
番木瓜WRKY转录因子家族基因鉴定及其响应短孢炭疽菌侵染的表达分析
2022-05-16 04:54周陈平邝瑞彬黄炳雄魏岳荣
西北农林科技大学学报(自然科学版) 2022年5期
杨 敏,周陈平,杨 护,邝瑞彬,黄炳雄,魏岳荣
(广东省农业科学院果树研究所,农业农村部南亚热带果树生物学与遗传资源利用重点实验室, 广东省热带亚热带果树研究重点实验室,广东 广州510640)
番木瓜(CaricapapayaL.)是我国岭南特色水果,具有很高的营养价值和药用价值,随着人们对其认可程度的提高,需求量持续增加,已成为世界上第四大热带、亚热带畅销水果[1-3]。然而,番木瓜采后果实易受炭疽菌侵染而影响贮运和货架期,导致商品果损失,损失率可高达50%[4],极大地限制了番木瓜产业的发展。防治番木瓜炭疽病的传统方法是理化处理,该方法虽有一定效果,但需投入巨大的人力、物力,且影响环境[5-6],而通过分子育种获得抗炭疽病番木瓜品种是防治炭疽病最根本、最有效的方法。因此准确筛选抗病候选基因是番木瓜抗炭疽病分子育种的关键环节。大量研究表明,转录因子尤其是WRKY转录因子在植物抗病中发挥着关键作用[7-9]。
WRKY转录因子家族是植物中特有的最大转录因子家族之一,WRKY蛋白的共同特征是含有约60个氨基酸的保守结构域,包含位于N端的七肽(WRKYGQK)WRKY结构域和位于C端的锌指结构,其中N端七肽序列为高度保守的核心序列,C端由C2H2型(CX4CX22-23HXH/C)或C2HC型(CX7CX23HXC)锌指结构组成[10-11]。根据结构域特征及聚类分析,可将WRKY转录因子家族分为Ⅰ、Ⅱ、Ⅲ 3大类,其中Ⅰ类的锌指结构为C2H2,含 2 个 WRKY 结构域;Ⅱ类的锌指结构为C2H2,含 1 个 WRKY 结构域,根据 WRKY 转录因子结构可进一步分为Ⅱ a、Ⅱ b、 Ⅱ c、Ⅱ d 和Ⅱe等5个亚类; Ⅲ类家族锌指结构为 C2HC,含 1 个 WRKY 结构域[10,12-13]。WRKY转录因子可参与植物生长发育及多种生物和非生物胁迫,介导植物防御调节功能,尤其在真菌、细菌和病毒病原体侵染等生物胁迫防御中发挥着重要作用。在拟南芥(Arabidopsisthaliana)中,WRKY33可以正调控植株对腐生型病原菌黑斑病菌(Alternariabrassicicola)引起的黑斑病和灰霉菌(Botrytiscinerea)引起的灰霉病的防御作用[14-15],WRKY18和WRKY40可以负调控植株对活体营养型真菌白粉病菌(Golovinomycesorontii)的防御作用[16],WRKY3和WRKY4可以正调控植株对腐生性真菌B.cinerea的抗性[17]。水稻(Oryzasativa)转录因子OsWRKY22过量表达时可增强植株对半活体营养型病原菌稻瘟病菌(Magnaportheoryzae)的抗性[18]。小麦(Triticumaestivum)TaWRKY70可正向调控高温条件下幼苗对活体营养型小麦条锈菌(Pucciniastriiformisf.sp.Tritici,Pst)的抗性[19]。葡萄(Vitisvinifera)在受到白腐病菌(Coniothyriumdiplodiella)胁迫后,57%的WRKY转录因子表达量会发生变化[20]。在百合(Liliumlongiflorum)中也鉴定到多个WRKY转录因子,这些转录因子可能参与了百合对灰霉病病菌(B.elliptica)侵染的防御反应[21]。烟草(Nicotianabenthamiana)NtWRKY7、NtWRKY8、NtWRKY9和NtWRKY11能够与RBOHB基因启动子中的W-box结合,通过ROS途径激活免疫反应[22]。
番木瓜WRKY转录因子的鉴定虽有报道[23],但分析相对简单,到目前为止,番木瓜WRKY家族在对病原菌侵染,尤其是在半活体营养型真菌炭疽菌侵染响应中的作用还未见报道。笔者前期对炭疽菌侵染易感和耐病番木瓜品种进行了转录组测序与分析,基于这些数据,本研究利用生物信息学方法鉴定番木瓜WRKY(CpWRKY)转录因子家族基因,并对其基因结构、蛋白保守结构域、系统进化、启动子顺式作用元件及炭疽菌侵染下CpWRKYs基因表达量进行分析,以期为CpWRKYs家族的功能研究和利用奠定基础。
1 材料与方法
1.1 材 料
番木瓜果实,采自广东省农业科学院果树研究所标准化种植的炭疽病耐性品种‘G20’和易感品种‘Y61’2年生果树。短孢炭疽菌(C.brevisporum),本课题组参照Duan等[24]的方法分离和鉴定。
1.2 CpWRKY转录因子家族的鉴定
CpWRKY转录因子家族的蛋白序列下载于Plant TFDB V4.0 数据库[25],将这些成员利用在线软件SMART (http:// smart.embl-heidelberg.de/) 和 InterProScan工具 ( http://www.ebi.ac. uk/Tools/pfa/iprscan/) 进行筛选和鉴定。
1.3 CpWRKY生物信息学分析及进化树构建
利用ProPAS 软件对筛选获得的CpWRKY家族各成员的理化性质进行分析[26]。从Phytozome数据库(https://phytozome.jgi.doe.gov/pz/portal.html)中下载CpWRKY家族基因成员的基因组序列、ID号和编码序列(CDS),通过Gene Structure Display Server (GSDS) v2.0确定内含子、外显子数量及基因结构[27]。
在Uniport蛋白质数据库(https://www.uniprot.org/)中下载拟南芥和番木瓜WRKY转录因子家族蛋白,利用MAFFT v7.471进行比对,基于邻接法使用FastTreeFastTree v2.1.11构建系统进化树,并使用iTOL v6(https://itol.embl.de/)对系统发育进化树进行美化。对获得的48个CpWRKY转录因子蛋白序列利用MAFFT v7.471进行多序列比对,用jalview对CpWRKY蛋白序列进行展示。利用在线软件Multiple EM for Motif Elicitation (MEME, version 5.02) (http://meme-suite.org/tools/ meme)进行保守基序预测[28]。在植物基因组网站(http://plants.ensembl.org/index.html)下载CpWRKY各成员起始密码子上游2 000 bp的启动子序列,通过软件PlantCARE[29](http//bioinformatics.psb.ugent.be/webtools/plantcare/html/)进行顺式作用元件在线预测和分析。
1.4 短孢炭疽菌侵染后CpWRKY家族基因的表达分析
1.4.1 差异表达基因的筛选 从NCBI网站下载转录组数据(NCBI登录号:PRJNA692338),分析番木瓜采后果实在短孢炭疽菌侵染后CpWRKY家族基因的表达情况,炭疽菌侵染后CpWRKYs基因的表达量为处理样品的表达量减去对照样品的表达量。以|lb差异倍数|≥1和P<0.05为表达量明显变化的判定标准筛选差异表达基因。
1.4.2 差异表达基因的验证 以真核翻译起始因子4A(Eukaryotic initiation factor 4A,EIF4A)为内参基因[30],对5个在‘G20’中特异表达的CpWRKY基因(CpWRKY22、CpWRKY11、CpWRKY25、CpWRKY33和CpWRKY2)进行RT-qPCR检测。利用Primer Premier 5.0软件设计引物(表1)。
将采摘的无机械外伤、成熟度一致的番木瓜品种‘G20’和‘Y61’(‘G20’和‘Y61’分别为NCBI网站转录组数据所用的耐炭疽病和易感炭疽病番木瓜品种)果实用体积分数75%酒精冲洗,之后再用无菌水冲洗,室温晾干后,利用针刺法在果实相同部位接种短孢炭疽菌(接种菌液的细菌浓度为106mL-1),在接种0(对照),24和48 h时取接种点周围半径约2 cm圆内果肉样品,备检。用RNA 提取试剂盒(TaKaRa)提取样品总RNA,利用DNase Ⅰ (TaKaRa)去除基因组DNA,根据试剂盒说明,用反转录酶(TaKaRa)将1 μg RNA 反转录成cDNA。以cDNA为模板,用Applied Biosystems 7500实时定量PCR仪进行RT-qPCR试验。反应体系为:2×SYBR Green ProTaqHS Premix 10 μL,正向引物F(10 μmol/L) 0.4 μL,反向引物R(10 μmol/L) 0.4 μL,cDNA 模板 2 μL,50×ROX Reference Dye 0.4 μL,ddH2O 6.8 μL,总体积20 μL。两步法扩增程序为:95 ℃ 30 s;95 ℃ 5 s,56 ℃ 30 s,40 个循环。熔解曲线标准程序:95 ℃ 15 s,60 ℃ 1 min,95 ℃ 15 s。试验重复3次,利用2-ΔΔCt方法计算基因相对表达量。
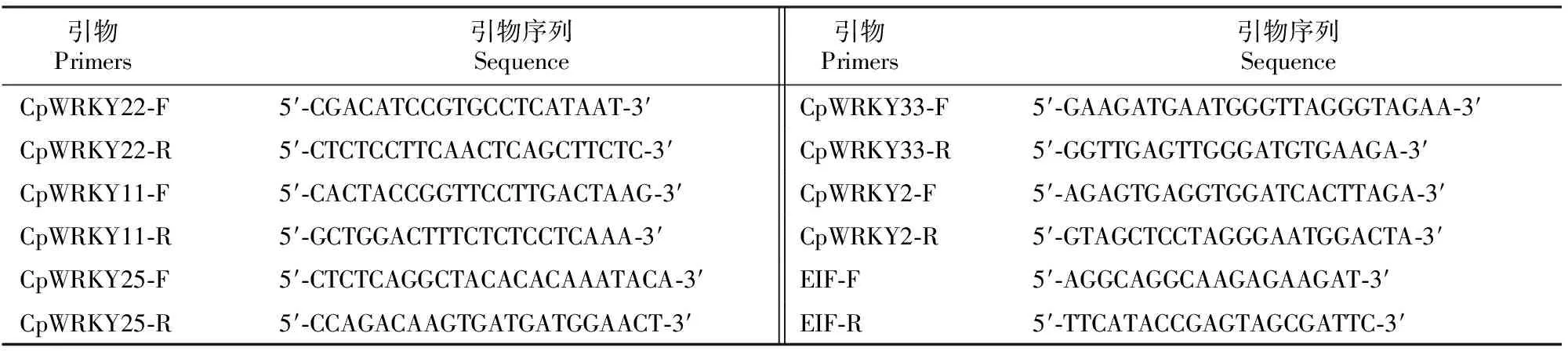
表1 RT-qPCR试验所用引物Table 1 Primers used for RT-qPCR in this study
2 结果与分析
2.1 CpWRKY家族的鉴定和理化性质分析
从Plant TFDB V4.0数据库下载共获得49个CpWRKY蛋白序列,用SMART在线软件和InterProSca工具进一步鉴别和筛选,发现除基因号为evm.model.supercontig_1195.2的蛋白外,其余48个都有WRKY结构域,即番木瓜WRKY转录因子家族共有48个成员,将其命名为CpWRKY1~CpWRKY48。理化性质分析结果(表2)显示,CpWRKY1~CpWRKY48蛋白的氨基酸数为97~747个,分子质量在10 867.25~80 553.73 u,等电点(pI)为4.83~9.71,均为亲水性蛋白,亲水性平均值均为负值,为-0.54~―1.08。
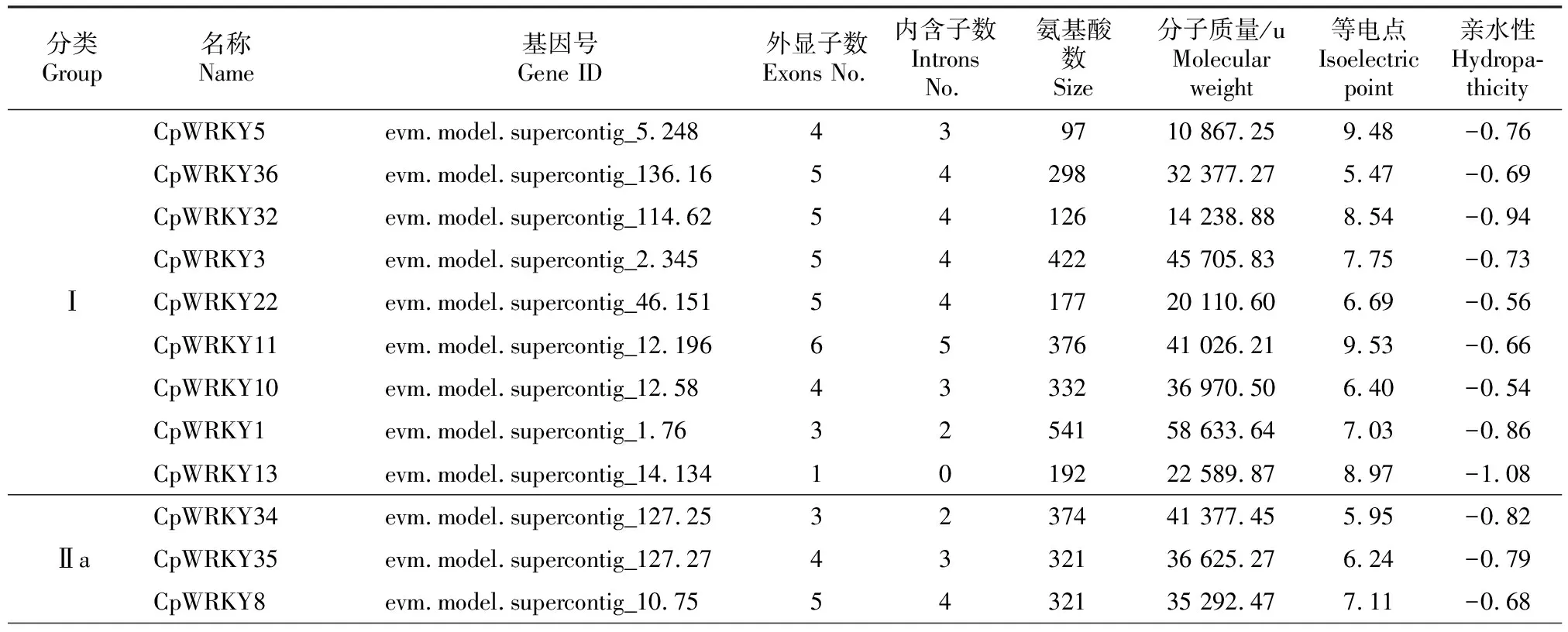
表2 CpWRKYs家族成员信息Table 2 Information of CpWRKY family
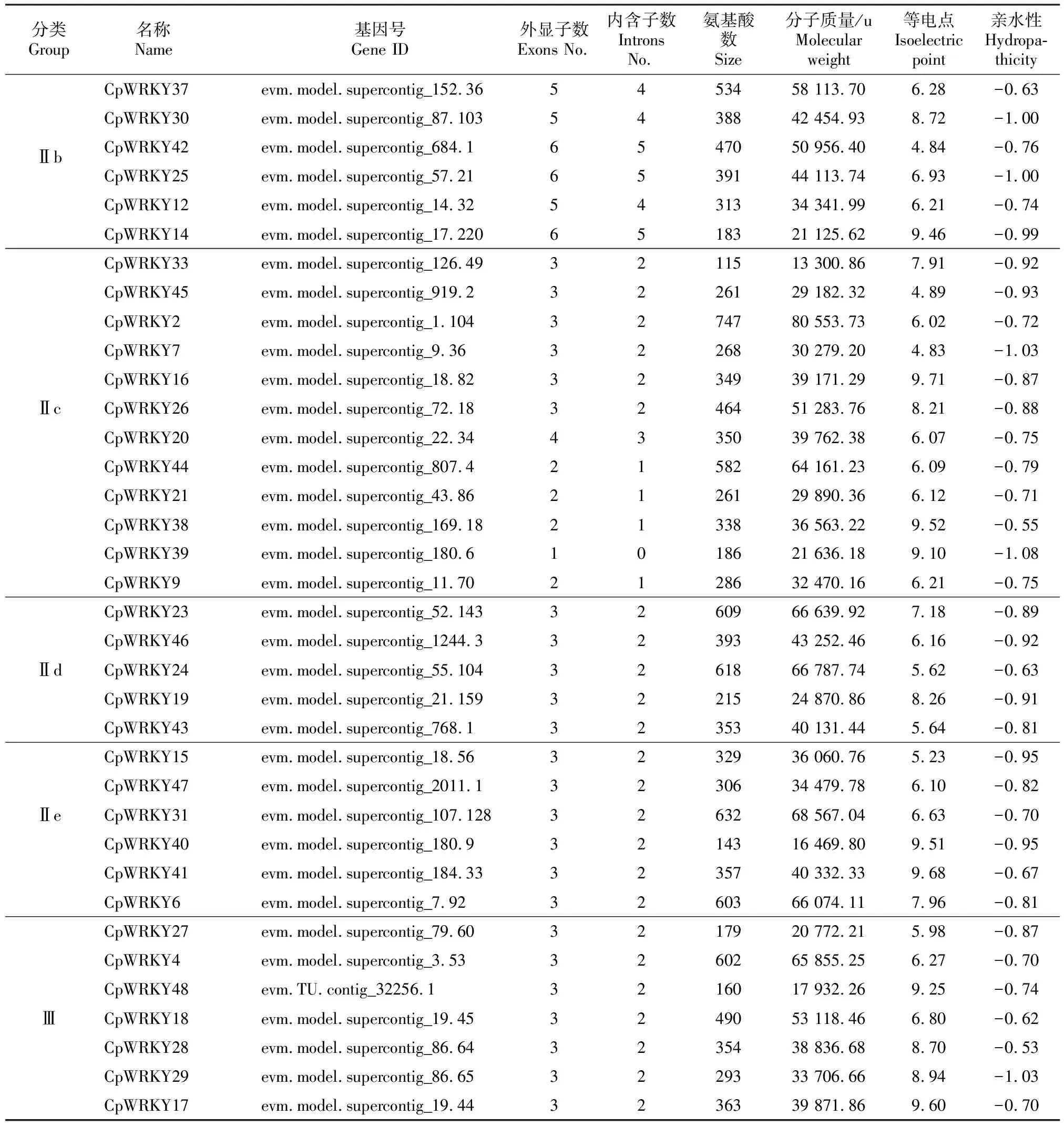
表2(续) Conutined table 2
2.2 CpWRKYs家族分类与保守结构域的序列分析
为了对48个CpWRKY 蛋白进行准确分类,按照之前报道的WRKY蛋白分类方法[10,12-13],引入拟南芥WRKY(AtWRKY)蛋白序列进行系统发育进化分析,结果表明,CpWRKY家族可分为Ⅰ、Ⅱ和Ⅲ 3大类,其中Ⅰ和Ⅲ类分别有9和7个成员,第Ⅱ大类成员数最多,有32个(占总数的67%),且第Ⅱ大类CpWRKY又可分为5个亚类:Ⅱ a、Ⅱ b、Ⅱ c、Ⅱ d和Ⅱ e,分别包括3,6,12,5和6个成员(图1,表2)。
同时,对CpWRKY的保守结构域进行多序列比对和分析,结果(图2)显示,Ⅰ类CpWRKY的9个成员中,除CpWRKY22、CpWRKY11、CpWRKY13外,其余6个均含有2个WRKY七肽(WRKYGQK)结构域,分别位于蛋白的N端(Ⅰ-N)和C端(Ⅰ-C),WRKY七肽结构域高度一致,C2H2锌指结构的保守性稍弱。Ⅱ类CpWRKY的32个成员中,除CpWRKY9、CpWRKY41外,其余30个成员都包含1个WRKY七肽结构域,其中Ⅱa、Ⅱ b、和Ⅱ d 3个亚类的保守结构域(WRKY七肽结构域和C2H2锌指结构)较为保守,Ⅱc和Ⅱe的氨基酸序列结构域存在较多变异。Ⅲ类CpWRKY的7个成员均具有保守的WRKY七肽结构域和C2HC锌指结构。
2.3 CpWRKYs基因结构及其蛋白保守基序分析
基因结构多样性可以反映多基因家族的演化,对48个CpWRKY基因的结构分析结果显示,CpWRKYs由1~6个外显子组成,其中CpWRKY13和CpWRKY39没有内含子,只由1个外显子组成;CpWRKY9、CpWRKY21、CpWRKY38和CpWRKY44由1个内含子和1个外显子组成。同一家族/亚家族的成员也表现出相似的基因结构特征,CpWRKYs Ⅱd和Ⅱe亚家族基因都含有2个内含子和3个外显子,Ⅲ类CpWRKYs均由2个内含子和3个外显子组成(图3,表2),即CpWRKYs基因结构呈现组间差异性和组内保守性的特征。

蓝色为疏水性氨基酸,红色为正电荷氨基酸,紫色为负电荷氨基酸,绿色为极性氨基酸,粉色为半胱氨酸,橙色为甘氨酸,黄绿色为脯氨酸,蓝绿色为芳香族氨基酸,无配色为非保守性氨基酸。黑色框为WRKY七肽(WRKYGQK)结构域,蓝色框为C2H2锌指结构域,黄色框为C2HC锌指结构域The blue font represents hydrophobic amino acids,the red font represents positively charged amino acids,the purple font represents negatively charged amino acids,the green font represents polar amino acids,the pink font represents cysteine, the orange font represents glycine,the yellow green font represents proline,and the blue-green font represents aromatic amino acids,and unmatched fonts represent non conservative amino acids.The amino acids in the black frame are WRKY heptapeptide (WRKYGQK) domain,the amino acids in the blue frame are C2H2 zinc finger domain,and the amino acids in the yellow frame are C2HC zinc finger domain
本研究利用MEME在线软件对48个CpWRKY蛋白进行基序分析,共预测到10个保守基序(图3和图4),这些基序的长度为17~50个氨基酸,其中基序1、2和3包含WRKY保守结构域,基序1包含七肽保守序列,基序2包含锌指结构,基序3既包括七肽序列又包括锌指结构(图4)。如图3所示,CpWRKYs各大类蛋白成员的基序比较一致,但各亚类之间存在一定差异。48个CpWRKY蛋白中有21个含有基序4-基序1-基序2的基序串,占总蛋白数的42.50%。除此之外,Ⅰ类蛋白多数还包含基序3,位于基序4-基序1-基序2基序串的前面,这与Ⅰ类WRKY蛋白含有2个WRKY结构域的结果一致。Ⅱ类和Ⅲ类CpWRKY蛋白都含有基序1-基序2基序串,即包含1个保守七肽序列和1个锌指结构域,这也与上文的结果一致。Ⅱ类CpWRKY蛋白除具有典型的WRKY结构域基序外,Ⅱa亚类还含有基序5和基序7,其中CpWRKY34还包含了基序6;Ⅱb亚类还含有基序5、6、7和9;Ⅱd亚类还含有基序8和10。结果表明,Ⅱa、Ⅱb和Ⅱd亚类的基序更加丰富多样。
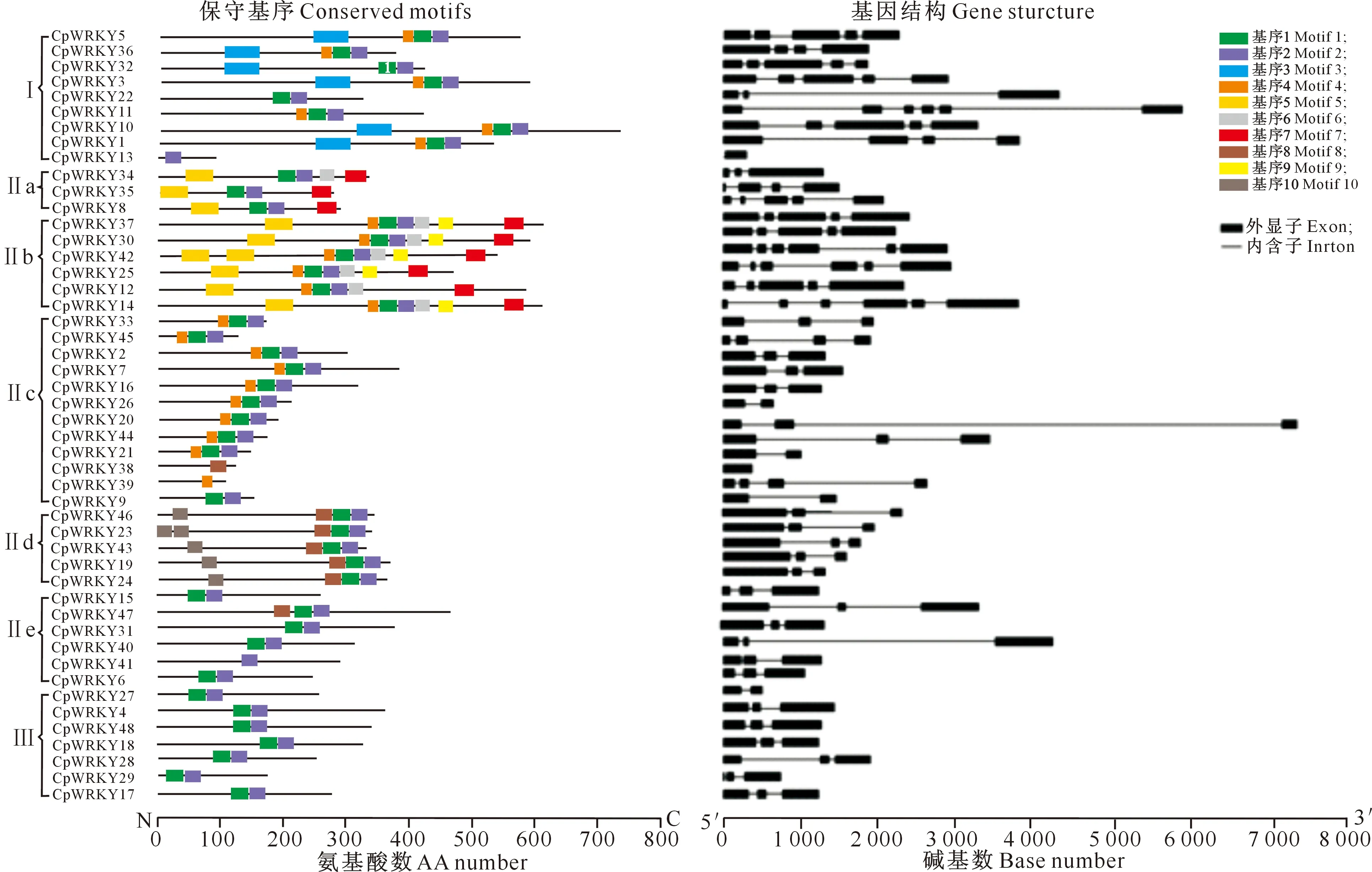
图3 CpWRKYs家族蛋白保守基序及其基因结构Fig.3 Conserved motifs of CpWRKYs and gene structure of CpWRKYs
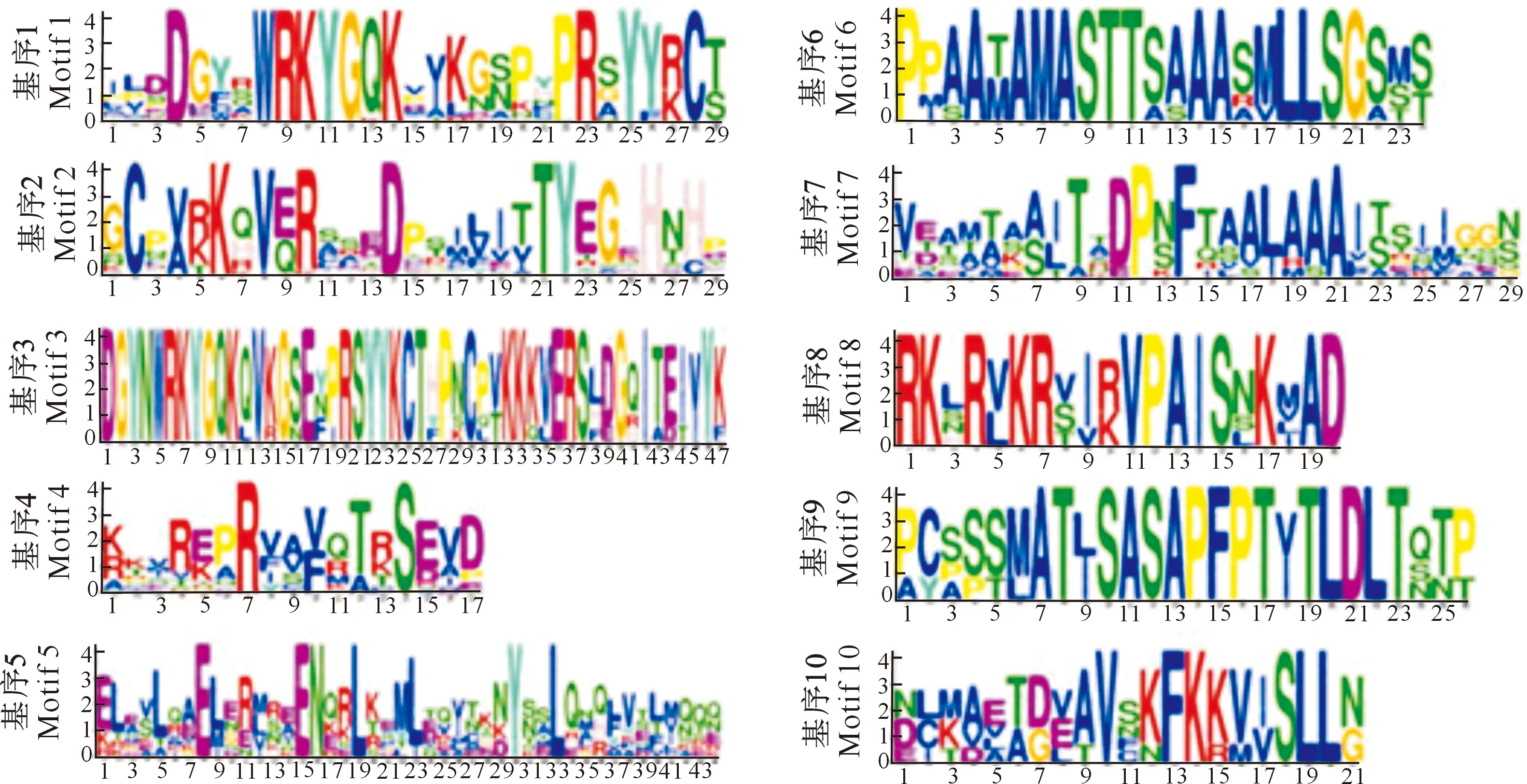
基序1和3包含WRKY七肽保守序列,基序2和3包含锌指结构Motifs 1 and 3 contain conserved WRKY heptopeptides,and motifs 2 and 3 contain zinc finger domains图4 CpWRKYs的保守基序Fig.4 Conserved motifs of CpWRKYs
2.4 CpWRKYs启动子顺式作用元件分析
为了进一步研究CpWRKYs的功能和调控模式,利用PlantCARE对CpWRKYs起始密码子上游2 000 bp 启动子区域进行分析,共检测到514个与生物和非生物胁迫相关的元件,包括33个GA响应元件(15个P-box、13个GARE-motif、5个TATC-box),占比6.42%;27个生长素响应元件(3个AuxRR-core、21个TGA-element、3个TGA-box),占比5.25%;162个MeJA响应元件(81个CGTCA-motif 和 81个TGACG-motif),占比31.52%;24个SA响应元件(23个TCA-element和1个SARE),占比4.67%;158个ABA响应元件(ABRE),占比30.74%;31个伤害响应元件(WUN-motif),占比6.03%;19个防御胁迫元件(TC-rich repeats),占比3.70%;26个低温及其他非生物胁迫元件(1个DRE和25个LTR),占比5.06%。结果暗示,CpWRKYs广泛参与番木瓜的生物胁迫与非生物胁迫过程。
2.5 短孢炭疽菌侵染后CpWRKY转录因子家族的表达分析
利用前期研究所得转录组数据,进行基因表达差异性分析,结果(表3)显示,在短孢炭疽菌侵染‘G20’和‘Y61’果实48 h内,与0 h各自对照相比,表达差异的CpWRKY基因分别为37和36个,共同差异表达的有32个,这32个CpWRKY基因在侵染后的‘G20’采后果实中表达量均上调;在‘Y61’果实中有29个表达量上调,有3个(CpWRKY44、CpWRKY19和CpWRKY42)表达量下调。另外,短孢炭疽菌侵染后,在‘G20’和‘Y61’果实中特异表达的CpWRKY基因分别为5个(CpWRKY22、CpWRKY11、CpWRKY2、CpWRKY33和CpWRKY25)和4个(CpWRKY1、CpWRKY39、CpWRKY32和CpWRKY46),且‘G20’中5个CpWRKY基因的表达量全部上调;‘Y61’中的CpWRKY1和CpWRKY39表达量上调,而CpWRKY32和CpWRKY46表达量下调。可见短孢炭疽菌侵染后,‘G20’与‘Y61’之间差异表达的CpWRKY基因有12个,这些CpWRKY类型丰富,其中Ⅰ类4个(CpWRKY1、CpWRKY32、CpWRKY11、CpWRKY22),Ⅱ b类2个(CpWRKY42、CpWRKY25),Ⅱ c类4个(CpWRKY33、CpWRKY2、CpWRKY44、CpWRKY-39),Ⅱ d类2个(CpWRKY46、CpWRKY19)。
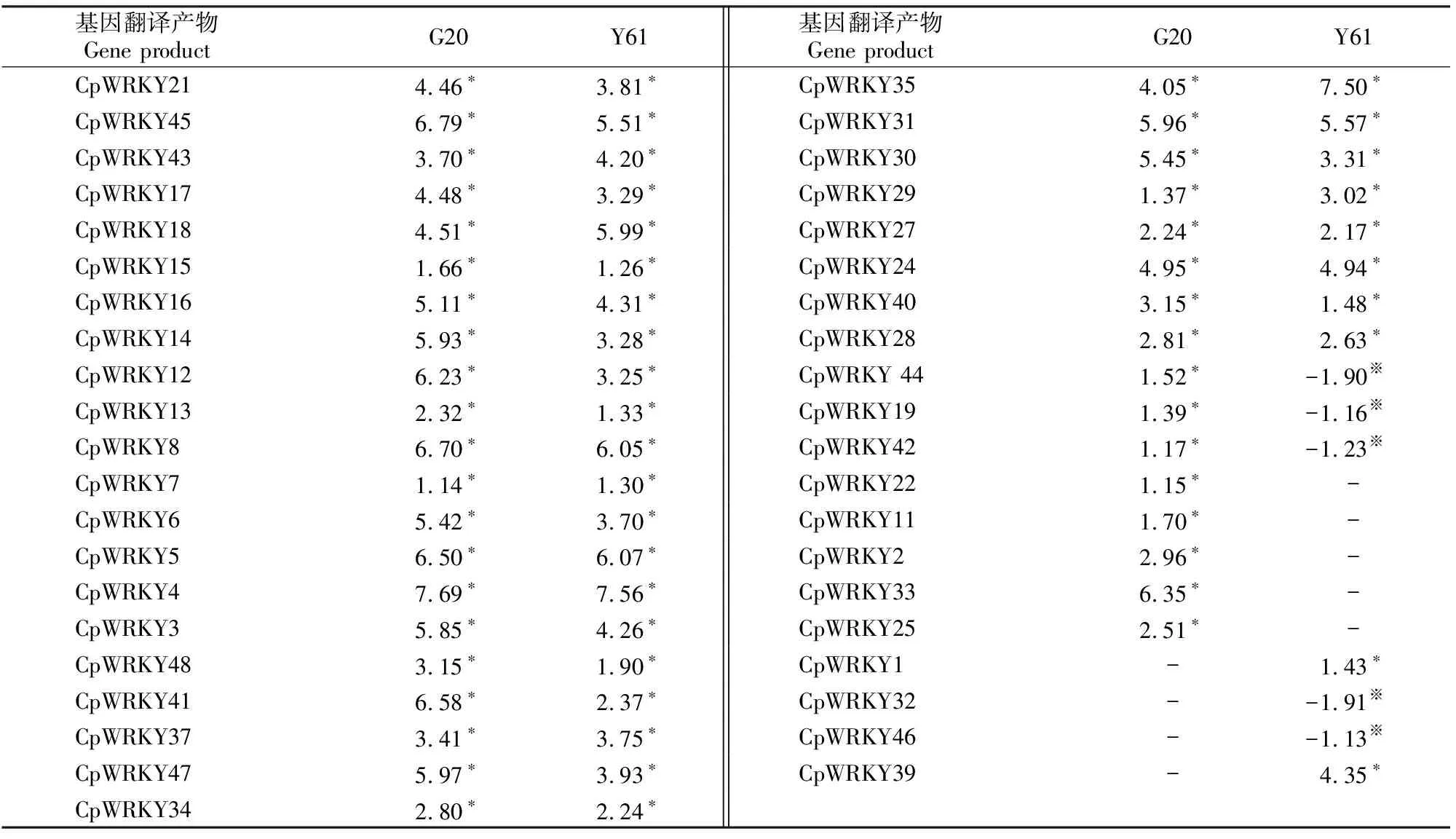
表3 炭疽菌侵染48 h内CpWRKY基因的表达量变化Table 3 Differential expression of CpWRKYs in G20 and Y61 at 48 h after C.brevisporum infection
2.6 短孢炭疽菌侵染后5个CpWRKYs在‘G20’和‘Y61’中表达量的变化
图5显示,在短孢炭疽菌接种‘G20’和‘Y61’果实后,CpWRKY22、CpWRKY11、CpWRKY2、CpWRKY33和CpWRKY25表达量在‘Y61’中无明显变化,而在‘G20’中表达量均显著上调,其中CpWRKY33上调了10倍以上,其他4个基因也均上调2倍以上。说明这5个WRKY基因可能是参与‘G20’抵抗炭疽菌侵染的重要候选基因。

与对照(0 h)相比,同一品种同一基因图柱上标*表示差异显著(P<0.05),标**表示差异极显著(P<0.01)Compared with the control (0 h), * on the same gene column of the same cultivar indicates significant difference at 0.05 level (P<0.05),and * * indicates extremely significant difference at 0.01 level (P<0.01) 图5 短孢炭疽菌侵染后5个重要候选CpWRKY在‘G20’和‘Y61’中表达水平的变化Fig.5 Changes in expression levels of five candidate CpWRKY genes in G20 and Y61 after C. brevisporum infection
3 讨 论
番木瓜为我国岭南特色水果,是支撑华南地区农业供给侧结构性改革和乡村振兴战略实施的重要特色经济树种。然而,番木瓜采后果实易受炭疽菌侵染,影响果实品质和货架期,严重制约番木瓜产业的发展。炭疽菌属于半活体生物营养型真菌,其中胶胞炭疽菌(Colletotrichumgloesporioides)作为主要的番木瓜炭疽病致病菌已被广泛报道,而短孢炭疽菌(Colletotrichumbrevisporum)是近年来新发现的炭疽病致病菌,现已在我国广泛流行,能够导致番木瓜、苦瓜、豆类、辣椒等经济作物炭疽病,严重影响作物品质和产量[24,31-33]。但目前对番木瓜炭疽病的侵染及抗性机理尚不明确,防控主要采取理化方法,挖掘番木瓜抗炭疽病关键候选基因,利用分子育种方法选育抗病新种质是解决该问题最有效的手段。
在植物对抗病原体的防御反应中,需要大规模的转录重编程,而转录重编程离不开转录因子的作用,其中WRKY转录因子在与病原体防御相关的早期反应中具有重要作用[34]。因此,鉴定番木瓜WRKY家族基因,并研究和筛选参与炭疽病抗性的重要基因,对于番木瓜抗炭疽病分子育种具有重要意义。本研究从番木瓜全基因组中筛选获得了48个WRKY转录因子家族成员,与拟南芥[10](72个)、水稻[35](102个)、枣树[36](54个)、甘薯[37](82个)、菠萝[38](54个)、高粱[39](94个)、苹果[40](116个)等植物相比,CpWRKY家族较小,这可能与番木瓜的参考基因组较小(只有372 Mb)有关[41]。48个CpWRKY家族成员被分为Ⅰ、Ⅱ和Ⅲ 3大类,其中Ⅱ类又被分为5个亚类。从蛋白数量上来看,Ⅱ类CpWRKY成员最多,其次为Ⅱ类。Ⅱ类CpWRKY各亚类中,Ⅱc的成员最多,有12个,占家族成员总数的25.00%。这与前人对拟南芥[10]、甘薯[37]、苹果[40]和草莓[42]的研究结果一致。基因结构及基序分析结果显示,CpWRKY家族在基因结构和保守基序组成上存在组间差异,但组内具有相似性,即各类CpWRKY之间内含子、外显子数目及基因结构差异较大,但同一类或同一亚类成员之间又有一定的相似性,如Ⅱd和Ⅱe亚家族都含有2个内含子和3个外显子,Ⅲ类CpWRKY均由2个内含子和3个外显子组成;在保守基序组成上也有类似特点,各类CpWRKY既具有一致的保守域,又有比较丰富的其他基序。这些特点与WRKY功能的多样性相一致。顺式作用元件分析结果显示,CpWRKY启动子序列中有大量参与生物胁迫与非生物胁迫的相关元件,暗示CpWRKY广泛参与这些过程,这与低温、干旱、伤害、PRSV等胁迫可诱导CpWRKY基因表达的研究结果[23]一致。
本研究发现,炭疽菌侵染后番木瓜耐性品种‘G20’和敏感品种‘Y61’中分别有37和36个CpWRKY基因表达量发生了变化,分别占CpWRKY家族总数的77.08%和75.00%,说明CpWRKY广泛参与炭疽病胁迫过程,这与葡萄经白腐病胁迫后,超过57%的WRKY表达量发生变化的结果[20]一致。本研究进一步分析发现,‘G20’和‘Y61’中有29个相同的CpWRKY基因被炭疽菌诱导上调表达,但表达强度有差异,且这些基因中包括大量参与植物免疫反应基因的同源基因。如CpWRKY45的同源基因WRKY51参与拟南芥茉莉酸诱导的防御反应[43];CpWRKY18的同源基因WRKY55可通过调控拟南芥活性氧和水杨酸生物合成相关基因的转录,正向调控拟南芥叶片衰老和防御反应[44];CpWRKY12的同源基因WRKY72参与番茄和拟南芥的基础免疫以及番茄R基因Mi-1介导的基因对基因的抗性[45];CpWRKY8的同源基因WRKY40参与拟南芥镰刀菌胁迫[46];CpWRKY40的同源基因WRKY29在拟南芥先天免疫反应和对小麦赤霉病抗性的免疫途径中发挥重要作用[47-48]。结果表明,这些基因应是广泛参与番木瓜炭疽菌侵染响应过程的基因,其表达强度不同可能是导致‘G20’和‘Y61’抗病性不同的原因之一。
本研究还发现,‘Y61’中有CpWRKY1、CpWRKY32、CpWRKY39和CpWRKY46等4个特异表达的基因,仅CpWRKY1的同源基因WRKY4有参与植物抗病过程的报道[49];而‘G20’中有CpWRKY22、CpWRKY11、CpWRKY2、CpWRKY33和CpWRKY25等5个特异表达基因,均在炭疽菌侵染后上调表达,除CpWRKY22外,其余4个的同源基因都参与植物抗病途径[50-53],其中CpWRKY33的同源基因AtWRKY50编码的转录因子能与TGA2和TGA5相互作用,协同增强转录激活PR1(拟南芥PR1是SA诱导的系统获得性抗性SAR标记基因)的表达,抵御病原菌侵染[50]。CpWRKY25的同源基因WRKY47过量表达可以提高水稻对半活体生物营养型真菌稻瘟病菌的抗性,但具体作用机制还不清楚[51]。CpWRKY11的同源基因AtWRKY20编码的转录因子可以与编码病程相关蛋白的基因4PR4/AtHEL的启动子结合,介导SAR反应,从而对多种植物病原真菌产生抗性[52]。CpWRKY2的同源基因AtWRKY57在拟南芥受到腐生型真菌灰霉菌侵染时表达明显上调,其与茉莉酸ZIM-DOMAIN1(JAZ1)和JAZ5的启动子结合,激活JAZ1和JAZ5转录(JAZ1和JAZ5编码JA信号通路的2个重要抑制子),负调控拟南芥对灰霉菌的抗性[53]。本研究炭疽菌接种后,CpWRKY2的表达也上调,与拟南芥的结果一致,但其对番木瓜抗性影响的具体机制还需进一步研究。
本研究鉴定出了48个CpWRKY家族基因,其结构和编码的蛋白基序具有组间多样性和组内保守性。CpWRKY家族成员强烈响应番木瓜短胞炭疽菌的诱导,其中CpWRKY22、CpWRKY11、CpWRKY2、CpWRKY33和CpWRKY25是潜在的番木瓜炭疽病抗性基因,对分子育种有潜在的应用价值。
猜你喜欢
湖北农业科学(2022年11期)2022-07-18
中文信息(2017年2期)2017-04-13
江苏农业科学(2016年11期)2017-03-21
智能计算机与应用(2016年4期)2016-09-26
世界热带农业信息(2016年7期)2016-08-23
飞碟探索(2015年9期)2015-11-05
红领巾·探索(2015年9期)2015-09-10
世界热带农业信息(2014年1期)2014-08-09
世界热带农业信息(2014年2期)2014-08-07
世界热带农业信息(2014年4期)2014-08-07
