
“虚烦不得眠”证候的阐释:大鼠状态性焦虑及其睡眠时相变化
2022-05-24 10:40:24杨悦王艳艳卞宏生于爽李廷利黄莉莉
中医药学报 2022年5期
杨悦,王艳艳,卞宏生,于爽,李廷利,黄莉莉
(黑龙江中医药大学药学院,黑龙江 哈尔滨 150040)
黄莉莉(1978-),女,博士,教授,硕士生导师,主要从事中药生物活性及机制研究。
“虚烦不得眠”是酸枣仁汤主治证候。现代有关酸枣仁汤的药理研究或单独开展拮抗焦虑作用的研究,或单独开展改善睡眠研究,这种研究模式难以阐明酸枣仁汤的药理作用本质。在当前药理研究中焦虑动物模型和失眠动物模型都是单独复制,利用现有模型不能整体反映出“虚烦不得眠”的证候。基于这种现状,复制“虚烦不得眠”的动物模型是全面阐明酸枣仁汤作用实质的关键。因此我们利用条件性恐惧来复制动物的焦虑模型,并利用皮层脑电描记与分析技术分析其睡眠障碍的类型,以此来复制“虚烦不得眠”的动物模型,为今后酸枣仁汤的中药药理研究奠定基础。
1 材料与方法
1.1 实验动物
SD大鼠,雄性,体质量(220±20)g,SPF级,购自黑龙江中医药大学实验动物中心,许可证号:SCXK(黑)2018-007。饲养于标准独立通气笼IVC,自动定时光控系统实行12 h光照/12 h黑暗交替处理(7∶00 am开灯/7∶00 pm熄灯),恒温恒湿条件,湿度(50%±10%),温度(23±1)℃。4只/笼,可自由获取食水。
1.2 主要仪器和设备
RU25H5型独立通气笼IVC,苏州市苏杭科技器材有限公司;XR-XC404型SuperFcs条件性恐惧实验分析系统,上海欣软信息科技有限公司;XR-XG201型大鼠高架十字迷宫装置(EPM),上海欣软信息科技有限公司;MP-150型十六通道生理记录仪,美国Biopac公司;51600大鼠脑立体定位仪,美国Biopac公司。
1.3 方法
1.3.1 分组与动物模型的复制
1.3.1.1 大鼠皮层脑电与肌电电极的埋置
4%水合氯醛将大鼠麻醉后,固定于脑立体定位仪,碘酒消毒头部创口并剔除毛发,剪去头部皮肤,酸蚀剂去除组织筋膜,盐水清洁使充分暴露颅骨,待颅骨干洁时于前囟前1 mm,额骨中线旁开1.5 mm;人字缝前1 mm,顶骨中线旁开1.5 mm埋置2根皮层脑电电极,颈部肌肉埋置肌电电极,牙托粉固定后恢复1周,期间自由摄入食水。
1.3.1.2 条件性恐惧所致大鼠状态性焦虑模型的复制
SD雄性大鼠随机分为对照组和模型组,每组8只,IVC系统适应性饲养7 d,条件性恐惧刺激的前3天开始,每天同一操作人员抚摸大鼠 2~3 min,持续3 d,使大鼠习惯实验人员。实验于12∶00—18∶00进行。第8天开始实验。在条件性恐惧刺激前30 min,将实验动物分单笼放置于条件性刺激装置所在环境中适应。每次抓取一只大鼠轻抚鼠背后提取鼠尾托住大鼠,使头部正对箱门放入。刺激箱中适应3 min后开始条件性刺激。模型复制方法采取5 s噪声(1 000 Hz,75 dB),第5 s持续1 s足底电击共同终止的刺激方案(0.5 mA),该事件循环发生15次,每次间隔1 min;对照组则只施加5 s噪声而无电击(1 000 Hz,75 dB),其他实验要求与模型组相同,循环15次,每次间隔1 min。每只动物实验结束后于刺激箱中适应3 min再放回原单笼,送回饲养室,并用10%的酒精擦除箱中的动物排泄物及残留气味。连续进行4 d模型复制。
僵直反应(Freezing)指标的判断:模型复制后翌日,将大鼠置于条件性刺激箱,适应3 min后,记录5 min内每次声信号后大鼠头、躯体、四肢均不动的行为时间。
1.3.2 高架十字迷宫实验方法与观测指标的判断
测试于12∶00开始。在行为学测试前30 min,待测动物分单笼放置于EPM所在安静环境中适应。实验开始时,将实验动物置于EPM中央平台区,面向一侧开放臂释放同时开始计时,每只动物自由探索5 min,通过录像采集行为指标,每次测试结束后将动物放回原笼送回饲养室,并用10%的酒精彻底清洁,去除动物排泄物和气味。指标的生物学意义与判断方法如下[1-3]。
开放臂运动时间(Time in open arms):直观反映动物对高悬敞开环境的厌恶程度(四肢完全进去任意开放臂的停留时间,直至四肢完全脱离开放臂);
封闭臂运动时间(Time in closed arms):直观反映动物对相对封闭环境的偏好(四肢完全进去任意封闭臂的停留时间,直至四肢完全脱离封闭臂);
开放臂进入次数(Open arm entries):体现对新奇环境的探究性(四只爪完全进去任意开放臂的频次,只两前肢进入记为探究开臂);
总入臂次数(Total arm entries):总入臂次数可体现动物的运动活性,在无其他外因干扰下,次数减少反映为焦虑水平增加(四只爪完全进去任意开放臂和封闭臂频次之和);


1.3.3 条件性恐惧所致大鼠状态性焦虑持续时间及其睡眠-觉醒周期变化的研究
在上述实验条件下,实验动物经历连续4 d的条件性刺激。模型复制后的翌日开始首次条件性刺激后的EPM行为学测试(第1天)。所有实验动物分别于条件性刺激后第1、7、10、14、21天进行EPM中的行为学评价,录像记录相关行为参数。每日测试完成后连接十六通道生理记录仪,连续描记24 h的脑电图。
1.4 统计学处理方法
本研究应用IBM SPSS Statistics 24软件进行统计分析,两组间比较采用独立样本t检验;连续检测数据采用重复测量方差分析,非正态性数据采用Mann-WhitneyU非参数检验,计量资料以均数±标准差或中位数(四分位距)统计描述。P<0.05认为差异有统计学意义。
2 结果
2.1 条件性恐惧所致状态性焦虑大鼠僵直反应时间的变化
结果显示,与对照组相比,造模后大鼠僵直反应时间显著增加(P<0.01),见表1。

表1 条件性恐惧所致状态性焦虑大鼠僵直反应时间的变化
2.2 条件性恐惧所致状态性焦虑大鼠EPM中的行为学变化
2.2.1 开放臂运动时间、封闭臂运动时间、开放臂进入次数及总入臂次数的变化
与对照组相比,不同测试天数下的模型组大鼠开放臂运动时间于第1、7和10天显著缩短(P<0.05)、开放臂进入次数于第1、7、10、14天显著降低(P<0.05),总入臂次数于第1、7、10和21天显著减少,封闭臂运动时间略有延长而未见明显变化,见图1。

注:与同期对照组比较,*P<0.05。图1 条件性恐惧大鼠EPM中开放臂运动时间、封闭臂运动时间、开放臂进入次数及总入臂次数的变化
2.2.2 进入开放臂时间与次数比例的变化
与对照组相比,不同测试天数下的模型组大鼠开放臂进入次数比例和运动时间比例均降低,其中第1 、7、10、14天呈显著降低,差异有统计学意义(P<0.05),见图2。
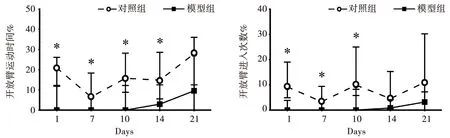
注:与同期对照组比较,*P<0.05。图2 条件性恐惧大鼠EPM中开臂时间比例及次数比例的变化
2.3 条件性恐惧所致状态性焦虑大鼠觉醒-睡眠时间和睡眠时相的变化
2.3.1 条件性恐惧所致状态性焦虑大鼠觉醒-睡眠时间的变化
2.3.1.1 条件性恐惧所致状态性焦虑大鼠第1、7、10、14、21天Wake时间的变化
与对照组相比,不同测试天数下状态性焦虑大鼠的Wake时间明显延长(P<0.01,P<0.05),其中第1、7、10、14天大鼠Wake时间的变化更加明显,结果见表2~表6。

表2 条件性恐惧所致状态性焦虑大鼠第1天Wake时间变化
表3 条件性恐惧所致状态性焦虑大鼠第7天Wake时间变化
表4 条件性恐惧所致状态性焦虑大鼠第10天Wake时间变化
表5 条件性恐惧所致状态性焦虑大鼠第14天Wake时间的变化
表6 条件性恐惧所致状态性焦虑大鼠第21天Wake时间变化
2.3.1.2 条件性恐惧所致状态性焦虑大鼠第1、7、10、14、21天NREMS时间的变化
与对照组相比,不同测试天数下状态性焦虑大鼠的NREMS时间明显缩短(P<0.01,P<0.05),其中第1、7、10、14天大鼠NREMS时间的变化更加明显,见表7~表11。
表7 条件性恐惧所致状态性焦虑大鼠第1天NREM睡眠时间变化
表8 条件性恐惧所致状态性焦虑大鼠第7天NREM睡眠时间变化
表9 条件性恐惧所致状态性焦虑大鼠第10天NREM睡眠时间变化
表10 条件性恐惧所致状态性焦虑大鼠第14天NREM睡眠时间变化
表11 条件性恐惧所致状态性焦虑大鼠第21天NREM睡眠时间变化
2.3.1.3 条件性恐惧所致状态性焦虑大鼠第1、7、10、14、21天REMS时间变化
与对照组相比,不同测试天数下状态性焦虑大鼠REM睡眠时间明显增加(P<0.01,P<0.05),其中第1、7、10、14天大鼠REMS时间的变化更加明显,见表12~表16。
表12 条件性恐惧所致状态性焦虑大鼠第1天REM睡眠时间变化
表13 条件性恐惧所致状态性焦虑大鼠第7天REM睡眠时间变化
表14 条件性恐惧所致状态性焦虑大鼠第10天REM睡眠时间变化
表15 条件性恐惧所致状态性焦虑大鼠第14天REM睡眠时间变化
表16 条件性恐惧所致状态性焦虑大鼠第21天REM睡眠时间变化
2.3.2 条件性恐惧所致状态性焦虑大鼠觉醒-睡眠时相比例的变化
与对照组相比,不同测试天数下状态性焦虑大鼠的Wake时间比例于第7、10、14天明显增加(P<0.01,P<0.05),伴有NREMS时间比例第1、7、10、14天的明显减少(P<0.01,P<0.05)和REMS时间比例第1天的明显增加(P<0.05),见表17。

表17 条件性恐惧所致状态性焦虑大鼠觉醒-睡眠时相比例的变化
3 讨论
酸枣仁汤出自汉·张仲景《金匮要略》,主治“虚劳,虚烦不得眠”。通过原著的温习及参考相关文献,我们认为“虚劳,虚烦不得眠”的内涵是“虚劳”,是“虚烦不得眠”病因病机之所在,而“虚烦不得眠”是“虚劳”的证候之一,这一推断可从《金匮要略·血痹虚劳病脉证并治六》分析得出。“血痹虚劳病脉证并治六”篇中涉及脉证九条及方九首,通过对“脉证九条及方九首”进行分析,“方九首”所治之病均为“血痹或虚劳病”,但不同的方主治证候各有不同,酸枣仁汤主治证候是“虚烦不得眠”,而且酸枣仁汤主治的“虚烦不得眠”之证候是一个整体不可分割。但是纵观现代开展的有关酸枣仁汤的中药药理研究,不是基于其“虚烦”之症单独开展拮抗焦虑作用的研究,就是基于其“不得眠”之症单独开展改善睡眠作用的研究,这种研究范式,难以阐明酸枣仁汤的药理作用本质。我们认为要想阐明酸枣仁汤的药理作用本质就必须围绕酸枣仁汤主治“虚烦不得眠”这一特殊证候来开展。基于此,复制“虚烦不得眠”这一动物模型就成为了阐明酸枣仁汤的药理作用本质的关键之所在。
在药理研究中焦虑与失眠动物模型是分别复制的,故不能反映“虚烦不得眠”这一证候,而且无论是焦虑还是失眠动物模型的复制都各自存在许多不同的复制方法。我们基于酸枣仁汤所治“虚烦不得眠”之证候由肝血不足,阴虚内热而起。肝藏血,血舍魂,若肝血不足,魂不守舍,心失所养,则虚烦不眠,心悸不安;血亏阴虚,易生内热,虚热内扰,虚烦不安。《医学正传》指出:“恐通肾,惊通心肝”。惊恐太过则可导致肝血不足,魂不守舍,心失所养,则虚烦不眠的认知,利用条件性恐惧复制大鼠状态性焦虑模型,并利用皮层脑电描记与分析技术,来复制“虚烦不得眠”证候的模型,为今后阐明酸枣仁汤的药理作用本质奠定基础。
在当前中医情志病证的实验研究中多用行为学方法来判定[4-5],其中焦虑动物模型大多借用国际上经典的动物模型,包括高架十字迷宫、天敌暴露、孤养等非条件化模型,以及Geller-Seifter和Vogel饮水冲突实验、防御性掩埋等条件化模型,条件性恐惧所致焦虑模型可作为焦虑动物模型的一种。DAVIS[6]发现经历条件性恐惧训练动物的行为表现,与临床焦虑症患者的表征是相似的。其理论基础是条件性刺激引发动物对已知但又不可预期事件的恐惧和焦虑,可模仿人类生理反应的行为表型(僵直或回避),且苯二氮类、巴比妥类、阿片类、SSRI等均已表现出对条件性恐惧模型的抗焦虑活性。
动物模型的效果为暂时性的生理改变,随时间推移造模效果趋于消散,明确焦虑动物模型的持续时间,以便更合理应用于实验研究,对焦虑发病机制和有关药物评价研究具重要意义。就目前21 d行为学测试周期的研究结果可知,条件性恐惧可增加大鼠对厌恶环境的恐惧程度和对安全区域的偏好,并伴有运动性水平的降低,整体行为呈现明显的探究性抑制,模型效果随时间延长逐渐消退[7-8],前14天更显著的焦虑行为指标提示14 d内更明显的焦虑水平。以往条件性恐惧模型多在训练后短期内争取模型效果消退前的药效评价[9-10]。虽然中药在治疗焦虑上有很好的疗效,且与化学合成药相比,中医药治疗具有个体化用药、治疗多靶点多途径的优势[11-13],但存在起效慢、给药周期长的特点,明确条件性恐惧诱导大鼠状态性焦虑模型的持续时间,为今后中药药效治疗而非预防作用的评价提供方法学借鉴。
睡眠障碍常作为广泛性焦虑症的一项症状学诊断标准,而睡眠又是多项指标代表的复杂过程,脑电图是睡眠测量的客观手段,焦虑症患者的睡眠问题体现在入睡潜伏期延长,总睡眠时间和睡眠深度降低,睡眠连续性不佳等[14]。中医在探讨情志学说时常出现“不寐”“不得眠”的证候,如《类证治裁·不寐》曰:“思虑伤脾,脾血亏损,经年不寐”;《景岳全书·不寐》谈到:“无邪而不寐者,必营气不足也,营主血,血虚则无以养心,心虚则神不守舍”;《金匮要略·血痹虚劳病脉证并治第六》篇指出:“虚劳虚烦不得眠,酸枣汤主之”;《伤寒论·辨太阳病脉证并治》第76条提到:“发汗吐下后,虚烦不得眠,若剧者,必反复颠倒,心中懊恼,栀子豉汤主之”等。表明了情志不遂,劳动思虑太过诱发失眠的病因,说明焦虑症与虚劳不寐之间一定的相关性[15-16]。本研究在状态性焦虑模型复制基础上进行睡眠监测,发现状态性焦虑大鼠伴有以高觉醒和NREMS减少为主要特征的睡眠-觉醒周期变化,REMS于模型后第1天有明显增加,后期略有减少。相关报道指出,NREMS在抵御焦虑症的发展中扮重要角色,NREMS纺锤波和κ复合波阶段是记忆巩固的关键过程,慢波睡眠阶段具焦虑缓和作用,而高度焦虑个体表现为NREMS百分比降低[17]。REMS在情境记忆巩固中起关键作用,REMS压力包括更短的REM潜伏期,更多的REM密度和REM睡眠持续时间,但有关REMS的探讨缺乏一致性,不同的睡眠深度和REM改变可能反映不同的症状[18]。明确条件性恐惧所致状态性焦虑对睡眠-觉醒周期的影响,为阐明“虚烦不得眠”证候的科学实质提供实验依据,为今后开展中医焦虑性失眠的相关研究增设了平台基础。
综上所述,本研究通过条件性恐惧方法诱导大鼠的状态性焦虑模型,并观察其睡眠时相的变化,“虚烦不得眠”证候主要表现为状态性焦虑并发的睡眠时相障碍性改变。在探讨中医焦虑性失眠研究中,可作为中药治疗而非预防作用的评价手段。
猜你喜欢
保健与生活(2023年17期)2023-10-16 23:22:40
保健与生活(2023年19期)2023-08-15 00:48:36
基层中医药(2021年12期)2021-06-05 06:56:48
今传媒(2019年10期)2019-11-11 12:32:26
家庭医学(2017年9期)2017-10-24 15:44:23
中国卫生标准管理(2015年25期)2016-01-14 09:29:28
哈尔滨医药(2015年2期)2015-12-01 03:57:34
当代党员(2014年11期)2014-11-05 21:50:58
心理科学进展(2014年3期)2014-04-29 00:44:03
食品科学(2013年22期)2013-03-11 18:29:26
