
三江源区高寒坡地退化植物群落多样性和地上生物量沿海拔梯度的变化特征
2022-05-26 10:02:22王采娥王文银李子好白彦福王彦龙施建军龙瑞军王晓丽马玉寿尚占环
生态学报 2022年9期
王采娥,黄 梅,王文银,李子好,张 涛,2,马 林,白彦福,王彦龙,施建军,龙瑞军,刘 玉,王晓丽,马玉寿,尚占环,*
1 兰州大学生命科学学院,草地农业生态系统国家重点实验室,兰州 730000 2 中国科学院西北生态环境资源研究院,兰州 730000 3 青海大学畜牧兽医科学院,西宁 810006 4 西北农林科技大学水土保持研究所,中国科学院水利部水土保持研究所,黄土高原土壤侵蚀与旱地农业国家重点实验室,杨凌 712100
生物多样性沿环境梯度的变化以及与生产力的相互关系是生物多样性研究的一个重要内容[1]。物种多样性是研究草地生态系统多样性及结构的重要方面和重要指标,有助于理解草地生态系统的稳定性维持机制[1—2],地上生物量能够反映植被特征和生产力[3],是草地生态系统稳定的基础[4]。植物物种多样性是维持生态系统结构、功能和稳定性的基础[5],植物群落多样性和生物量的深入研究对于多样性保护、生产力维持以及生态环境恢复具有极其重要的意义[6]。因此,草地生态系统物种多样性与生物量关系已成为生态学领域最受关注的焦点问题之一[7]。
山地植物群落沿环境梯度的变化一直是生态学家关注的问题,在山地生态学研究中有着重要价值[8]。海拔和地形变化能够直接影响环境因子,从而导致山地植被物种组成和群落结构发生变化[9—11]。海拔变化能够引起温度、湿度和太阳辐射等环境因子的变化,从而影响植物生物量及多样性分布格局[12]。海拔梯度上,国内外学者主要集中在草地物种多样性、地上生物量及相互关系的研究[6,13—14],邓清月等研究表明灌木群落多样性与海拔变化呈“单峰”格局,草本群落呈波动变化[15];杨元合等研究得出物种丰富度与海拔变化呈负相关[16];牛钰杰等研究得出物种丰富度在阳坡与海拔成正相关,在阴坡呈单峰分布格局[17];王飞等研究表明不同坡向灌丛和草甸群落多样性沿海拔变化呈“中间高度膨胀”[18]。刘哲等在高寒山坡草甸研究发现,地上生物量随海拔升高线性降低[6];王金兰等和罗黎鸣等研究得出灌丛和灌下草本生物量随海拔升高先增加后减少,呈单峰格局[19—20]。
近年来,由于气候变化[21]和人类活动加剧,导致青藏高原高寒草甸生态系统遭到破坏,生物多样性减少,初级生产力下降,高寒草甸面临严峻的退化态势[22]。植被恢复是解决青藏高原生态系统退化的有效生物手段之一,如果植被具有较大生物量和多样性,其能够更好的抑制侵蚀和剥蚀,植被空间分布格局对于坡地水土流失治理方面发挥重要作用[23—24]。当前青藏高原关于植物群落多样性和地上生物量的研究多集中在正常的高寒山地生态系统及高寒草甸[6,25—26],只有少数是以高寒灌丛群落为研究对象。因此,研究退化高寒坡地草甸和灌丛植物群落物种多样性和地上生物量的分布格局对高寒山地生态系统的保护与恢复具有重要参考价值。
山地退化态势下,了解坡地退化高寒草甸和灌丛植物多样性与地上生物量沿海拔梯度的变化规律及分布格局,有助于认识生态环境退化如何影响山地生态系统结构和功能改变,以及为山地植被恢复提供科学参考。实地调查中发现,三江源区黑土滩退化草地蔓延至山坡,形成“黑土山”造成大范围的山地退化态势。目前已经出现大量坡地退化,给三江源区生态建设带来新的挑战,因此需要深入认识这种坡地植被退化引起的高寒山地生态系统变化[27],从而为进一步的恢复治理提供理论依据和科技支撑。因此,本研究以青海省果洛藏族自治州达日县典型的高寒坡地退化植被为研究区,选取高寒草甸和高寒灌丛两种群落类型作为研究对象,通过实地群落学调查分析坡地退化高寒草甸和灌丛植物物种组成、物种多样性及地上生物量对海拔变化的响应,揭示退化高寒坡地不同海拔梯度上植物多样性和地上生物量分布变化的规律,为三江源区高寒坡地退化植被的生态恢复治理、保护提供参考。
1 材料与方法
1.1 研究区概况
达日县位于青藏高原青海省的东南部,果洛州的南部,地形呈蝴蝶状,地势西北高,东南低(图1)。窝赛乡地处达日县域东部达日县城南侧,地理坐标为99.43°E,33.40°N,平均海拔4000 m 以上,属于大陆性高原气候,年均气温-0.9℃,最高气温20.1℃,最低气温 -27.6℃,年降水量544.7 mm,气象站20 cm专用口径水面蒸发量为1219 mm,一年没有明显四季区分,只有冷暖两季,冷季漫长,暖季短暂,无绝对的无霜期[28]。达日县土壤质地粗劣,多为砾质轻壤-砾质中壤,pH值6.5—8.0。主要有三种土壤类型:高山草甸土,分布在海拔4000—4600 m的滩地和坡地;高山沼泽土,见于海拔4000 m以上的山谷,低洼地等;高山寒漠土,分布在海拔4600 m以上的高山地带[29—30]。
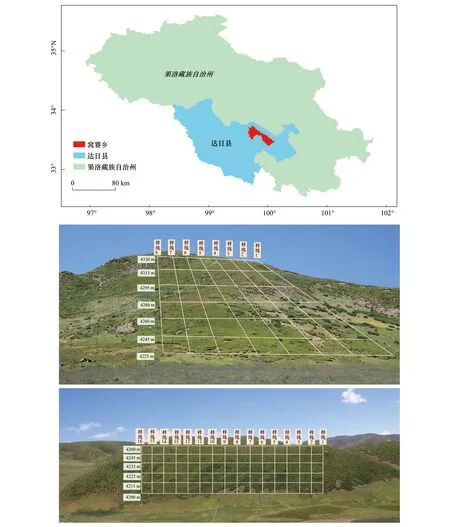
图1 研究区域和样地设置Fig.1 Location of study area and sample sites
达日县植被分布具有明显的垂直分布规律,植被类型主要有高寒草甸、沼泽草甸、灌丛草甸、高寒稀疏植被等。极大多植被类型为高寒草甸,高寒灌丛分布面积较小。高寒稀疏植被位于海拔4700—4800 m;高寒草甸位于海拔3900—4700 m,分布在山坡阳面以及山顶;高寒灌丛位于海拔4000—4350 m左右,分布在阴坡,半阴半阳坡以及河谷滩地较多。优势种主要有高山嵩草(Kobresiapygmaea)、矮生嵩草(Kobresiahumilis)、线叶嵩草(Kobresiacapillifolia)、山生柳(Salixoritrepha)、金露梅(Potentillafruticosa)、高山绣线菊(Spiraeaalpina)等,而伴生种主要有星状雪兔子(Saussureastella)、条叶银莲花(Anemonecoelestina)、珠芽蓼(Polygonumviviparum)、钉柱委陵菜(Potentillasaundersiana)、毛果婆婆纳(Veronicaeriogyne)和肉果草(Lanceatibetica)等。
1.2 研究方法
植物群落调查时间为2019年7月—8月,按照方精云等[31]发表的研究方法,对研究区植物进行群落调查。首先,以“典型性、代表性、一致性”为原则,在研究区选取样地,根据群落组成,选取空间距离较近的以草甸植物为主(处于阳坡)和以灌丛植物为主(处于阴坡)的两个坡地。同时使用GPS记录实验样地地理位置和海拔。
草甸坡地调查样线和样方:草甸坡地从坡底到坡顶设置样线,共计8条样线,沿着样线从坡底到坡顶在海拔4225、4245、4260、4280、4295、4315 m及4330 m处做50 cm×50 cm标准样方,共56个。
灌丛坡地调查样线和样方:灌木和草本分层调查,从坡底到坡顶设置样线,共计16条样线,沿着样线在海拔4200、4215、4225、4235、4245 m及4260 m处做2 m×2 m标准样方。同时每个灌木样方内设置草本层标准样方50 cm×50 cm 3个,2 m×2 m样方89个,草本层样方共172个。
植物群落学调查:记录群落中物种种类、物种高度、植株个数、盖度等数据。物种高度:每一种植物随机测量10次,盖度通过自制的样方框用针刺法获得(草甸坡地采用50 cm×50 cm的样方框,灌丛坡地采用2 m×2 m样方框)。将实验样地不能够识别的植物采摘下来,并拍生境图片包括多角度的叶片、茎、根以及花朵,并加以记录,带回实验室鉴定,鉴定标准依据《中国植物志》。草甸坡地地上生物量的获取:将草本层样方内植物齐地刈割,称重,装进信封袋,后期测定地上生物量。灌丛坡地地上生物量的获取:灌丛坡地地上生物量包括两部分,一部分为灌木层地上生物量,另一部分为灌下草本层地上生物量。灌木只采取新萌发生长的枝叶,称重,装入信封袋,后期测定灌木层地上生物量,将灌下草本层齐地刈割,称重,装进信封袋,作为灌丛坡地草本层地上生物量,灌丛坡地灌木层地上生物量和灌丛坡地草本层地上生物量之和作为灌丛坡地地上生物量。将所采取的地上生物量样品放入烘箱,在105℃温度条件下杀青30 min,紧接着在65℃温度条件下烘48 h及以上直至达到恒重并称重记录,记作地上生物量。
1.3 数据处理分析
群落多样性根据方精云等[31]的植物群落多样性测度方法,选取以下4个指数,能够表示群落中所含物种多少以及群落中物种分布,避免单一选取造成的不足。因此采用Shannon-wiener指数、Simpson指数、Margalef指数和Pielou指数分析植被物种多样性[32—33]。
Shannon-wiener指数:
(1)
Simpson优势集中性指数:
(2)
Margalef丰富度指数:
(3)
Pielou均匀度指数:
(4)
式中,S为物种总数,Pi为物种i植物个体数占群落总个体数比例,N为样地所有个体数总和。
数据处理采用Excel 2019和SPSS 22.0处理相关数据以及Origin 2018进行作图分析。采用单因素方差分析(One-way ANOVA)对物种多样性指数进行方差分析,采用Duncan′s 法进行差异显著性检验,显著性水平P<0.05,在SPSS 22.0中完成统计分析。利用Origin 2018进行线性和非线性回归分析:(1)灌丛坡地和草甸坡地地上生物量、Shannon-wiener指数、Simpson指数、Pielou指数、Margalef指数与海拔之间的关系。(2)灌丛坡地与草甸坡地地上生物量与Shannon-wiener指数、Simpson指数、Pielou指数及Margalef指数的关系,利用曲线估计进行拟合,找出有意义且拟合度最优的曲线和方程。
2 结果与分析
2.1 坡地退化植物群落特征
2.1.1坡地退化高寒草甸植物群落特征
高寒草甸调查56个标准样方中样方内物种数变化范围为9—18种,植株平均高度2.43 cm,共调查到84种植物,隶属于66属,39科,其中杂类草植物较多,共76种,隶属于58属,36科,豆科、禾本科和莎草科植物分别有4、2、2种。杂类草中菊科(Asteraceae)占比最多(10种),其次毛茛科(Ranunculaceae),有9种;龙胆科(Gentianaceae),有6种植物;蓼科(Polygonaceae)、蔷薇科(Rosaceae)和玄参科(Scrophulariaceae)分别为5、5、3种植物。调查样地内植物有毛果婆婆纳、钉柱委陵菜、矮火绒草(Leontopodiumnanum)、肉果草、隐瓣蝇子草(Silenegonosperma)、驴蹄草(Calthapalustris)和圆穗蓼等。随着海拔升高,不同样线上物种数差异较大。样线1物种种类数目在不同样方内差异较大,海拔4260 m处,物种数只有4种,但是在海拔4330 m处,物种数达到17种,造成差异的主要原因是样线1处于两块坡地的交界处,易发生侵蚀,植被退化严重,部分采样点植被盖度低于30%,样线2在海拔4223 m植被盖度低于15%。随着海拔升高,植被盖度增加(图2),样方内物种种类数目变化较大,主要是因为有些样方处于秃斑块内,或者鼠洞分布较密集的地方和裸地,退化程度相对较严重,植被盖度低,植物群落物种多样性低。
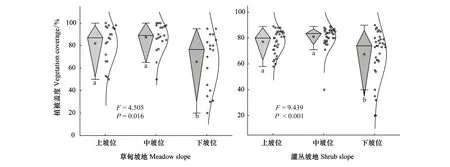
图2 坡地退化植被不同样线上植被盖度沿海拔梯度变化规律Fig.2 The vegetation coverage on different lines in degraded mountain areas varies with altitude
草甸坡地下坡位植被盖度显著低于中坡位和上坡位(P<0.05)(图2),上坡位和中坡位植被盖度差异不显著。由表1可知,不同海拔植被盖度差异不显著。不同样方间植被盖度空间异质性较大。海拔4245 m处,同一条样带八个样方中植被盖度达到95%,最小仅20%。

表1 坡地退化高寒草甸样地详细信息及调查结果(平均值±标准误)Table 1 Detailed information and survey results of degraded slopes in alpine meadow (Mean±SE)
2.1.2坡地退化高寒灌丛植物群落特征
高寒灌丛调查89个标准样方(2 m×2 m)中样方内物种数变化范围为18—24种,共调查到85种植物,隶属于68属、29科。沿海拔梯度植物群落主要有金露梅群落和山生柳群落,海拔4245—4260 m处,主要是高山嵩草和矮生嵩草草甸。海拔4200—4260 m处,灌丛植物仅有三种,山生柳、高山绣线菊和金露梅,三种灌木植物植株高度分别是55.89 cm、37.67 cm和20 cm。调查样方内杂类草居多,有75种,隶属于58属、26科。豆科、禾本科、莎草科植物分别有4、4、2种。其中菊科最多,有14种,其次毛茛科和龙胆科,均有9种植物。实地调查出现频次较少的植物主要有蓬子菜(Galiumverum)、单子麻黄(Ephedramonosperma)、垫状棱子芹(Pleurospermumhedinii)、双花堇菜(Violabiflora)以及沼生柳叶菜(Epilobiumpalustre)等。海拔4245—4260 m处,灌丛植物分布较少,主要分布草本层植物。植被覆盖度整体较好,海拔4200 m和4245 m处,不同样线间植被盖度变化较大,根据调查,主要是由于人类干扰和山体发生严重侵蚀和沟蚀,造成植被退化、啮齿动物泛滥以及坡地秃斑化等现象。
灌丛坡地下坡位植被盖度显著低于中坡位和上坡位(P<0.001)。中坡位和上坡位植被盖度差异不显著。由表2可知,灌丛坡地在海拔4200 m处植被盖度显著低于其他坡位植被盖度(P<0.05)。下坡位植被盖度低;4225—4235 m处,不同样方植被盖度差异较小;海拔4235m处草地退化严重,有些样方植被盖度仅达到40%。植被盖度空间异质性较大(图2)。
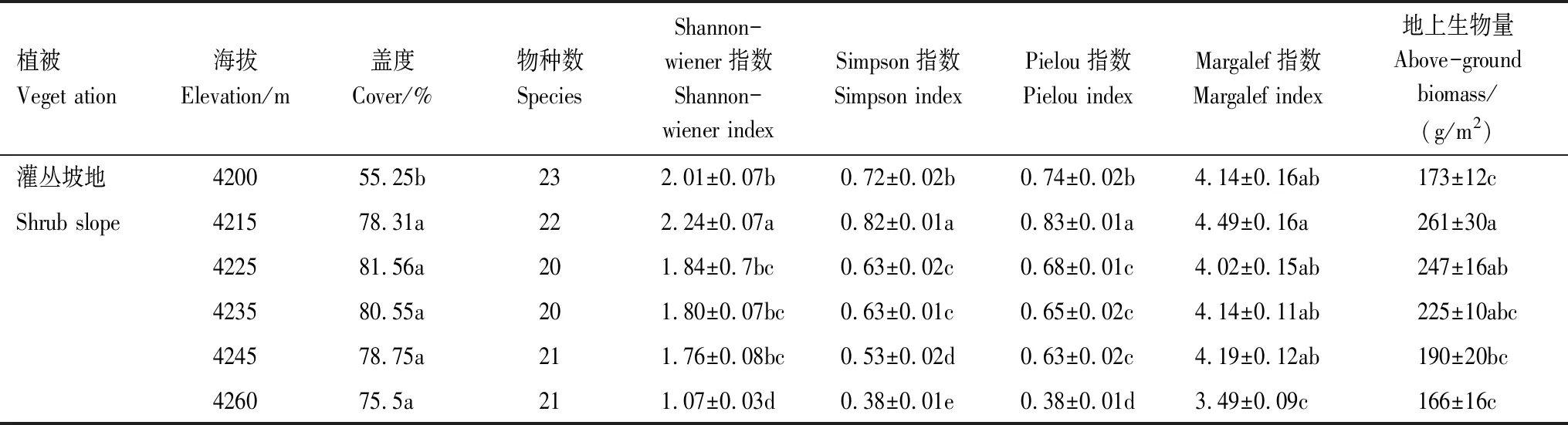
表2 坡地退化高寒灌丛样地详细信息及调查结果(平均值±标准误)Table 2 Detailed information and survey results of alpine shrub degraded slopes (mean ± standard error)
高寒草甸和高寒灌丛相比较,高寒灌丛群落调查样方内出现的物种数要比高寒草甸群落出现的物种数高达6种以上,高寒草甸84种植物隶属于39科,高寒灌丛85种植物隶属于29科。灌丛坡地地上生物量也要高于草甸坡地地上生物量。Shannon-wiener指数和Margalef指数在高寒灌丛群落中也大于高寒草甸群落,说明高寒灌丛退化较高寒草甸轻,物种多样性高,物种较丰富。
2.1.3坡地高寒草甸、灌丛不同坡位物种组成相似性
草甸坡地上坡位、中坡位与下坡位之间物种组成相似度较高,灌丛坡地在上坡位、中坡位和下坡位之间物种组成差异性较大(图3)。灌丛坡地不同坡位差异性大可能是由于灌丛植物的存在,使灌下草本层植被分布不同,灌丛坡地的坡顶灌木生长较少,坡中生长着山生柳、绣线菊,坡下生长着金露梅,造成灌下草本层植物生长微环境的差异,从而使物种组成存在差异。
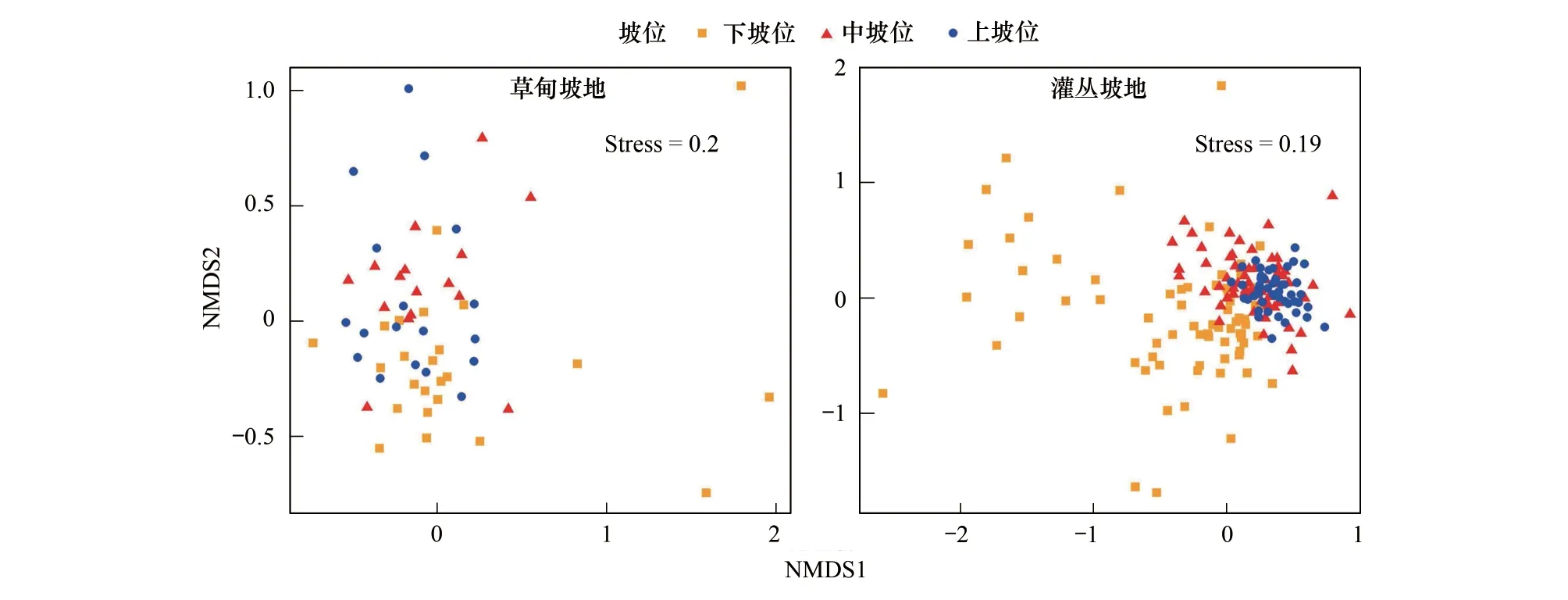
图3 高寒草甸、灌丛坡地不同坡位NMDS分析Fig.3 NMDS analysis of different slope positions of alpine meadow and shrub slope NMDS:非度量多维尺度分析 Non-metric multidimensional scaling
2.2 坡地退化植物群落物种多样性沿海拔梯度的变化特征
2.2.1坡地退化高寒草甸植物群落多样性沿海拔梯度的变化特征
教师对学生的评价包括书面评价和口头评价,书面评价的优点在于经过深思熟虑,因而正式而切实,口头评价的优点则在于它的自由度和及时性。每个学生都渴望得到他人的肯定,初中生正处在自我意识觉醒的重要时期,教师评价的导向作用不容小觑。因此教师必须牢记评价分层的重要性,善于鼓励、及时提醒,以正确而适时的评价为学生指明方向。
由图4和表1可知,高寒草甸植物群落Shannon-wiener指数、Simpson指数和Pielou指数的最大值均处于中间海拔高度,呈峰值明显的单峰曲线。二次曲线回归分析中,海拔与Shannon-wiener指数、Simpson指数最优拟合曲线均为二项式,显著相关(P<0.05)(图4、图5)。海拔与Margalef指数最优拟合为线性回归,显著相关(P<0.05)(图4)。但是Pielou指数二次回归曲线拟合虽然呈单峰曲线(图4),但是P>0.05,不能够解释随着海拔升高Pielou指数先增大后减小的变化趋势。二次回归方程曲线拟合解释度较高,说明海拔对香农指数、辛普森指数和丰富度指数具有显著影响,表明坡地退化植被在中间海拔梯度具有较高的物种多样性。Margalef指数随海拔升高呈线性增加,说明植物群落在高海拔处丰富度较高。同一海拔高度不同样方间物种多样性空间异质性较大。

图4 坡地高寒草甸和高寒灌丛物种多样性随海拔梯度的变化规律Fig.4 Changes of species diversity in alpine meadow and alpine shrub communities with elevation gradient
2.2.2坡地退化高寒灌丛植物群落多样性沿海拔梯度的变化特征
高寒灌丛植物群落物种多样性随海拔梯度变化呈现出一定变化规律。如图4、图5所示,二次曲线回归分析中,Shannon-wiener指数、Simpson指数、Pielou指数和Margalef指数均随海拔升高先增大后减小,最大值在海拔4215 m,最小值在4260 m(表2),呈单峰曲线,海拔与多样性指数最优拟合为二项式,显著相关(P<0.05)。海拔与Margalef指数成单峰曲线趋势,但是没有显著性意义。多元回归方程解释度达到90%以上,说明海拔是影响灌丛物种多样性指数的关键因素。同一海拔不同样方内物种多样性空间异质性较大。
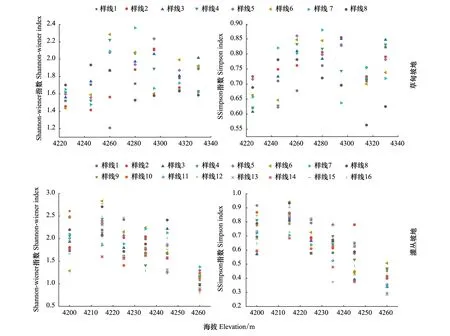
图5 坡地高寒草甸和高寒灌丛物种多样性沿海拔梯度的变化规律Fig.5 Changes of species diversity in alpine meadow and alpine shrub slopes along elevation gradient
高寒草甸物种多样性沿海拔变化规律呈“单峰”曲线,高寒灌丛物种多样性沿海拔梯度的变化规律呈“偏单峰”曲线,高寒草甸Shannon-wiener指数1.94,出现在海拔4280 m处,高寒灌丛Shannon-wiener指数最大值2.24,出现在海拔4215 m处;高寒草甸Margalef指数最大值(2.54)在海拔4295 m和4330 m,高寒灌丛Margalef指数最大值(4.49)在海拔4215 m。随着海拔升高,物种丰富度先增大后减小,坡顶物种丰富度最低。
2.3 坡地退化植物群落地上生物量沿海拔梯度的变化
2.3.1坡地退化高寒草甸地上生物量沿海拔梯度的变化
草甸坡地地上生物量随海拔变化如图6所示,地上生物量随着海拔升高先增加后减小,地上生物量最大值在4280 m,最小值在4220 m(表1)。二次曲线回归分析中,最优拟合曲线为二项式(R2=0.88,P<0.05)。因此,坡地草甸植物群落地上生物量随海拔变化特征符合中间高两头低的“单峰”趋势。较低海拔段,植物生长热量比较充足,但土层相对较薄,侵蚀较严重,土壤沙质化,持水能力较差,以及由于人类干扰活动,导致地上生物量较低。
2.3.2坡地退化高寒灌丛地上生物量沿海拔梯度的变化
灌丛坡地地上生物量随海拔升高先增加后减少的变化趋势(图6)。地上生物量最大值在海拔4215 m,最小值在4260 m左右(表2),差异显著(P<0.05)。野外调查结果表明,该地区由于放牧活动,造成植被覆盖度较低。灌丛坡地地上生物量包括草本层地上生物量和灌木层地上生物量两部分,灌丛坡地地上生物量随海拔升高呈“单峰”曲线,主要原因是坡底侵蚀严重,植被盖度低,灌丛植物生长较少,故地上生物量较低;山坡中间部位水热条件较好以及由于灌丛植物的存在,阻碍了家畜践踏与啃食,地上生物量较高,而坡顶由于受到风蚀影响,土层较薄,水热条件差,灌木植物生长较少,故地上生物量与中间坡位相比较低。
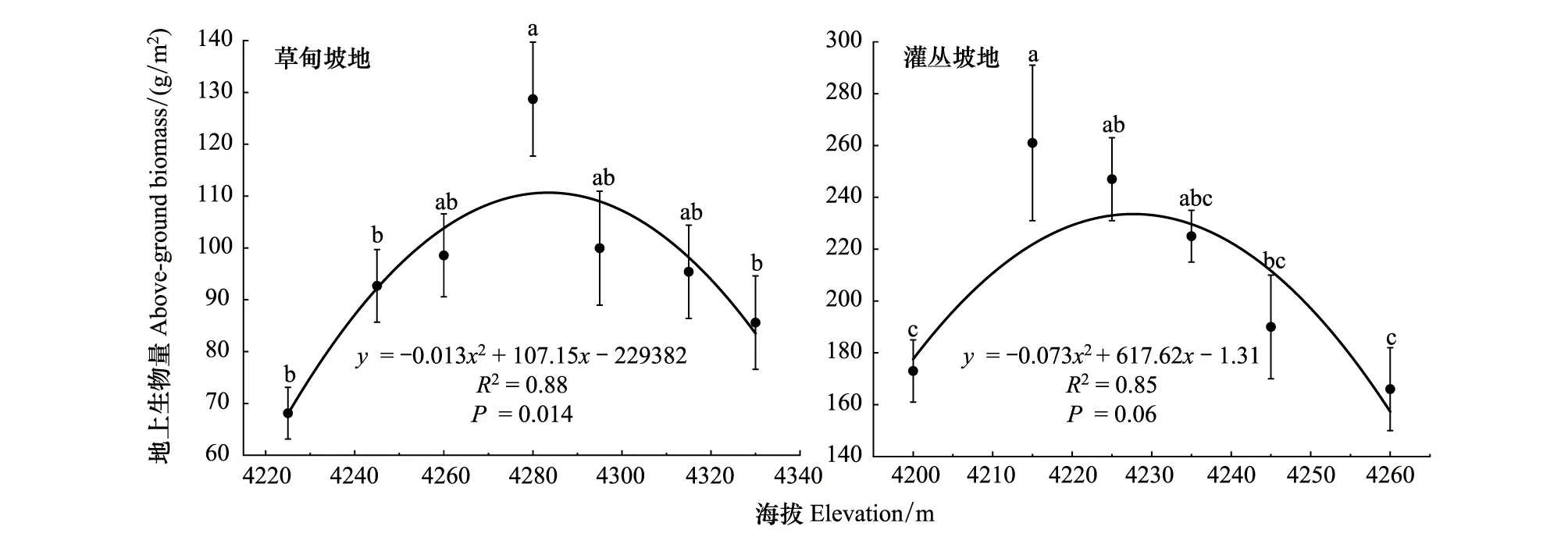
图6 坡地高寒草甸和高寒灌丛地上生物量随海拔梯度的变化规律Fig.6 Changes of above-ground biomass in alpine meadow and alpine shrub with elevation gradient
2.4 坡地退化植物群落物种多样性与地上生物量的关系
回归分析高寒草甸地上生物量和物种多样性(图7),最优拟合均为一次函数方程,显著相关(P<0.05)。Shannon-wiener指数和Simpson指数与地上生物量呈线性正相关关系(P<0.05),Pielou指数随地上生物量变化呈线性增加趋势。地上生物量分别能够解释Shannon-wiener指数和Simpson指数变化的70%和80%,而对Pielou指数和Margalef指数解释度小于50%,且没有显著相关性(P>0.05),说明地上生物量对Pielou指数和Margalef指数没影响,不能用来判断生产力水平。
2.4.2坡地退化高寒灌丛物种多样性与地上生物量的关系
高寒灌丛地上生物量与Shannon-wiener指数、Simpson指数、Pielou指数和Margalef指数曲线最优拟合为一次函数和二次函数(图7)。Shannon-wiener指数和Simpson指数随地上生物量变化呈线性增加趋势,地上生物量对Shannon-wiener指数和Simpson指数的解释度达到60%。Pielou指数与地上生物量之间呈二次函数相关,即随着地上生物量增加Pielou指数先减小再增大 (P=0.06),地上生物量变化对Pielou指数的解释度达到90%,Margalef指数随地上生物量增加而增大的变化趋势。

图7 坡地高寒草甸和高寒灌丛地上生物量与物种多样性的关系Fig.7 Relationship between above-ground biomass and species diversity in alpine meadow and alpine shrub communities
从图7可知坡地退化高寒草甸和高寒灌丛地上生物量与香农指数和辛普森指数之间均为线性正相关关系,即随着海拔升高,物种多样性成线性增加。均匀度指数在草甸和灌丛坡地之间不同,草甸坡地均匀度指数与生物量之间成正相关,灌丛坡地均匀度指数与地上生物量之间呈二次函数相关,说明均匀度受到植被类型及环境的影响。高寒草甸和高寒灌丛坡地的物种丰富度与地上生物量没有相关性,可能是因为坡地退化较严重,物种多样性受到严重影响。因此,结合群落调查数据可以得出多样性指数和地上生物量呈正相关。
3 讨论
3.1 坡地退化植物群落特征
三江源区草地退化严重,窝赛乡小流域处于中度退化状态,治理恢复难度大。调查样方中,草甸坡地共计84种植物,隶属于66属39科,灌丛坡地85种植物,隶属于68属29科。灌下草本层物种数和草甸坡地物种数沿海拔变化不同,草甸坡地物种数随海拔升高先增加后减少,灌下物种数随海拔升高而减少,主要是因为坡底和坡中草本层植物受到灌丛庇护。研究表明,在高山环境下,护士植物(nurse plant)对微生境的改善能频繁减弱环境压力对环境的影响[34]。随着海拔升高,灌丛坡地物种数减少,可能是海拔相对较高,风蚀严重,水热条件差,不利于灌木植物生长,易于草本植物生存,而草本层没有灌丛庇护,故多样性较低[15],由此推断,灌丛植物在高寒草甸生态系统结构和生产功能的稳定性中扮演重要角色。张蕊等[35]2013年在达日县窝赛乡植物群落多样性研究表明,天然草地Shannon-wiener指数、Simpson指数和Pielou指数均最大,“黑土滩”次之,退化人工草地最低。本研究高寒坡地植物群落多样性均小于张蕊等[35]研究结果,表明窝赛乡小流域内坡地植被仍在持续退化。
青海省达日县窝赛乡小流域内高寒坡地下坡位退化严重,植被覆盖度较低,遍布大面积秃斑块,啮齿动物泛滥,生物多样性低,这是草地严重退化的表现。由于人类活动在下坡位较频繁以及家畜践踏和啃食影响等共同作用,造成下坡位植被盖度降低,水土流失严重,土壤贫瘠,土地斑块化,石头分布密集,生物多样性降低等现象。上坡位表层土壤等被泥水冲蚀聚集到下坡位,使表层土壤被冲蚀,深层岩石裸露出来,土壤沙质化,从而不适宜植被生长发育;同时由于坡度较低的区域适合人类活动以及啮齿动物的生存,从而加重草地退化和环境恶化;下坡位遍布大量毒杂草,可食牧草匮乏,土壤植被恢复慢,不能够满足人类活动,于是人类活动向草地植被良好的区域扩展,活动范围扩大,造成其他坡位草地生态退化严重,从而导致草地退化由山坡底部向山坡其他部位蔓延。随着退化时间及退化强度增加,毒杂草逐渐向中坡位和上坡位侵略,成为优势种,从而造成大面积坡地发生退化。
3.2 坡地退化植物群落物种多样性沿海拔梯度的分布规律
植物群落多样性的垂直梯度格局受到植被演化、物种进化、地理差异及环境因子等多重因素共同影响[14]。海拔已被证明对各种生态系统的物种多样性和物种组成起着重要的控制作用[11],海拔变化(温湿度和光照等多种环境因子)影响山地生态系统的生态-水文过程[36],影响山地物种组成和群落结构,能够使物种多样性与海拔梯度变化呈现不一致的变化规律[15]。植物群落多样性是研究群落组成的重要指标[1,17,37],能够反映出植物群落特性、生态学特性、分布状况及其对环境的适应性[1,38]。
青藏高原研究表明,物种多样性随海拔变化规律最常见的模式是单峰曲线[6—7,20,30,39],约占以往研究的50%,随海拔升高而单调下降的曲线约占25%,另有25%单调递增或无明显趋势[40]。本研究得出坡地退化植物群落物种多样性与海拔之间呈单峰变化趋势(图4、5),这与Bhattarai等[41]的研究结果相近;与Tanaka和Sato研究结果不同,Tanaka和Sato认为在海拔梯度变化小的山区,物种多样性沿海拔变化呈双峰型变化趋势[42]。灌丛坡地Shannon-wiener指数和Simpson指数最大值出现于4215 m海拔处,草甸坡地出现于4280 m海拔处,两个坡地峰值出现海拔不一样,主要是因为灌丛坡地随着海拔升高,灌丛数量逐渐减少,坡顶甚至没有灌丛,草本层植物缺少了灌丛庇护以及受到强烈的太阳辐射,不适合部分植物生存,故物种多样性较低。
草甸坡地(图5)海拔4223 m左右Shannon-wiener指数和Simpson指数最小,主要是因为人类过度放牧,动物践踏,从而使植物盖度较低;海拔4260 m八条样线Shannon-wiener 指数异质性大,秃斑块分布密集,鼠洞和石头较多。海拔4320—4340 m处,八条样线Shannon-wiener指数差异较小,此海拔高度坡度较大,人类干扰相对较少,物种丰富度低,以嵩草属为主,其根对土壤具有缠绕支撑和串联等作用,故植被盖度高。Wu 等[43]研究表明,多样性指数主要受到地理环境、土壤条件以及干扰强度等影响。物种分布的上限通常被认为是由低温、降雪量、生长期和风力强度所决定的,而下限则可能是由气候和生物因素共同决定的[41]。
灌丛坡地多样性指数与海拔之间呈偏单峰曲线(图4、图5),海拔4215 m处,多样性指数达到峰值,随后沿海拔呈下降趋势,这从侧面反映出灌木及灌下草本层的生存和发展可能受到海拔制约;同一海拔不同样方间植被盖度空间异质性较大,主要是草地退化程度不一,鼠洞分布不均,土壤秃斑化,造成植被盖度降低,并随海拔升高,水热条件差,风力侵蚀严重,物种多样性较低。海拔4220—4240 m处以山生柳、高山绣线菊及金露梅为主,其隔离了畜群采食和践踏;改善局部光照、温度、水因子的分配,植物生长热量较充足,为草本植物发育提供了空生态位,减少了物种间竞争[20],其根系分布较深,对土壤具有缠绕吸附等作用,土层相对深厚、增强了土壤抗侵蚀能力;残体能够保护土壤,拦截径流,减弱剥蚀,灌木植物对草本植物分布产生明显影响[44];同时良好的气候条件为山地生态系统提供了最有效的初级生产力,并支持了更多物种的生存。
关于植物群落多样性与海拔之间呈现单峰格局的主要原因是人类过度干扰、水热环境以及物种竞争能力共同作用而形成的。低海拔地区,环境相对优越,像一些竞争力强的物种在群落中占据优势。但是,由于人类过度干扰,像放牧、播种等活动影响了一些物种,从而导致一些物种丧失,物种多样性较低,同时也由于受到坡度等影响,使人类活动在低海拔区域活动频繁,造成多样性低。高海拔地区,环境相对恶劣,比如气候寒冷,太阳辐射强烈等严酷环境,不适合部分物种生存,使其受环境影响较大。同时由于坡度等影响,坡地上坡位坡度较陡,再加上受到雨蚀、风蚀等作用,使水土流失更加严重,虽然人类干扰降低,但是水热条件及微气候不适合大部分物种生存,从而造成物种多样性较低。而在中海拔地区,人类干扰较低海拔地区少,水热条件优越,适宜更多物种生存,坡度较缓,土壤相对于高海拔地区肥沃,中间海拔地带则成为两个极端区域植物种类分异的过渡区域,因此物种多样性较高[6,45—46]。
3.3 坡地退化植物群落物种多样性与地上生物量的关系
地上生物量是反映群落植被生产力的重要指标[47],能够体现群落的结构特征和生长状况,山地的海拔梯度变化导致温度、湿度、热量和光照的变化,进而影响植物物种组成和群落结构[10]。草甸坡地地上生物量与海拔呈单峰曲线,灌丛坡地呈负相关(图6)。草甸坡地在海拔4280 m处,放牧活动相对较少,水热条件比较优越,土壤相对于高海拔地区较肥沃,中间海拔地带是两个极端区域植物种类分异的过渡区域,地上生物量较高,而灌丛坡地地上生物量呈下降趋势,海拔4260 m处最低,主要是随着海拔升高,土壤养分降低,温度降低,水热条件差,灌木不适宜生长,草本植物喜湿耐阴,灌木减少,草本层没有了灌木层的庇护,草本层植物较少,从而影响了地上生物量和多样性。
青藏高原相关研究中,物种多样性与地上生物量的关系有“单峰”曲线[30]、负相关[6,48]、正相关[16,49—50]和不相关[51]。本研究物种多样性与地上生物量的关系为正相关,物种丰富的植物群落往往表现出更高的生产力[52]。也有学者研究得出地上生物量与物种丰富度之间呈S型曲线[6],这与本研究结果不同。灌丛坡地地上生物量和物种丰富度大于草甸,坡地这与曲波等研究结果相似[45]。刘旻霞和马建祖[53]研究指出,北坡光照时间短,土壤含水量以及土壤养分等均大于南坡,因此北坡更适合更多物种生存,这与本研究结果相近,灌丛坡地(阴坡)地上生物量及物种多样性高于草甸坡地(阳坡)。
高寒草地是青藏高原主要的生态系统,其独特的地理位置和复杂的地理环境,对气候变化以及放牧干扰十分敏感[54]。许多高寒草甸生态系统由于过度放牧和气候变化而遭到破坏,逐渐退化形成大范围退化草地“黑土山”。在低海拔地区,缺乏足够降雨,不能满足植物生长所需的水分,植物生长热量比较充足,但土层相对较薄,持水能力较差[45]。坡度较低的地区植被覆盖度较低,人类活动对低海拔地区的生物多样性造成负面影响,该地区由于过度人类干扰,植被盖度较低,以及低海拔温和的条件导致种间竞争发生,造成其他物种消失,物种多样性降低,这可能主要是因为放牧活动在低海拔地区较频繁所导致的[55]。高海拔地区,寒冷的气候减缓植物生长和土壤发育,而且其他恶劣的环境,超过多数物种的生长耐受极限,限制了大多数物种的生长,从而导致植被盖度、物种多样性等均较低[10,45]。为了寻求更好的放牧条件,人类放牧活动从下坡位逐渐向植被盖度较高的上坡位迁移,从而造成上坡位发生退化,载畜量加重,草地生态日益恶化,最终导致草地生态系统失衡和崩溃。
本研究中物种多样性与地上生物量呈现正相关趋势,这种趋势可能是放牧强度差异、气候变化和植被退化共同作用的结果。物种多样性和生产力之间的关系也有部分可能是由于环境干扰所造成的。当环境同时促进多样性和生产力时,两者关系呈正相关,反之亦然;如果环境对一方是促进,另一方抑制,则呈现负相关甚至不相关趋势[5]。山坡温度、热量等因素随海拔高度的增加而降低。高海拔地区对气候变化和环境恶化异常敏感,温度升高的速率和幅度始终大于低海拔地区[56—57]。因此,深入研究坡地退化植被高寒草甸和高寒灌丛群落沿地理梯度的空间变化规律,可以丰富山坡植被生态研究,能够为未来的生态环境的恢复治理提供理论依据和科学支撑。
4 结论
三江源高寒山地生态系统中,气候变化、人类活动、啮齿动物以及地形等因素造成山地生态系统植被退化严重,坡地植被的下坡位受到人为因素和自然因素影响较大,退化比上坡位严重。坡地退化植物群落多样性和地上生物量随海拔升高先增大后减少,中间坡位植物多样性和地上生物量较高,植被受到干扰程度低,植被条件较好,退化程度轻。植物多样性和地上生物量之间呈线性增加趋势,物种丰富的植物群落往往能够表现出更高的地上生物量。地形、坡度、降雨和人类干扰等造成坡地植被发生严重退化,即使是在严重退化的环境中,植物群落多样性与地上生物量之间的关系仍符合青藏高原普遍分布规律。
三江源区坡地退化植被在治理恢复过程中,人类干扰活动要适度,以及在进行补播、围栏封育等措施的情况下,重视其管理和保护,制定合理的草地利用制度,以保证群落植被的完整性和生产力,以免植被群落再次受到破坏,导致生物多样性降低,生态系统稳定性降低。
猜你喜欢
水土保持学报(2022年5期)2022-10-10 14:20:06
西北农林科技大学学报(自然科学版)(2022年3期)2022-04-01 11:26:42
黑龙江工程学院学报(2020年5期)2020-10-21 05:37:10
水土保持研究(2019年6期)2019-10-19 03:33:32
草业科学(2019年5期)2019-06-04 09:36:24
幸福·婚姻版(2017年9期)2017-09-22 13:48:28
天津师范大学学报(自然科学版)(2016年4期)2016-12-14 06:04:26
防护林科技(2015年5期)2015-06-10 11:42:52
海南热带海洋学院学报(2015年5期)2015-03-14 09:51:07
河南科技(2014年12期)2014-02-27 14:10:46
