
马铃薯与曼陀罗交互嫁接后药用生物碱的积累特征
2022-05-27 08:30郭开基李有涵郭华春
贵州农业科学 2022年5期
郭开基, 李有涵, 王 琼, 郭华春*
(1.云南农业大学 农学与生物技术学院, 云南 昆明 650201; 2.云南农业大学 机电工程学院, 云南 昆明 650201)
0 引言
【研究意义】曼陀罗(DaturastramoniumL.)是茄科曼陀罗属(Datura)的一年生草本植物,资源分布广泛,其富含多种生物碱,在植株中现已发现超过70种托品烷类生物碱(Tropane alkaloids,TAs)[1-2]。在曼陀罗的TAs中,阿托品、莨菪碱、(±)山莨菪碱和东莨菪碱是主要成分[3-4],这些生物碱对作物虫害防治有显著效果[5],也是现代医学广泛应用的抗菌[6]、抗癌[7-8]、消炎、镇痛[9-10]等有效药用成分,对曼陀罗药用生物碱的积累特征展开研究,为获取大量TAs进行开发利用具有重要的意义。【前人研究进展】TAs的生物合成以鸟氨酸、精氨酸和苯丙氨酸为起点[11-12],经多步酶促反应最后合成莨菪碱(Hyoscyamine)和阿托品(Atropine)[12],莨菪碱和阿托品的药物活性类似,但莨菪碱的作用强度强于阿托品[5,13]。莨菪碱可经H6H(Hyoscyamine 6 beta-hydroxylase)酶羟基化为山莨菪碱(Anisodamine),山莨菪碱为手性分子,根据其旋光性分为左旋山莨菪碱(-山莨菪碱)和右旋山莨菪碱(+山莨菪碱),两者除旋光性不同外,药理性质和药物活性强度基本一致[14]。(±)山莨菪碱可在H6H的催化作用下,继续羟基化形成东莨菪碱(Scopolamine)[15]。东莨菪碱被认为是TAs代谢途径中的最后一个生物碱,也是曼陀罗所有TAs中活性最强和药用价值最高的生物碱[16]。东莨菪碱在植株中含量很低,通常仅有1~1 000 μg/g[17-18]。已有研究表明,曼陀罗的TAs主要在根中合成,由维管组织运输至植株地上部,总生物碱在叶和种子中含量较高,茎中最低[4]。嫁接是果木、花卉和蔬菜品种改良常用的手段,通过利用接穗和砧木的各自优势,将优良性状集于一体[19-20]。嫁接技术在园艺作物的抗旱、抗涝和抗盐碱性状的改良中有重要作用[21-22],也是烟叶品质改良的有效方法[23]。大量研究表明,砧木和接穗的营养物质和遗传信息存在相互交流和转运[24],如以烟碱含量低,且钾素利用率高的烟草品种为砧木,可提高烟草接穗的钾素含量,并降低烟碱含量[25];利用不同辣椒品种进行嫁接,能够提高接穗辣椒的香草醛、水杨酸和木质素含量[26];杨怀亮等[27]发现,与自根嫁接的黄瓜相比,双砧木嫁接(将1个接穗嫁接到2株砧木上)可显著提高黄瓜游离氨基酸、类黄酮和单宁含量。【研究切入点】目前TAs的主要获取方式有TAs资源植物直接提取,基因工程生物合成和化学合成,但成本较高,且不易获取。为满足医疗和农业生产对TAs生物碱的需求,有必要对曼陀罗TAs的合成和积累特征开展研究。【拟解决的关键问题】为研究曼陀罗TAs的合成和积累规律,将曼陀罗与马铃薯进行交互嫁接,分析不同砧木和接穗中TAs含量,探究TAs在不同器官中的积累特性。同时重点关注嫁接后马铃薯块茎中是否能够富集TAs,探索利用块茎的高生物量特性来富集曼陀罗TAs的可能性,以期为进一步拓宽TAs的研究方法和制取方法提供参考。
1 材料与方法
1.1 试验材料
马铃薯品系滇薯1418(D1418)种薯、曼陀罗种子均由云南农业大学薯类作物研究所繁育保存。
1.2 试验方法
1.2.1 马铃薯和曼陀罗嫁接及取材方法 各植物材料种植于云南农业大学农场温室大棚。将曼陀罗种子播种,待幼苗出现第4片真叶后用于嫁接。将D1418种薯置于基质中催芽直至出苗,当幼苗长出3~4片叶时,从种薯上连根掰开后分株移栽,栽约20 d后选择生长一致的马铃薯幼苗用于嫁接。
两材料交互分别作砧木和接穗,即嫁接时以马铃薯幼苗为砧木,曼陀罗为接穗的嫁接组合(曼陀罗/D1418);以曼陀罗为砧木,马铃薯为接穗的嫁接组合(D1418/曼陀罗)。嫁接幼苗成活后,对嫁接组合D1418/曼陀罗进行培土,用土将接穗马铃薯嫁接伤口上部的2~5个节充分覆盖,以促进马铃薯茎基部叶腋处形成块茎。嫁接50 d后对曼陀罗/D1418嫁接组合的曼陀罗茎、叶、果和D1418的块茎进行取材;对D1418/曼陀罗嫁接组合的马铃薯茎、叶、叶腋处块茎和曼陀罗的根进行取材,每个样品3个生物学重复。将所取样品液氮速冻后,于-80℃保存。
1.2.2 用于TAs测定的植物样品前处理 在液氮中将各植物组织研磨成细粉,分别精确称取研磨后的样品各1.000 g置于具塞锥形瓶中。加入浓氨水-乙醇-乙醚(4︰5︰10,V/V)溶液5 mL,振荡混匀,并静置过夜。随后,进行超声破碎及提取(频率:40 kHz;功率:250 W;温度:25~30℃)2次,每次30 min。将提取液进行5 000 r/min离心10 min,吸取上清液至干净试管中,于60℃水浴挥干溶剂。试管底部残渣用0.5 mol/L硫酸1 mL复溶,再加入蒸馏水5 mL充分混匀。将上述获得的溶液转移到分液漏斗中,用氯仿10 mL萃取3次,合并上清液后水浴挥干。挥干后的残渣用甲醇定容至10 mL,并振荡混匀,取上清液1 mL所得溶液经0.22 μm微孔滤膜过滤后备用。
1.2.3 TAs标准品制备 精确称取适量硫酸阿托品、莨菪碱、(-)山莨菪碱、(+)山莨菪碱和氢溴酸东莨菪碱标准品,用色谱级甲醇溶解,并定容至5 mL,分别配制成1.0 mg/mL的标准品溶液。将硫酸阿托品稀释成0.11 μg/mL、0.22 μg/mL、1.1 μg/mL、2.2 μg/mL、11 μg/mL、22 μg/mL;将莨菪碱标准品稀释成0.148 μg/mL、0.296 μg/mL、1.48 μg/mL、2.96 μg/mL、14.8 μg/mL、29.6 μg/mL;将(-)山莨菪碱和(+)山莨菪碱分别稀释成0.15 μg/mL、0.30 μg/mL、1.5 μg/mL、3.0 μg/mL、15 μg/mL、30 μg/mL;将氢溴酸东莨菪碱稀释成0.1 μg/mL、0.2 μg/mL、1.0 μg/mL、2.0 μg/mL、10 μg/mL、20 μg/mL。将上述标准品溶液经0.22 μm微孔滤膜过滤,备用。因(-)山莨菪碱和(+)山莨菪碱仅旋光性不同,作用性质和强度相似,在计算山莨菪碱含量时,将二者合并计算表示为(±)山莨菪碱。
1.2.4 TAs的HPLC测定 用高效液相色谱仪(安捷伦1200)进行TAs测定。色谱柱为250 mm×4.6 mm×5 μm VWD C18,柱温为25℃,检测波长210 nm。洗脱方式为等度洗脱,流动相为乙腈︰0.085%甲酸=80︰20(V/V),流速1 mL/min,样品进样量为10 μL。
1.2.5 托品烷类生物碱测定方法的建立 利用等度洗脱法对5个托品烷类生物碱进行分离,其中,莨菪碱和阿托品出峰时间基本一致,混合测定不能将两者进行区分,因此将(-)山莨菪碱、(+)山莨菪碱、东莨菪碱和阿托品标品混合测定,莨菪碱标品单独测定。HPLC不能直接定量,需使用回归分析,因此测定不同浓度的4种TAs混合标品和莨菪碱标品峰面积,并绘制各生物碱的标准曲线(即回归分析得到标准曲线,运用标准曲线倒推生物碱含量)。为验证色谱方法的正确性,对交互嫁接后马铃薯和曼陀罗各器官中的TAs进行提取,样品的出峰时间与标准品出峰时间基本一致,则色谱方法正确,反之亦然;为保持试验条件的一致性,根据确定的标准品测定方法测定5种托品烷类生物碱含量。
1.2.6 托品烷类生物碱含量的测定 以曼陀罗为接穗,D1418为砧木进行嫁接的测定马铃薯块茎、曼陀罗茎、叶和果实中5种托品烷类生物碱的含量。以D1418为接穗,曼陀罗为砧木进行嫁接的测定马铃薯茎、叶、叶腋处块茎及曼陀罗根中的5种托品烷类生物碱的含量。
2 结果与分析
2.1 5个托品烷类生物碱的测定方法建立
对5个托品烷类生物碱进行分离测定结果(图 1)显示,4种混合生物碱标品可形成4个独立峰,均有较好的分离度,(-)山莨菪碱、(+)山莨菪碱、东莨菪碱和阿托品的保留时间分别为5.888 min、6.886 min、7.906 min、10.968 min。莨菪碱标品的保留时间为10.868 min。
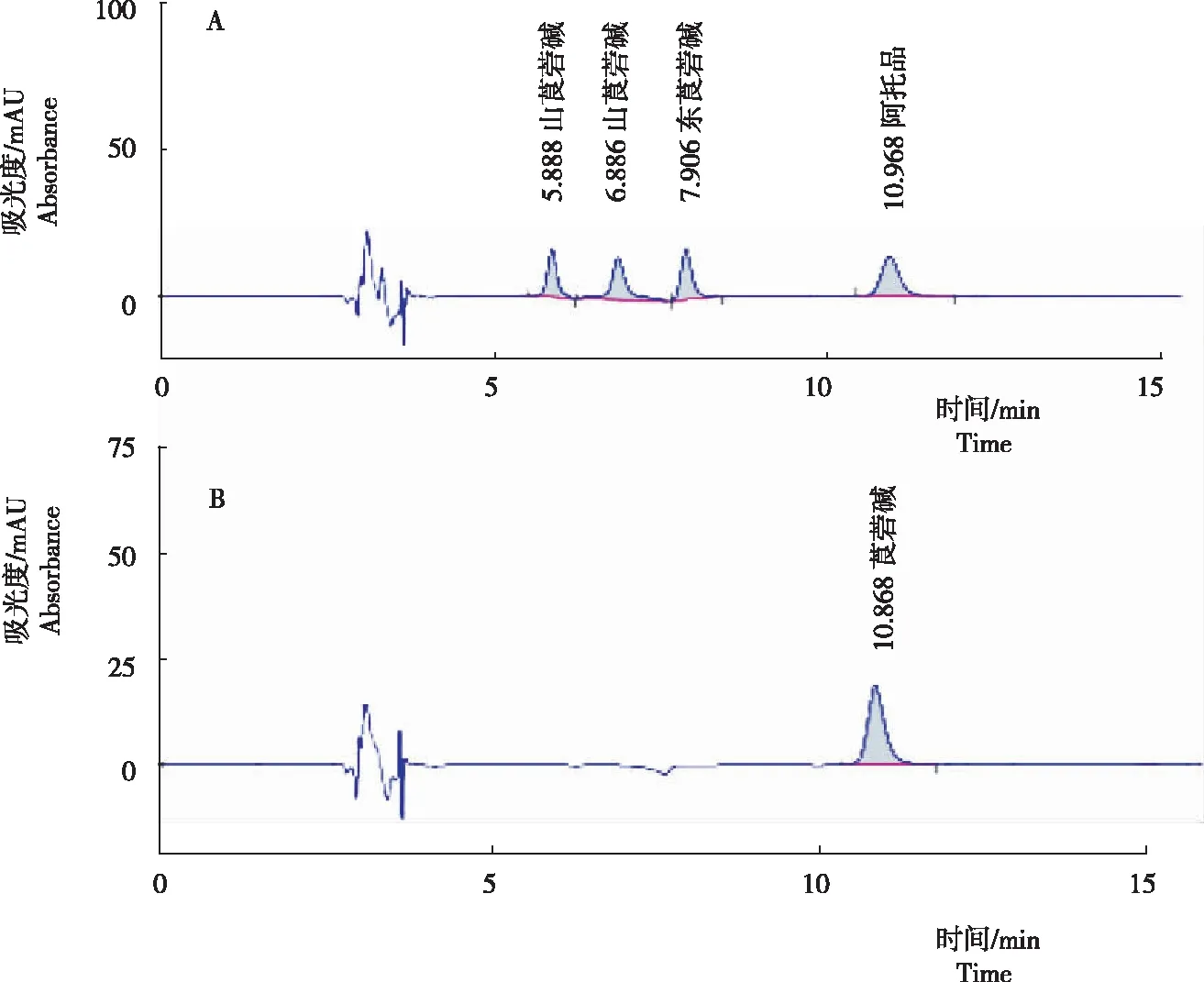
注:A为阿托品、(-)山莨菪碱、(+)山莨菪碱和东莨菪碱混合标品色谱峰图;B为莨菪碱标品色谱峰图。
从图2看出,各托品烷类生物碱标准品的浓度和峰面积均有较好的线性关系,标准曲线R2均大于0.999,标准曲线线性拟合度较好,可用于后续样品测定的计算。
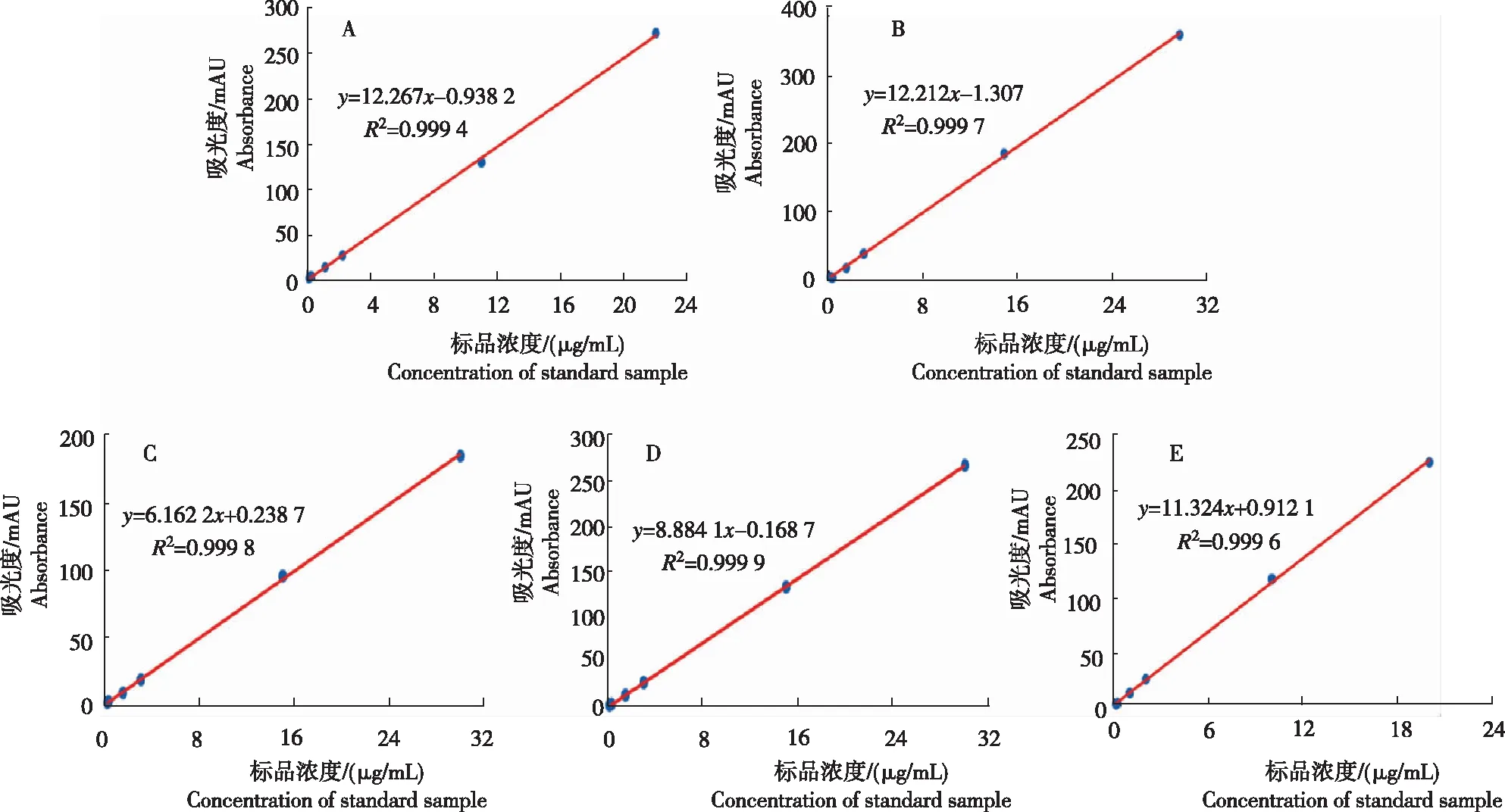
注:A为阿托品,B为莨菪碱,C为(-)山莨菪碱,D为(+)山莨菪碱,E为东莨菪碱。
从图3看出,以曼陀罗果实为例,样品提取和HPLC分离后,能够形成(-)山莨菪碱、(+)山莨菪碱、东莨菪碱和阿托品的特征峰,各成分的保留时间分别为5.817 min、6.771 min、7.751 min和10.791 min,均与混合标准品中各成分的保留时间相符。在曼陀罗果实样品中,莨菪碱能够形成较好的特征峰,保留时间为10.876 min。认为,该方法稳定可靠;在上述色谱条件下,使用HPLC可进行样品中5种TAs的测定。
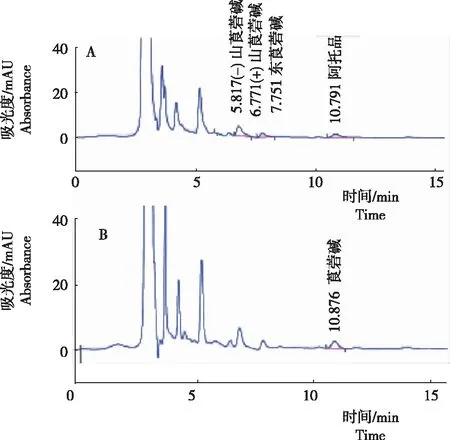
注:A为阿托品、(-)山莨菪碱、(+)山莨菪碱和东莨菪碱色谱峰图;B为莨菪碱色谱峰图。
2.2 曼陀罗/D1418嫁接后各器官托品烷类生物碱含量
对以曼陀罗为接穗,D1418为砧木进行嫁接的曼陀罗茎、叶和果实及马铃薯块茎中5种药用生物碱含量的测定结果(表1)显示,在砧木和接穗不同器官中,均可检测到5种托品烷类生物碱。曼陀罗接穗各器官中,TAs生物碱含量在果实中最高,为121.77 μg/g;叶片中最低,为35.61 μg/g。曼陀罗接穗各器官中,均是(±)山莨菪碱含量最高,为29.15~87.98 μg/g;东莨菪碱含量最低,为1.02~9.07 μg/g。在马铃薯砧木所形成的块茎中,5种生物碱含量总和仅为8.16 μg/g,其中莨菪碱含量最高,为2.87 μg/g;阿托品含量最低,为1.50 μg/g。曼陀罗茎和马铃薯块茎的(±)山莨菪碱含量间差异显著。表明,曼陀罗果实中托品烷类生物碱含量最高;以曼陀罗为接穗,马铃薯为砧木所形成的块茎中能够富集一定量的托品烷类生物碱,说明托品烷类生物碱能够在维管组织中从上至下运输。
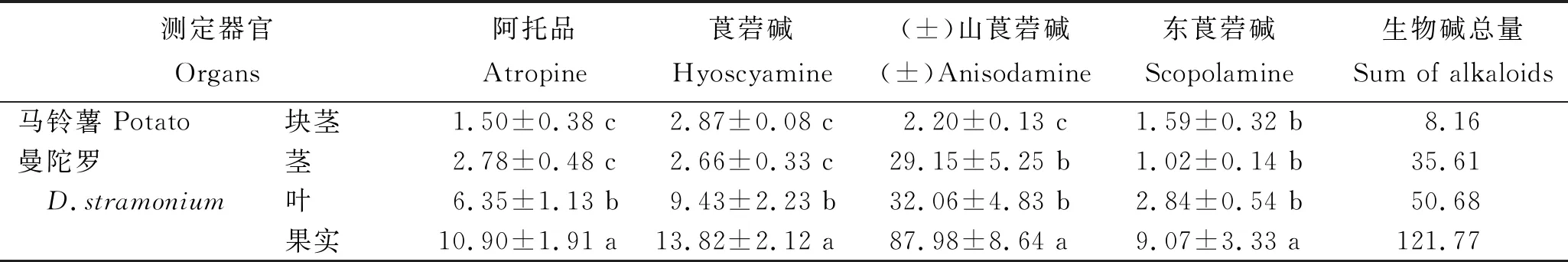
表1 曼陀罗/D1418嫁接后不同器官中5种托品烷类生物碱含量
2.3 D1418/曼陀罗嫁接后各器官托品烷类生物碱含量
对以D1418为接穗,曼陀罗为砧木进行嫁接后马铃薯的茎、叶、叶腋处块茎及曼陀罗根中5种托品烷类生物碱含量的测定结果(表2)显示,砧木和接穗器官中均存在5种托品烷类生物碱。生物碱总量以马铃薯叶片中的最高,为214.23 μg/g;曼陀罗砧木的根中较低,为34.13 μg/g;马铃薯叶腋处块茎的最低,为13.22 μg/g。4个器官中均是莨菪碱含量最高,阿托品含量其次,(±)山莨菪碱和东莨菪碱含量较低。表明,曼陀罗根系中托品烷类生物碱能够在维管组织中从下至上运输,并在马铃薯接穗茎叶中富集,也能在叶腋处的气生薯中富集。
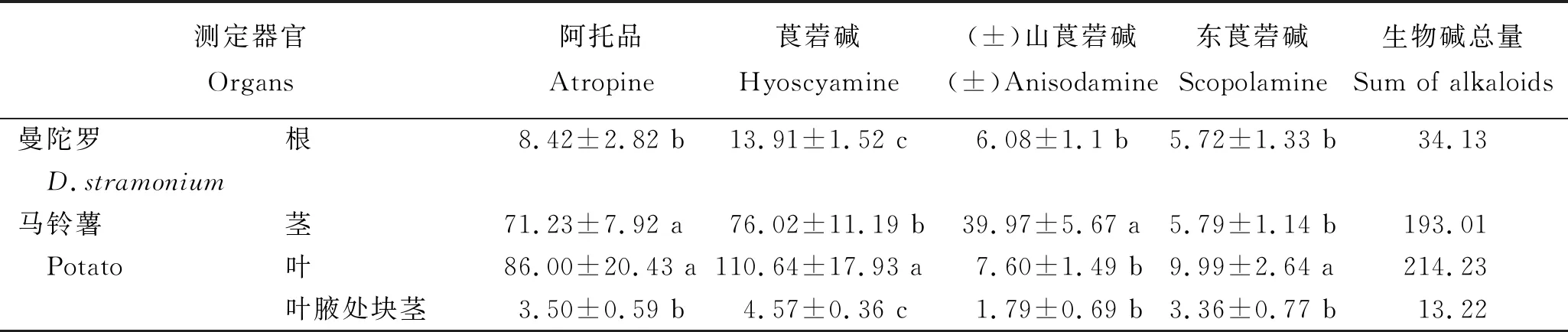
表2 D1418/曼陀罗嫁接后不同器官中5种托品烷类生物碱含量
3 讨论
3.1 交互嫁接后曼陀罗不同器官中托品烷类生物碱的积累规律
曼陀罗中主要的生物碱,如阿托品、莨菪碱、山莨菪碱和东莨菪碱等均在根系中进行生物合成[28-31],并通过维管组织运输到地上部分。D1418/曼陀罗嫁接组合中,马铃薯接穗茎叶的托品烷生物碱含量高于曼陀罗/D1418嫁接中的曼陀罗茎叶。曼陀罗砧木的根系能够不断合成TAs,是生物碱合成的“源”;而曼陀罗砧木的根系所产生的TAs,从维管组织大量转运到地上部马铃薯接穗茎叶中,使马铃薯接穗成为“库”;当以马铃薯为砧木时,失去了TAs合成的“源”,因此曼陀罗接穗中的生物碱含量低于马铃薯接穗。在曼陀罗/D1418嫁接组合中,马铃薯砧木的块茎中也积累了一定量的TAs,说明TAs在曼陀罗茎中能够自上而下运输。
在TAs生物合成途径的下游,莨菪碱和阿托品是(±)山莨菪碱的合成前体,而(±)山莨菪碱是东莨菪碱的合成前体,这2步酶促反应都受到同一限速酶H6H的催化[15,32]。因此,在曼陀罗中莨菪碱和阿托品的含量高于东莨菪碱。曼陀罗各组织器官中的东莨菪碱含量均较低,该现象与TAs生物合成过程相符。
曼陀罗根系中莨菪碱是5种生物碱的主要组成成分,而在曼陀罗地上部组织中(±)山莨菪碱是主要的组成成分,说明曼陀罗茎叶和果实可能特异性地积累(±)山莨菪碱,同时在茎叶和果实中也可能自主合成(±)山莨菪碱。
3.2 马铃薯接穗对曼陀罗中托品烷类生物碱的积累作用
通过嫁接能够使砧木和接穗之间的营养物质产生相互交流[24]。马铃薯和曼陀罗都是茄科作物,马铃薯的主要生物碱是糖苷生物碱[33],曼陀罗的主要生物碱是托品烷类生物碱[2,34]。D1418/曼陀罗嫁接后,马铃薯茎叶中也能够积累大量的TAs,说明马铃薯维管组织也能够转运和富集曼陀罗的TAs。同时,马铃薯接穗和曼陀罗接穗茎叶中的托品烷类生物碱主要组成成分不同,说明马铃薯维管组织积累托品烷类生物碱的具体机制可能与曼陀罗不同。马铃薯和曼陀罗交互嫁接中,在马铃薯的地下块茎及地上部叶腋气生薯中均检测到曼陀罗的TAs,该结果为利用马铃薯块茎来特异性积累TAs奠定了基础。另外,在马铃薯地上或地下块茎中莨菪碱均是含量最高的TAs,说明块茎组织可能有偏好性地积累托品烷类生物碱。
4 结论
托品烷类生物碱通过维管组织既能从上至下运输,也能从下至上运输。以曼陀罗为接穗,马铃薯为砧木所形成的块茎中能够富集一定量的托品烷类生物碱;以曼陀罗为砧木,马铃薯为接穗时,托品烷类生物碱能在马铃薯接穗茎叶中富集,同时也能在马铃薯叶腋处的气生薯中富集。
猜你喜欢
中国马铃薯(2022年2期)2022-07-05
中国药学药品知识仓库(2022年7期)2022-05-10
中草药(2022年5期)2022-03-03
源流(2021年8期)2021-09-27
中国现代医生(2020年9期)2020-06-03
分析化学(2014年10期)2014-10-24
中国民族民间医药·下半月(2014年4期)2014-09-26
少年科学(2014年1期)2014-02-24
中国社区医师(2009年16期)2009-09-10
中学生百科·悦青春(2009年2期)2009-03-03
