(PAPD)和江苏省研究生科研与实践创新计划项目(KYCX21_3235)资助。
This study was supported by the National Natural Science Foundation of China (32071947, 31871557), the Jiangsu Agriculture Science and Technology Innovation Fund (cx(18)3007), the Open Project of Jiangsu Key Laboratory of Crop Genetics and Physiology (YSCL201807), the National Key Research and Development Program of China (2016YFD0300502, 2017YFD0301206), the Priority Academic Program Development of Jiangsu Higher Education Institutions (PAPD), and the Postgraduate Research & Practice Innovation Program of Jiangsu Province (KYCX21_3235).
URL: https://kns.cnki.net/kcms/detail/11.1809.S.20211213.1000.002.html
猜你喜欢
Acta Mathematica Scientia(English Series)(2024年1期)2024-03-23 08:02:54
江西农业大学学报(2023年2期)2023-05-08 01:32:10
种子(2022年1期)2022-02-24 04:30:50
作物杂志(2022年6期)2022-02-03 04:56:22
北方水稻(2021年6期)2021-02-17 07:17:50
农家之友(2018年12期)2018-03-12 00:50:28
江苏农业科学(2017年17期)2017-11-15 02:46:54
福建稻麦科技(2015年1期)2016-01-06 12:22:06
发明与创新(2015年37期)2015-02-27 10:40:26
植物营养与肥料学报(2013年1期)2013-10-26 03:00:24
