
光照和基质对布氏轮藻生长、色素组成及有性繁殖的影响
2022-05-23 02:39:02王和云陈俊格易春龙
湖北农业科学 2022年8期
王和云,孙 彤,周 易,金 辉,陈俊格,易春龙
(1.湖北工业大学土木建筑与环境学院/河湖生态修复及藻类利用湖北省重点实验室,武汉 430068;2.中工武大设计集团有限公司,武汉 430223)
在湖泊、池塘和水库等水域中,轮藻植物是沉水植被的重要组分之一。轮藻科约有400个种,遍布于各类水体中,包括淡水和咸水、临时性和永久性的,从小池塘到大湖泊[1]。轮藻多占据被子植物不能利用的小生境,作为指示类群,能反映水体健康状况[2],是受损生态系统功能恢复的先锋种[3,4]。
光照是轮藻生长的关键因子[5,6]。不同轮藻种类,光适应机制不同,反映它们在湖泊中分布深度不同[7-9]。改变色素组成或含量,调整光合效率是自养生物在光合生理上最常采用的光适应机制[10,11]。近年来,已经有研究报道轮藻在弱光照[12]和强光照[8,9]条件下的光合色素含量以及组成。此外,沉水植物对光照的适应在形态上包括低光下茎的伸长和形成更大而薄的叶片以使单位生物量上的光吸收量增大[13-15]。轮藻在强光照下产生的小枝密集、茎粗壮矮小,而在弱光照下小枝分散、茎纤细[14,16]。
沉水植物生长对光照和基质的响应和它们的生长型密切相关,冠层生长型植物如穗花狐尾藻受基质的影响大,而底栖型植物如轮藻[17,18]和莲座型植物如苦草受光的影响大[15]。布氏轮藻(Ch ar a b rauniiGmel.)曾广布于世界各大洲[19],但随着水体环境的恶化,布氏轮藻在欧洲已成濒危种[20,21]。水稻田和池塘曾是布氏轮藻的主要分布水体[22],但随着经济发展和农业环境改变,轮藻种群数量锐减,部分敏感种类已在原分布区域消失[23]。本研究以布氏轮藻为研究对象,选择不同的基质类型,模拟3种光环境,研究光照和基质对轮藻形态、色素模式和有性繁殖的影响,以期为轮藻植物的修复提供基础数据。
1 材料与方法
1.1 试验材料
布氏轮藻(Ch ar a b r auniiGmel.),雌雄同株,被少量钙质,鲜绿色或者暗绿色。节间与小枝多等长,托叶单轮,与小枝数相等,二者互生,雌雄配子囊混生于小枝下部的1-3个节上。试验材料采自湖北省武汉市梁子湖。
1.2 试验设计
2018年4月15日至5月22日,试验在湖北工业大学资源与环境工程学院室外温室内进行,植株生长适应时间(不加盖遮阴网)为1周,试验时间为4周,共持续时间为5周。试验是光照和基质的2因子9处理(3×3)交叉试验,基质处理包括腐质底泥(来自梁子湖),黏土(400目网筛过滤)和湖水冲洗后的细沙;光处理包括50%(N)、25%(M)和5%(D)自然光照,3种不同光照水平通过加盖不同密度的同质遮阴网来获得。54个白色塑料桶(Φ=30 cm,h=35 cm)分别置于3种不同遮阴条件下,每种光照处理18个,桶中分别装有底泥(S)、黏土(C)和细沙(s),每个桶种1株布氏轮藻,植株间差异极小。预试验处理的1周中,细沙中生长的植株迅速产生大量白色的雌雄配子,而植株本身没有生长,有衰败迹象。因此,株高、色素和生物量的数据不包括细沙基质上各种光照处理,为3光照×2基质的2因子6处理(NS、MS、DS、NC、MC和DC)的交叉试验。塑料桶中水温通过循环换水保持在(25±1)℃。
1.3 指标测定
植株色素测量参照Küster等[8]方法,取轮藻顶端鲜重60 mg,清洗干净附着物,吸干水分,称重,用N,N-二甲基甲酰胺(DMF)定容至6 mL(当吸收值大于1时重新取样定容至8 mL),避光放置过夜,用752分光光度计测定664 nm和647 nm处吸光度。叶绿素a和叶绿素b的计算公式如下所示:
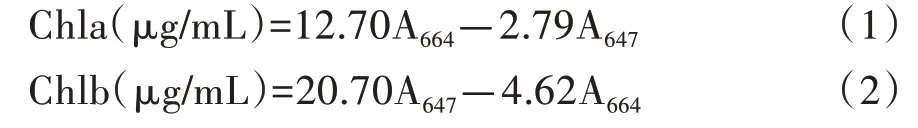
总类胡萝卜素含量需要利用萃取液470 nm处光吸收度,计算公式如下所示:
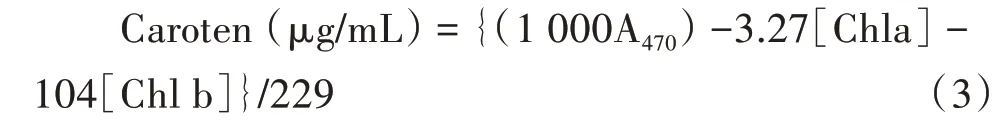
基质营养TN采用半微量凯氏定氮法,TP测量采样钼锑比色法。每隔3~4 d利用自动水质监测仪(YSI公司,American)测量水质(包括pH、电导率、浊度、溶氧量和温度)。
每隔1周记录植株上雌雄配子和受精卵的出现时间及其颜色,5周后收获,测量植株高度(假根到冠层的平均距离),测量植株干重(24 h、95℃)和灰分比率(3 h、450℃)。
1.4 数据分析
一元方差(ANOVA-one way)分别分析光照和基质对植物生长的影响,二元方差(ANOVA-two way)分析2×3因子的交叉影响。多重比较运用Tukey’s post-test。
2 结果与分析
2.1 水质理化指标
塑料桶中水质指标如表1。黏土基质中水体pH比底泥基质稍高,其他指标差异不一致。
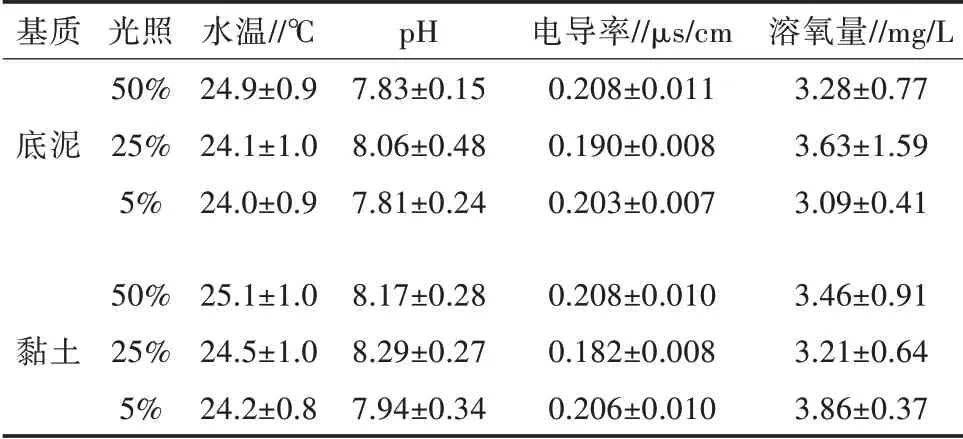
表1 水温、酸碱度、电导和溶氧量
2.2 植株高度
二元方差分析(ANOVA-two way)表明,光照和基质对植株的高度有显著影响,交叉作用也极显著(表2)。随着光照度的减少,两种基质上生长的植株高度均明显增加,黏土基质上的株高小于底泥基质上的株高(图1)。
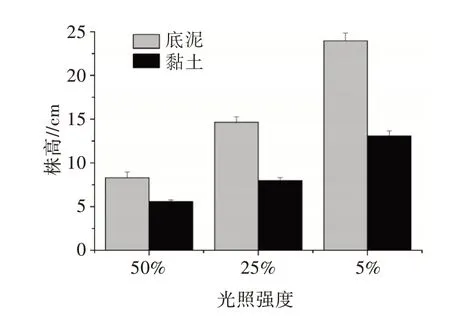
图1 不同光照和基质处理下布氏轮藻的株高
2.3 灰分比率和干重
灰分比率和光照强度正相关,黏土基质下的灰分比率高于底泥基质上的灰分比率(图2A)。光照和基质对植株的干重具有极显著影响和显著交叉影响(表2),两种基质条件下最大干重均出现在25%光处理条件下,最小干重在最低光条件下(图2B)。
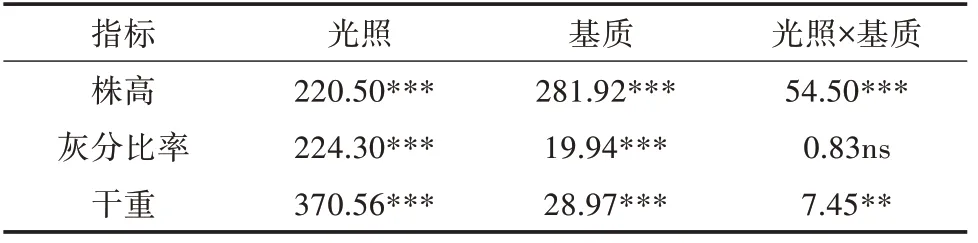
表2 光照和基质对布氏轮藻高度、灰分比率和干重的二元方差分析
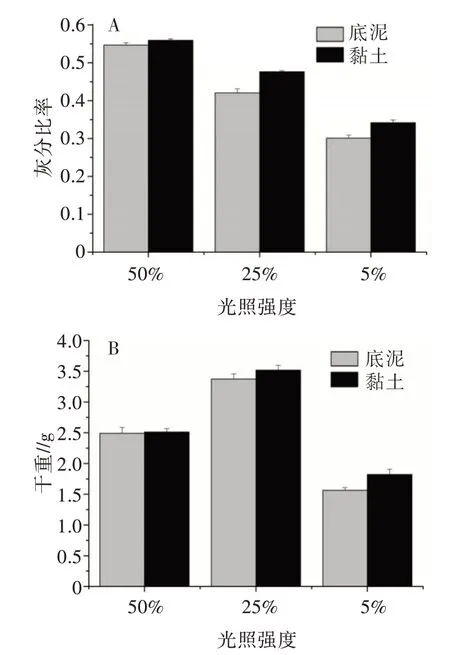
图2 不同光照和基质处理下布氏轮藻的灰分比率(A)和干重(B)
2.4 色素
随着光照的减少,两种基质上生长的植株顶端单位鲜重基础上的叶绿素a(Chla)、叶绿素b(Chlb)和类胡罗卜素(Caroten)含量均显著增加,黏土基质上的植株色素含量低于底泥基质上的植株色素含量(表3)。二元方差分析(ANOVA-two way)结果显示,光照和基质对Chla、Chlb、Chl(a+b)和Caro含量均有极显著影响,其交互作用也极显著(表3)。就处理因子对色素比率的影响而言,Chla/Caroten受到光照、基质及其交叉作用显著影响,基质对Chla/Chlb没有显著影响(表4)。随着光照的减少,两种基质条件下的Chla/Caroten表现出极显著的增加(图3B),Chla/Chlb表现出不同的趋势,在底泥基质处理条件下,在光照降至5%时,Chla/Chlb显著上升,而在黏土基质处理条件下,随着光照的减少,Chla/Chlb表现出递减的趋势(图3A)。
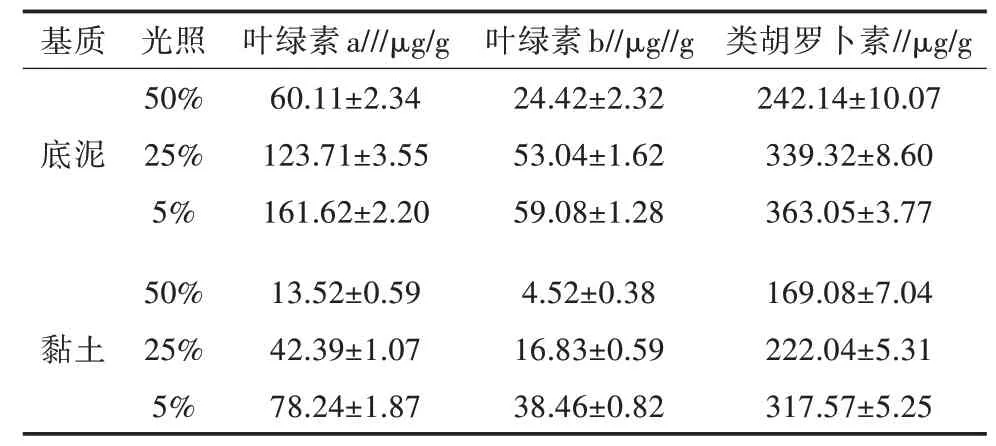
表3 布氏轮藻在3种光照和两种基质处理下单位鲜重上的叶绿素a、叶绿素b和类胡萝卜素含量
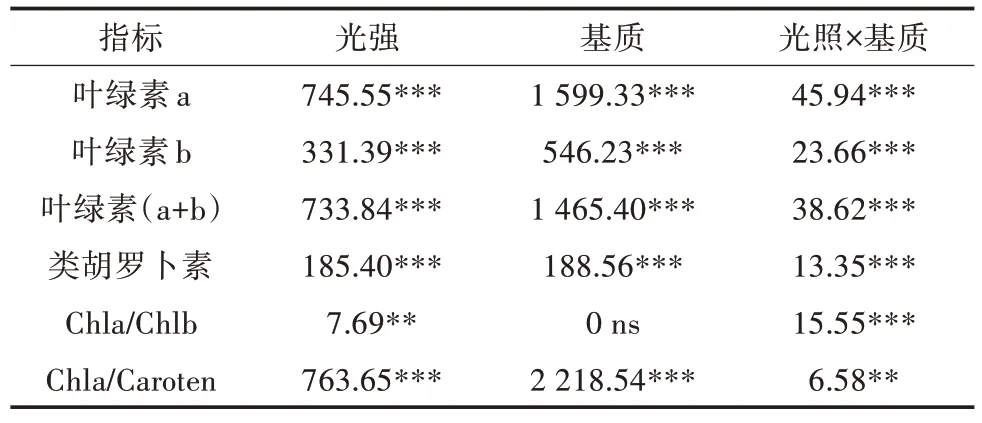
表4 不同光照和基质对布氏轮藻色素含量和比例的二元方差分析
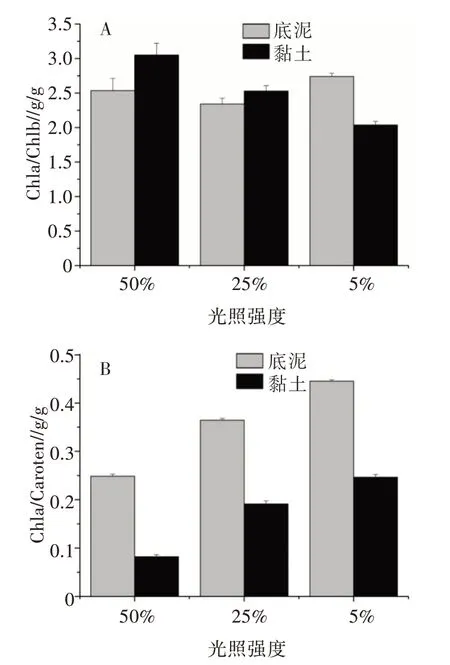
图3 不同光照和基质处理下布氏轮藻的Chla/Chlb(A)和Chla/Caroten(B)
2.5 有性繁殖
试验进行2周后(5月7日),所有处理中都出现了雌雄配子,黏土基质上的光照(NC)中已有受精卵出现。5月16日,所有处理中都出现了受精卵,其中底泥基质的5%光照下(DS)也出现了零星的黑色受精卵。
3 讨论
3.1 形态特征和色素
轮藻在强光照下产生茎粗壮小枝密集而在弱光下茎纤细小枝分散,前者是强光照下的自我保护[16],后者则是对低光照的适应[12]。由于Chlb属于捕光色素,Chla/Chlb的降低通常被认为是轮藻对低光条件的一种适应[12,14]。该研究中黏土基壤上光照的降低导致Chla/Chlb的降低,轮藻表现出对低光条件的适应;但其在底泥基壤上的Chla/Chlb变化不一致。随着光照的降低,布氏轮藻植株顶端色素的Chla比Chlb增加的比率大得多。不是所有的轮藻种类都通过调整Chla/Chlb来适应低光环境,如C.canes cens[8]和C.inter med iaBraun[16]在变动的光强范围内Chla/Chlb没有显著性变化。
Chla/Caroten的比值通常与植物的光保护机制联系在一起[11]。该研究中,轮藻植株顶端色素Chla/Caroten的比值随着光照的减少而增加(图4B),表明类胡萝卜素的比率与光照正相关,已在一些其他关于轮藻研究中发现同样的规律[8,12,16],植株在高光照条件下,光保护能力增加和光保护色素增加的象征[24,25]。同一光照下黏土基质上较低的Chla/Caroten,也是自我保护增强的象征,这可能表明当植物处于强光照胁迫下时,底泥上生长的轮藻植株具有更高的耐强光抑制能力。
3.2 灰分比率和干重
对于植物体表面能富集钙质的轮藻种来说,其灰分比率通常是其在特殊生境中光合能力的一个指标[26]。该研究中,在两种基质条件下,布氏轮藻的灰分比率均随着光照的下降而降低,表明植株光合作用随光照下降而降低。当水体pH升高时,布氏轮藻通常在植株表面产生大量结晶[27]。该研究中,黏土基质上生长的轮藻灰分比率高于底泥基壤上的轮藻,这可能与黏土基质水体pH更高有关。轮藻的灰分成分中除了钙盐外,还含有轮藻茎上附着的悬浮粒子,主要是一些泥沙,是植物在强光照下的一种自我保护[28]。黏土基壤上的Chla/Caroten显著小于底泥上的Chla/Caroten,也证明了相同光照条件下,黏土基质要求植物投入更多的保护色素。
3.3 有性繁殖
光照对轮藻有性繁殖具有促进作用,已被研究证明[14,29,30]。布氏轮藻可通过快速有性繁殖来适应即将枯水的逆境[27]和延迟有性繁殖来适应水深的增加[13]。本研究中不同光照下的布氏轮藻出现雌雄配子和受精卵的时间没有差异,说明光照的差异并没有影响有性繁殖的出现时间。这可能是因为该研究中各种光照处理下各个塑料桶中的水温没有显著性差异,因为温度相对于光照,对轮藻有性繁殖的发生时间的影响更大[31]。而黏土基质上的受精卵比底泥上先出现是植物对光照胁迫的反应。黏土基质上更高的灰分含量和更高的保护色素都表明黏土基质中自然光照对轮藻的生长有抑制作用。由于细沙、土壤和水比热容不同,在相同水温下,基质中轮藻根系处的温度可能存在显著差异,这种差异最终影响植物的生长和繁殖。因此,黏土基质中强光照对轮藻生长的抑制可能是通过基质内高温对轮藻生长的胁迫来实现,这可能也是沙基质上迅速产生大量受精卵的主要原因。
4 小结
布氏轮藻通过避离水表层、缩短株高和生成大量的类胡萝卜素(Caroten)来适应强光照环境,相反通过伸长至水表层的茎和大量的叶绿素b(Chlb)来适应弱光照。与底泥基质相比,黏土基质株高更低,植株灰分比率更高,类胡萝卜素比率增大,表明温室高光热条件下,黏土基质需要投入更多的能量用于自我保护机制上,黏土基质中植株上较早出现的受精卵可能也是对强光照胁迫的一种响应。
猜你喜欢
选煤技术(2022年2期)2022-06-06 09:13:10
选煤技术(2022年2期)2022-06-06 09:12:22
选煤技术(2021年6期)2021-04-19 12:22:08
四川蚕业(2021年4期)2021-03-08 02:59:56
皮革制作与环保科技(2020年14期)2020-03-17 07:16:22
中国医药指南(2017年30期)2017-01-15 15:23:07
水科学与工程技术(2016年2期)2016-07-10 10:19:57
浙江大学学报(工学版)(2016年9期)2016-06-05 09:20:57
浙江农业科学(2016年11期)2016-05-04 04:16:54
铜业工程(2015年4期)2015-12-29 02:48:34
