
人工修复措施对退化高寒草甸土壤养分及酶活性的影响
2022-05-19 12:49段成伟李希来马盼盼徐文印苏乐乐杨鑫光
西北农业学报 2022年4期
段成伟,李希来,马盼盼,徐文印,柴 瑜,苏乐乐,杨鑫光
(1.青海大学 农牧学院,西宁 810016;2.青海民族大学 生态环境与资源学院,西宁 810007)
草地作为陆地生态系统的重要组成部分,对维持陆地生态系统的稳定与平衡,人类社会可持续发展具有重要意义[1]。作为青藏高原独特的生态系统,高寒草甸生态系统对草地畜牧业持续发展和牧民生活水平的提高具有重要作用[2-3],其牧草生物量不仅决定草地载畜量和草地发展潜力,也反映草地的退化程度。高寒草甸生态系统具有防风固沙、调节气候、涵养水源等多种生态服务功能,是中国的重要生态安全屏障[4]。近年来,由于气候变化和人为破坏等一系列因素,高寒草甸植被生产力下降、草地地上生物量减少、物种多样性降低[5-6]、土壤养分流失[7-8]、草地植物生态系统遭到严重干扰和破坏,高寒草甸已呈现出不同程度的退化趋势[9-10]。因此,退化高寒草地急需快速有效治理。
草地退化的根本原因是土壤退化,土壤环境是动植物和微生物共同生活的载体,其理化性质在空间和时间异质性上共同决定着草地植被分布格局[11]。作为草地生态系统物质转换和能量流动的重要枢纽,草地土壤通过物理、化学、生物等过程耦合形成土壤养分,为植物的生长提供营养元素,并有效地调节植物的功能属性[12],土壤是植物生长的基质,土壤养分直接与土壤肥力相关[13]。土壤有机物经微生物分解转化成C、N、P等供植物可利用形态,而土壤C、N、P可反映出关键养分的限制。土壤 pH虽然不能直接表明土壤中的养分含量,但其大小可对土壤中微生物区系进行控制,从而影响土壤中营养元素的形态、转化方向和转化过程。土壤酶作为土壤物质循环和能量流动的重要组成部分,其活性可反映土壤中多种生物化学过程的相对强度,相比土壤特性其活性大小既反映土壤营养物质的储量,也体现土壤中生物化学反应的方向和强度[14]。
国内生态学者实施重度退化翻耕建植人工草地,中度退化免耕补播施肥,轻度退化围栏封育、休牧、啮齿动物防控等措施致力于三江源区不同退化程度草地的恢复[15-19]。通过对退化草地施肥补给养分,大量研究验证对植被恢复和毒草防控的影响[20-22]。退化草地有机肥添加是改善并维持草地生态系统营养物质平衡的治理措施,有机肥输入不仅会促进植物生长、提高草地生产力[23],有助于退化草地的恢复。免耕补播是成本低廉、投资少、技术易控、见效快的草地恢复措施,能快速增加草地丰富度、草地覆盖度以及草层的产量及品质[24]。目前草地恢复相关研究主要集中在各种人工修复措施对植被动态以及产量的影响,而对土壤养分和土壤酶方面的研究较少[25],并且以往的研究治理大多局限于小范围设置样方,对大面积样地退化草地的恢复治理研究较少,其研究没有代表性和整体性,达不到快速恢复的效果。本研究以青藏高原退化高寒草甸为研究对象,结合实地调查结果,选择发生重度退化的高寒草甸,在灭鼠的基础上采用免耕补播、施肥和围栏封育等技术措施开展退化草地土壤修复治理工作,探讨不同人工修复措施的适宜性。通过研究人工修复措施对土壤养分及养分相关酶活性特征的影响及其相互关系,为退化草地土壤改良和草地恢复等快速修复技术制定提供理论基础和技术支撑,为河南县生态大牧场草地的合理利用与科学管理提供科学依据。
1 材料与方法
1.1 研究区域概况
研究区位于青海省东南部河南蒙古族自治县多松乡黄河二级流域永曲河流域附近(N:34°18′43.30″~34°19′59.54″,E:101°22′24.98″~101°28′19.26″),海拔3 418~3 567 m,样地坡向南偏东15~18 °,坡度3~5 °为缓坡,地势呈东高西低。草地类型为高寒草甸,土壤类型为山地草甸土。整个区域属高原大陆性气候,全年四季变化不明显,常年多为大风天气。年均温为0.5 ℃,最冷月均温为-10.0 ℃,最热月均温为9.4 ℃。年降水量597.0~615.5 mm,年均蒸发量1 350 mm。年日照时数2 551~2 577 h,日照率60%。研究区高寒草甸已发生重度退化,高原鼠兔活动频繁,植被盖度降低,裸露秃斑面积增加,属于黑土滩退化草地。研究区域内物种组成以禾草和杂草为主,优势种主要为鹅绒委陵菜(Potentillaanserina)、垂穗披碱草(Elymusnutans)、黄帚橐吾(Ligulariavirgaurea)和矮嵩草(Kobresiahumilis)等。
1.2 样地设置
结合实地调查结果,在研究区选择10 hm2发生重度退化的高寒草甸,且退化程度相对一致、质地均一、地势较为平坦的阳坡缓坡坡底。在灭鼠的基础上采用免耕补播(牧草免耕播种机GB/T 25421)、施肥(人工撒施)和围栏封育的技术措施开展退化草地修复治理工作(表1和图1)。为防止放牧家畜以及其他大型草食动物的干扰,试验区自2020年起进行围栏封育,围栏采用刺丝封育。2020年6月初在试验区内随机选择4个 33 331.8 m2(182.57×182.57 m)样地,分别对草地进行施肥(F)、免耕补播(S)及施肥+免耕补播(F+S)等修复措施。同时选取面积相同且不经过任何处理的草地作为对照(CK)。

图1 样地设置

表1 试验地基本概况
施肥种类为有机肥(有机质≥45%,水份≤30%,总养分(N+P2O5+K2O)≥5.0%,执行标准:NY525-2012)900 kg·hm-2;免耕补播处理为草地牧草混播,草种为垂穗披碱草(Elymusnutans)、中华羊茅(Festucasinensis)及冷地早熟禾(Poacrymophila)3种当地优势草种,垂穗披碱草15 kg·hm-2,中华羊茅7.5 kg·hm-2,冷地早熟禾7.5 kg·hm-2,混播比例为2∶1∶1;施肥+免耕补播处理为双因素控制。种子均由当地草籽繁殖场生产,有机肥购于睿泽科技有限 公司。
1.3 样品采集
于2020年8月中旬植物生长旺盛期取样。在每个样地内间隔10 m设置4条长50 m的样线,分别在每条样线的0 m、10 m、20 m、30 m、 40 m处设置50 cm×50 cm的样方,进行植被调查并采集植物和土壤样品。植被调查时,记录各物种和总盖度,并齐地面分种剪取植物地上部分,测定植物地上生物量,然后在每个样方内用直径 10 cm 的根钻取0~20 cm根样,用直径5 cm的土钻采集0~20 cm土壤样品,并将每个样方内的土样混合,剔除动植物残体和小石块,制备成1个混合土样带回实验室,部分于4 ℃保温箱保存,用于分析土壤酶活性;部分置于阴凉通风处风干,用于分析土壤化学性质。
1.4 样品分析
土壤pH:电位法(土水体积比1∶2.5);土壤有机碳:重铬酸钾—H2SO4氧化外加热法;土壤硝态氮:双波长紫外分光光度法;土壤氨态氮:KCl浸提—靛酚蓝比色法;土壤速效磷:NaHCO3浸提—钼锑抗比色法;土壤酶:土壤水解酶类活性的测定—对硝基酚底物比色法[26-27]。土壤酶测定方法:(1)土壤悬浊液的制备。土壤过2 mm筛,称取5 g鲜土于100 mL小白瓶中,加入100 mL pH 7.5缓冲液,在恒温摇床180 r·min-1室温震荡40 min,制成均匀的土壤悬浊液。(2)土壤样品测定。吸取2 mL土壤悬浊液和2 mL待测酶对应的底物溶液置于10 mL离心管中,在旋涡振荡器上充分混匀。每份土样做4个重复。将10 mL离心管置于恒温摇床170 r·min-1、25 ℃振荡培养。培养完毕后,将离心管放入低速冷冻离心机,在4 ℃下2 000 r·min-1离心5 min。吸取2 mL上清液置于装有0.2 mL 1 mol·L-1NaOH溶液的15 mL离心管中(在测定亮氨酸氨基肽酶时,不需要加NaOH,直接吸取上清液即可),加8 mL去离子水,并在旋涡振荡器上充分混匀。用去离子水调节分光光度计零点,在410 nm处测定吸光值。表2为所测定酶的种类、底物及具体功能。
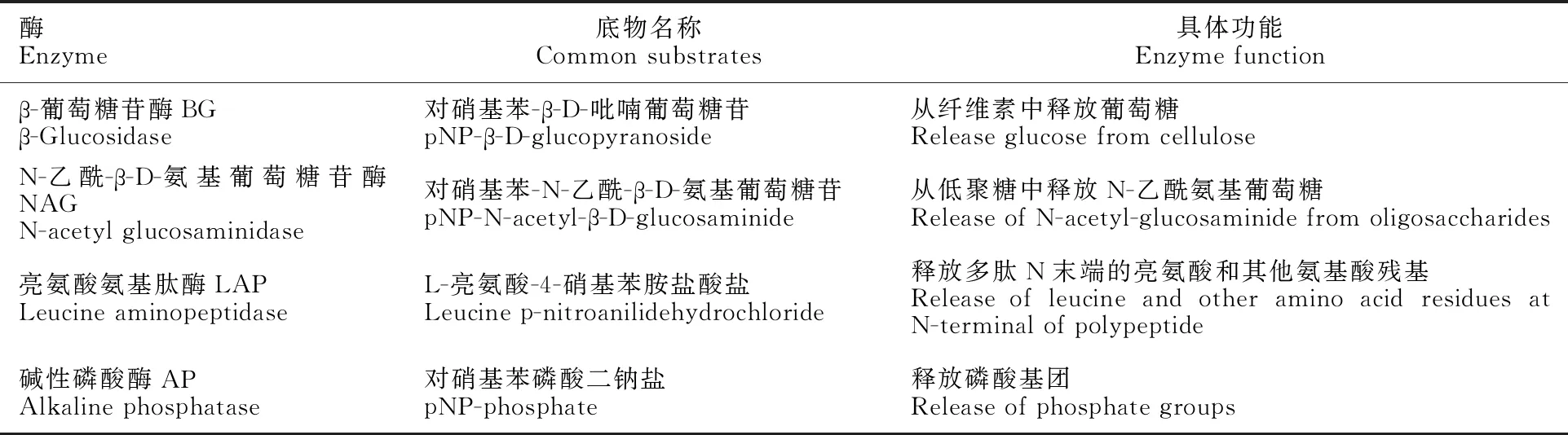
表2 不同酶对应的底物及具体功能
1.5 数据分析
利用 SigmaPlot 10.0(Systat 软件公司,伊利诺伊州,芝加哥)和 OriginPro 2021(OriginLab,北安普顿,马萨诸塞州,美国)进行绘图。采用 SPSS 26.0(IBM 公司,纽约州,美国)进行单因素方差分析(ANOV A),再进行Duncan’s差异显著性检验(P<0.05)。采用 Canoco 5统计软件对土壤性质进行 PCA(Principal component analysis)分析。
2 结果与分析
2.1 土壤pH的变化
与对照相比,施肥和免耕补播对土壤pH没有显著影响(P>0.05),施肥+免耕补播显著降低土壤pH (P<0.05),土壤更趋于中性,说明施肥+免耕补播能够有效改善人工草地土壤的酸碱问题(图2)。

不同字母表示差异显著 (P<0.05);下同
2.2 土壤养分分布特征
与对照相比,施肥显著促进土壤有机碳和铵态氮含量增加(P<0.05),且其含量高于免耕补播和施肥+免耕补播。而施肥、免耕补播和施肥+免耕补播对硝态氮均呈抑制作用,其中免耕补播处理下硝态氮含量显著低于对照和施肥+免耕补播处理(P<0.05),说明施肥可以通过促进硝化反应固定二氧化碳,从而进一步增加土壤有机碳含量。施肥+免耕补播对速效磷含量有显著促进作用(P<0.05),说明施肥+免耕补播明显加速高寒草甸土壤速效磷的合成积累 (图3)。
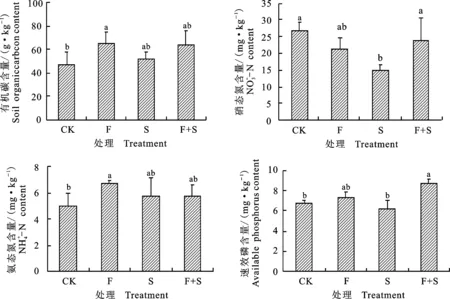
图3 不同人工修复措施处理后土壤养分分布特征
2.3 土壤酶分布特征
与对照相比,短期施肥和施肥+免耕补播对β-葡萄糖苷酶、N-乙酰-β-D-氨基葡萄糖苷酶、亮氨酸氨基肽酶和碱性磷酸酶活性均无显著影响(P>0.05)。免耕补播显著降低N-乙酰-β-D-氨基葡萄糖苷酶活性(P<0.05),对β-葡萄糖苷酶、亮氨酸氨基肽酶活性虽呈抑制作用,但与其他处理相比差异不显著(P>0.05),说明免耕补播措施下牧草氮素需求增加,土壤中氮素减少,进一步抑制N-乙酰-β-D-氨基葡萄糖苷酶活性。3种修复措施中,施肥+免耕补播处理下4种酶活性均为最高(图4)。
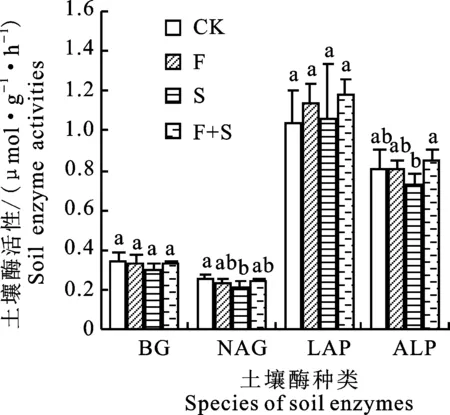
图4 不同人工修复措施处理后土壤酶分布特征
2.4 土壤养分和酶活性的相关性分析
相关性分析表明(表3),有机碳含量与亮氨酸氨基肽酶活性、硝态氮含量与β-葡萄糖苷酶活性和N-乙酰-β-D-氨基葡萄糖苷酶活性、β-葡萄糖苷酶活性与N-乙酰-β-D-氨基葡萄糖苷酶活性和碱性磷酸酶活性呈显著正相关关系(P<0.05),相关系数分别为0.947、0.963、0.973、0.966、 0.961。pH与速效磷含量呈显著负相关关系 (P<0.05),相关系数为-0.945。有机碳含量、氨态氮含量、速效磷含量、亮氨酸氨基肽酶和碱性磷酸酶活性等彼此间均呈正相关关系。pH与有机碳、硝态氮、氨态氮含量和β-葡萄糖苷酶、N-乙酰-β-D-氨基葡萄糖苷酶、亮氨酸氨基肽酶、碱性磷酸酶活性均呈负相关关系。
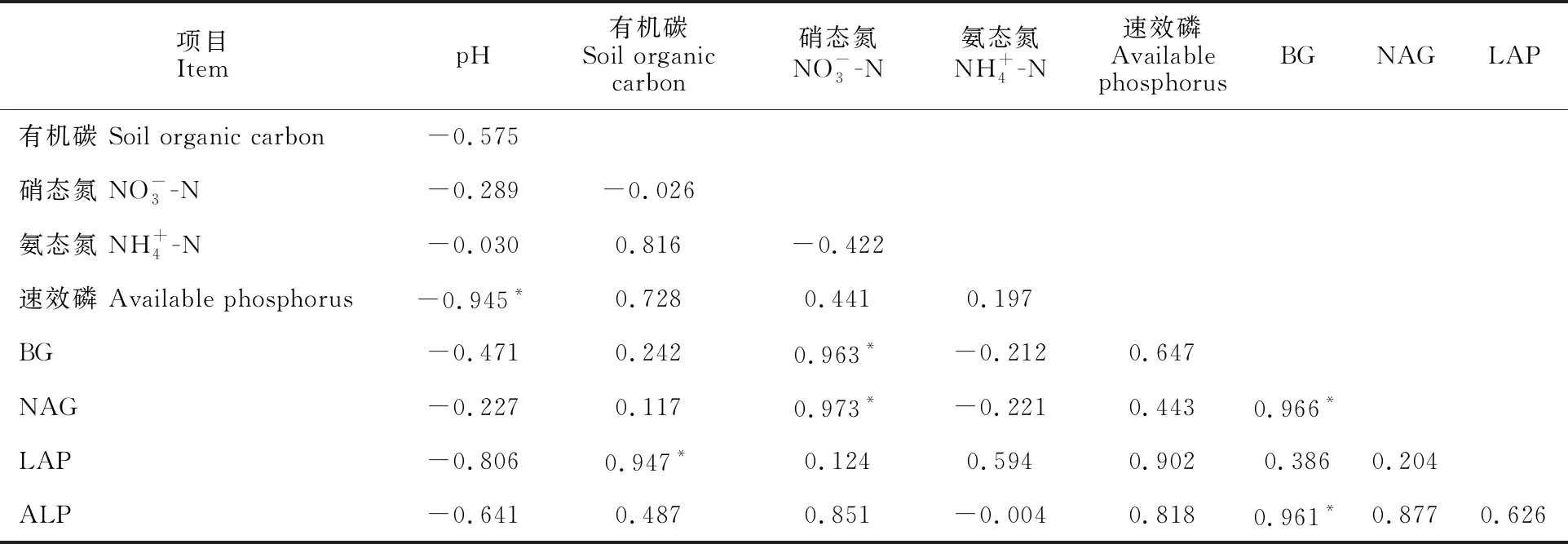
表3 不同处理后土壤性质 Pearson 相关分析
运用主成分分析(PCA)评价人工修复措施过程中土壤养分和土壤酶活性间的关系(图5),前两个轴(PC1,PC2)共同解释95.07%的变异,施肥+免耕补播处理在PC1轴上得分较高,施肥处理则在 PC2 轴得分较高,说明施肥+免耕补播对亮氨酸氨基肽酶和碱性磷酸酶酶活性及速效磷含量的维持贡献较大,而单施肥对有机碳和氨态氮含量贡献较大。

图5 不同人工修复措施处理后土壤性质主成分分析
3 讨 论
3.1 不同人工修复措施对退化草地土壤养分组成的影响
在草地生态系统中,土壤碳氮含量主要受气候变化、植被类型、土壤特性及人类活动的综合影响[28-29],从而导致土壤有机碳氮累积、分解和淋溶输出程度的不同[30]。天然草原土壤中的磷主要来源于矿物,在土壤中主要以与钙、镁、铁、铝及黏粒结合的无机态的形式存在[31],其含量主要受土壤母质和成土作用的影响[32]。另外,研究表明,海拔、土地利用方式及坡度等因素也可以通过影响气象因素或土壤侵蚀程度而间接影响土壤磷含量[33]。
有机质作为土壤中重要的组成成分,其本身含各种营养元素,还对土壤中各种肥力因素起着重要的调节作用[34]。本研究发现,施肥+免耕补播显著降低土壤pH,这可能是由于施肥和免耕补播缓解了高寒草甸的退化,降低了土壤水分的蒸发量,盐分不会聚表,导致土壤 pH和电导率逐渐降低。研究表明,土壤中施加有机物可以给微生物供给足够的底物,从而加速土壤原有有机碳的矿化,释放更多的活性碳组分促进土壤碳循环[35]。本试验中,施加有机肥短期内显著增加土壤有机碳含量,表明退化草地施加有机肥有利于提高有机碳含量,促使植物生长,这也证实了前人的研究结论[36]。姬万忠等[37]研究发现,补播3 a后土壤有机碳含量显著提高,这与本研究结果相反,免耕补播与施肥+免耕补播有机碳含量差异不显著的主要原因可能是试验年限较短,这还需要长时间的试验来探究。
土壤氮素和磷素是衡量土壤肥力和植被生产力的重要指标。铵态氮是有机氮向硝态氮转化过程中的中间产物,是土壤供氮能力的重要衡量指标[38],施入氮肥可以增加土壤铵态氮的供给,本研究结果表明,施肥能显著增加土壤氨态氮含量。土壤对硝态氮的吸附能力弱,易于淋失[38],土壤硝态氮主要集中在0~20 cm土层内。本试验结果表明免耕补播能显著降低土壤硝态氮含量,这与相关学者研究结果相反[37, 39],可能是由于免耕补播年限较短。施肥+免耕补播显著增加土壤速效磷含量,这与王伟等[40]研究结果一致,说明施肥+免耕补播对改良高寒草甸土壤速效磷含量具有明显的作用。
3.2 不同人工修复措施对退化草地土壤酶活性的影响
土壤酶作为土壤生态系统中进行生化反应的催化剂,参与土壤一系列复杂的生化过程,包括合成和分解腐殖质、分解动植物残体、固定和释放土壤养分及各种氧化还原反应等[41]。土壤酶不仅对土壤物质转化和能量流动有催化作用,而且对土壤中的多种有机物质及残体产生各种生命化学转化,并与土壤有机碳分解速率和有机碳库的周转模式密切相关,使土壤生态系统各组分在功能上有一定的联系,最终保持土壤的生物化学稳衡状态[41]。
本研究测定与土壤有机碳、氮和磷循环密切相关的4种土壤水解酶活性。相关研究发现,施肥对土壤酶活性的影响因土壤条件和肥料类型与用量不同而差异明显,Wang 等[42]研究发现在半干旱草原土壤,化肥氮投入增加对土壤团聚体 N-乙酰-β-D-氨基葡萄糖苷酶活性有促进作用,但是抑制β-葡萄糖苷酶活性。而本研究发现,施肥、免耕补播及施肥+免耕补播均降低β-葡萄糖苷酶和N-乙酰-β-D-氨基葡萄糖苷酶活性,并且在免耕补播时活性最低,可能是由于施肥种类不同,免耕补播等处理试验年限较短,短期内效果不明显。朱敏等[43]认为长期施用有机肥对微生物群落结构无明显作用,表面施用有机肥对土壤酶活性无显著差异。本研究结果表明,β-葡萄糖苷酶和亮氨酸氨基肽酶活性在不同人工修复措施间差异均不显著,而碱性磷酸酶活性在免耕补播与施肥+免耕补播差异显著。
3.3 土壤理化性质和酶活性的相关性
通过主成分分析得出土壤理化性质与酶活性间紧密联系,并存在一定的相关性。研究结果发现,有机碳含量与亮氨酸氨基肽酶、硝态氮含量与β-葡萄糖苷酶活性和N-乙酰-β-D-氨基葡萄糖苷酶活性有显著正相关关系,表明有机碳对亮氨酸氨基肽酶活性、硝态氮对β-葡萄糖苷酶活性和N-乙酰-β-D-氨基葡萄糖苷酶活性的影响最大,说明酶对土壤碳、氮养分变化敏感,可作为衡量土壤肥力的生物学指标[44]。pH与速效磷含量呈显著负相关关系,说明当pH升高时,微生物会调整自身的生长策略,相对更偏向于选择聚集更多的磷用于自身生长,因此,改善当地土壤碱化的趋势可能对于缓解磷限制有一定作用。本研究中,酶主要受有机碳和硝态氮含量的影响,说明土壤碳、氮养分是影响生态酶的重要因素。
4 结 论
施肥显著增加土壤有机碳和氨态氮含量,免耕补播显著降低土壤硝态氮含量,施肥+免耕补播对速效磷含量有显著促进作用。施肥通过增加土壤碳、氮养分来影响土壤酶活性,施肥+免耕补播处理效果最好,酶活性达到最大。
猜你喜欢
中草药(2022年20期)2022-11-15
食品科学(2022年20期)2022-10-31
现代食品科技(2022年9期)2022-10-09
农药学学报(2022年3期)2022-06-14
水土保持学报(2022年3期)2022-05-26
节能与环保(2022年3期)2022-04-26
安徽农学通报(2022年6期)2022-04-07
科技创新与应用(2022年6期)2022-03-24
中国生态农业学报(中英文)(2022年1期)2022-01-15
丝路视野(2018年15期)2018-05-14
