
清凉峰自然保护区紫茎群落的结构特征分析
2022-05-09 07:09张婷宇张明如万琦李清香蔡益杭赵楠楠
安徽农业科学 2022年8期
张婷宇 张明如 万琦 李清香 蔡益杭 赵楠楠



摘要 [目的]研究浙江清凉峰自然保护区珍稀濒危植物紫茎(Stewartia sinensis)群落的结构特征。[方法]基于取樣代表性的原则,选择可覆盖乔木层紫茎株数占92%地段的样地20 m×30 m,分析群落物种组成、区系成分特征、垂直结构和径级结构。[结果]群落样地共有维管束植物29科38属44种,属和种的数量占优势的科为山茶科(Theaceae)、樟科(Lauraceae)和壳斗科(Fagaceae);种子植物属的一级地理区系成分共含7个类型,其中温带和热带分布分别占总属数的52.78%和41.66%,具有温带至热带的过渡性特征和亚热带性质;群落垂直结构可分为乔木层、灌木层和草本层,其中乔木层的优势种为短柄枹栎(Quercus glandulifera)、黄山松(Pinus taiwanensis)、雷公鹅耳枥(Carpinus viminea)和紫茎(Stewartia sinensis),灌木层的优势种为杜鹃(Rhododendron simsii)、吴茱萸叶五加(Acanthopanax evodiaefolius)和黄丹木姜子(Litsea elongata),草本层的优势种为淡竹叶(Lophatherum gracile); 群落样地乔、灌、草层次的物种多样性表现为乔木层>草本层>灌木层,而盖度表现为乔木层>灌木层>草本层,草本层呈现稀疏的基本特征;清凉峰紫茎种群以乔木层大径级个体占据绝对优势地位,仅有2株种子更新的紫茎幼树,状况严重不良,因此清凉峰紫茎种群结构属于典型的衰退型。[结论]群落乔木层紫茎大径级植株数量远多于紫茎幼树和幼苗的数量,紫茎天然更新状况极差,其种群径级结构类型属于严重的衰退类型。
关键词 清凉峰;紫茎;群落结构;物种多样性;径级结构
中图分类号 Q 948.5;S 792.99 文献标识码 A
文章编号 0517-6611(2022)08-0104-05
doi:10.3969/j.issn.0517-6611.2022.08.028
开放科学(资源服务)标识码(OSID):
Analysis on the Structural Characteristics of the Stewartia sinensis Community in Qingliangfeng Nature Reserve
ZHANG Ting-yu,ZHANG Ming-ru,WAN Qi et al (College of Landscape Architecture, Zhejiang A&F University, Hangzhou, Zhejiang 311300)
Abstract [Objective]To study the structural characteristics of the rare and endangered plant Stewartia sinensis community in Qingliangfeng Nature Reserve of Zhejiang Province. [Method]Based on the principle of sampling representativeness, 20 m×30 m plot with 92% of S. sinensis trees was selected, the species composition, floristic composition, vertical structure and diameter class structure of the community were preliminarily analyzed.[Result]The results showed that there were 44 species of vascular plants belonging to 38 genera and 29 families in the sample plots. Theaceae, Lauraceae and Fagaceae were the dominant families;there were 7 types in the first grade flora of seed plants, of which temperate and tropical distribution account for 52.78% and 41.66% of the total genera respectively;the vertical structure of the community can be divided into tree layer, shrub layer and herb layer. The dominant species in tree layer were Quercus glandulifera, Pinus taiwanensis, Carpinus viminea and S. sinensis. The dominant species in shrub layer were Rhododendron simsii, Acanthopanax evodiaefolius and Litsea elongata, Lophatherum gracile was dominant species in herb layer;the order of species diversity was tree layer>herb layer>shrub layer, and the order of coverage was tree layer>shrub layer>herb layer;the S. sinensis population in Qingliangfeng was dominated by individuals with large diameter class in tree layer, there were only 2 young S. sinensis trees with seed regeneration, and the seed regeneration was seriously poor. Therefore, the population structure of S. sinensis in Qingliangfeng was a typical decline type.[Conclusion] In the tree layer of the S. sinensis community, the number of large diameter plants of S. sinensis was much more than that of young purple stem trees and seedlings. The natural regeneration of S. sinensis was very poor, and its population size structure belongs to a serious decline type.
Key words Qingliangfeng;Stewartia sinensis;Community structure;Species diversity;Diameter class structure
珍稀濒危木本植物的种子更新状况与幼苗、幼树生长发育进程,在很大程度上取决于其所生长分布的群落结构和下层群落非生物生境选择的共同影响,同时也受到种群自身生物学特性和生态适应性的影响。
紫茎(Stewartia sinensis)为山茶科(Teaaceae)紫茎属(Stewartia)中性喜光落叶乔木树种,生长地段土层深厚、疏松肥沃、排水良好;喜凉爽、湿润多雾山地森林环境;观赏价值极高,花瓣白色,树皮光滑呈红褐色或黄褐色。紫茎属于我国亚热带中部地区特有树种,对于研究东亚至北美植物区系及古植物地理有着重要的科学意义 [1-3]。由于天然更新能力弱,加之受生境破坏、人类活动等干扰,紫茎野生种群数量不断减少 [4-5],已被列入国家三级珍稀濒危植物 [1]和浙江省重点保护野生植物 [3]。
紫莖已有的研究成果包括地理区系成分 [6-7]、人工繁殖技术 [8-9]、群落特征 [10-13]、光合生理特性 [14]和药用价值 [15]等。其中,古田山自然保护区紫茎群落样地调查结果表明,组成植物种类丰富,属的分布区类型主要属于温带和热带,以高位芽植物为主,群落物种多样性表现为乔木层>草本层>灌木层,紫茎种群的年龄结构属衰退型 [10-11]。进一步分析紫茎群落树种的种间联结性,认为紫茎与白檀(Symplocos paniculata)、檫木(Sassafras tzumu)和山鸡椒(Litsea cubeba)等9个树种存在较强的竞争排斥关系 [12]。贵州榕江县的调查结果显示,紫茎群落处于演替顶极阶段 [13]。
在野外调查过程中,笔者观察到紫茎多散生分布于常绿落叶阔叶群落和沟谷地带。然而,在紫茎群落下层几乎未见实生幼苗;随海拔下降或升高,当垂直带谱更替为常绿阔叶林或落叶阔叶林,群落内的紫茎植株更为稀少,由此推测常绿落叶混交群落下层的生长光强可能有利于紫茎的种子萌发和幼苗幼树的生长。
因此,群落结构时空异质性变化将极大地影响群落组成植物的更新和定居过程。为了深入分析紫茎群落的结构特征,选择清凉峰自然保护区紫茎群落样地进行调查,分析该群落的物种组成和类型、垂直结构、物种多样性和径级结构 [16]。通过样地调查,旨在揭示紫茎群落的结构特征及紫茎种群天然更新的主要制约因素,为紫茎濒危机制研究和保育提供基础资料,进一步丰富紫茎群落的研究案例。
1 调查区概况与研究方法
1.1 调查样地自然概况 调查地段位于浙江省杭州市临安区境内的清凉峰国家级自然保护区,地理位置为118°53′E,30°11′N,海拔1 000 m,属于亚热带季风气候,年均气温7.8~15.3 ℃,1月均温3.4 ℃,7月均温28.1 ℃;全年降水量1 500~1 900 mm,雨热同期,无霜期234 d [17];森林群落类型属于常绿落叶阔叶混交群落;土壤类型主要为黄壤,坡位为中部,坡度43°,坡向192°S。
1.2 调查方法 基于“取样代表性”的原则 [18],即群落样地包含紫茎种群绝大多数植株,确定样地面积为600 m 2。用测绳将20 m×30 m的样地划分为6个10 m×10 m的乔木样方,在每个乔木样方的右下角分别划出1个4 m×4 m的灌木层小样方 (包括幼树、幼苗)和1个2 m×2 m的草本层小样方。乔木逐株调查测定,记录包括乔木层、灌木层和草本层的植物,并记载群落外貌、垂直结构层次。
1.3 数据处理 将调查群落垂直层次分为乔木层、灌木层、草本层(含层间植物)3个层次,重要值(importance value, IV)以相对多度、相对频度、相对优势度或相对盖度的累加值进行计算。其中,乔木层的重要值(%)=[相对多度(%)+相对频度(%)+相对优势度(%)]/3;灌木层、草本层的重要值(%)=[相对多度(%)+相对频度(%)+相对盖度(%)]/3。
物种丰富度指数(S)为样地内所有物种的种类数目;Shannon-Wiener指数(H′)=- si=1(Pi×lnPi);Simpson指数(优势度指数)(P)=1-∑ si=1Pi 2;Pielou指数(均匀度指数)(E)=H′/lnS;式中,Pi为种i的重要值(IV)。
2 结果与分析
2.1 群落科属构成 根据野外调查统计结果,紫茎群落共有维管束植物29科38属44种。其中,被子植物有27科36属42种,裸子植物有2科2属2种,未分布蕨类植物。乔木层植物有19科26属32种;灌木层植物有8科12属13种;草本层植物有12科13属13种;层间植物有2科2属2种。样地内属和种数量占优势的科为山茶科(Theaceae,4属6种)、樟科( Lauraceae,3属5种)、壳斗科(Fagaceae,3属4种);仅含1属1种的科有22个,占总科数的75.86%。
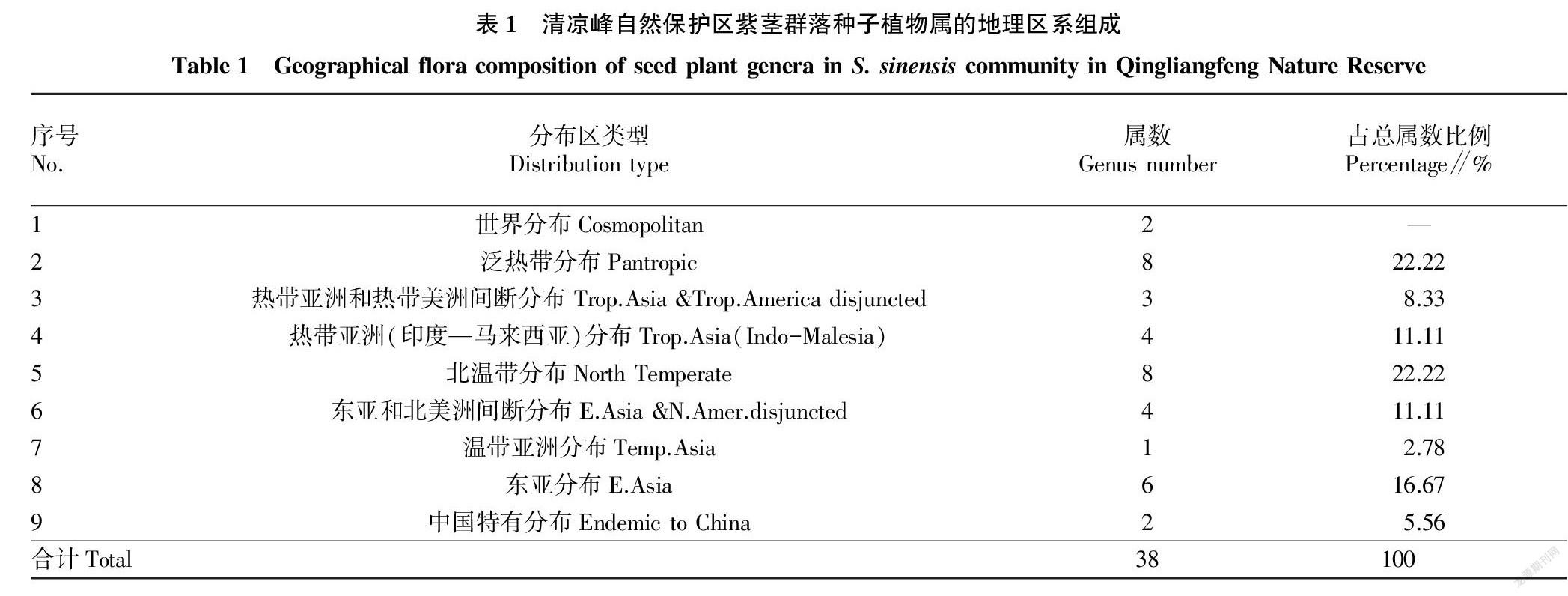
2.2 群落区系组成 植物区系组成在一定程度上直观地反映植被特征及地理环境变化 [19]。根据中国种子植物属分布区类型的划分标准 [20],对组成该群落的种子植物区系地理成分进行统计分析,结果见表1。由表1可知,组成该群落的38个属可划分为9个分布区类型,有2属为世界分布;在植物地理区系组成成分属热带分布型有15属,占总属数41.66%;温带分布型有19属,占总属数52.78%;中国特有分布有2属,占总属数5.56%,分别为杉木属(Cunninghamia)和香果树属(Emmenopterys)。其中,泛热带分布型8属,北温带分布型8属,东亚分布型6属,表明该群落具有一定热带向温带的过渡性特征和亚热带性质 [21]。样地未分布旧世界热带分布、热带亚洲至热带大洋洲分布、热带亚洲至热带非洲、旧世界温带分布、地中海区和西亚至中亚分布、中亚分布类型。
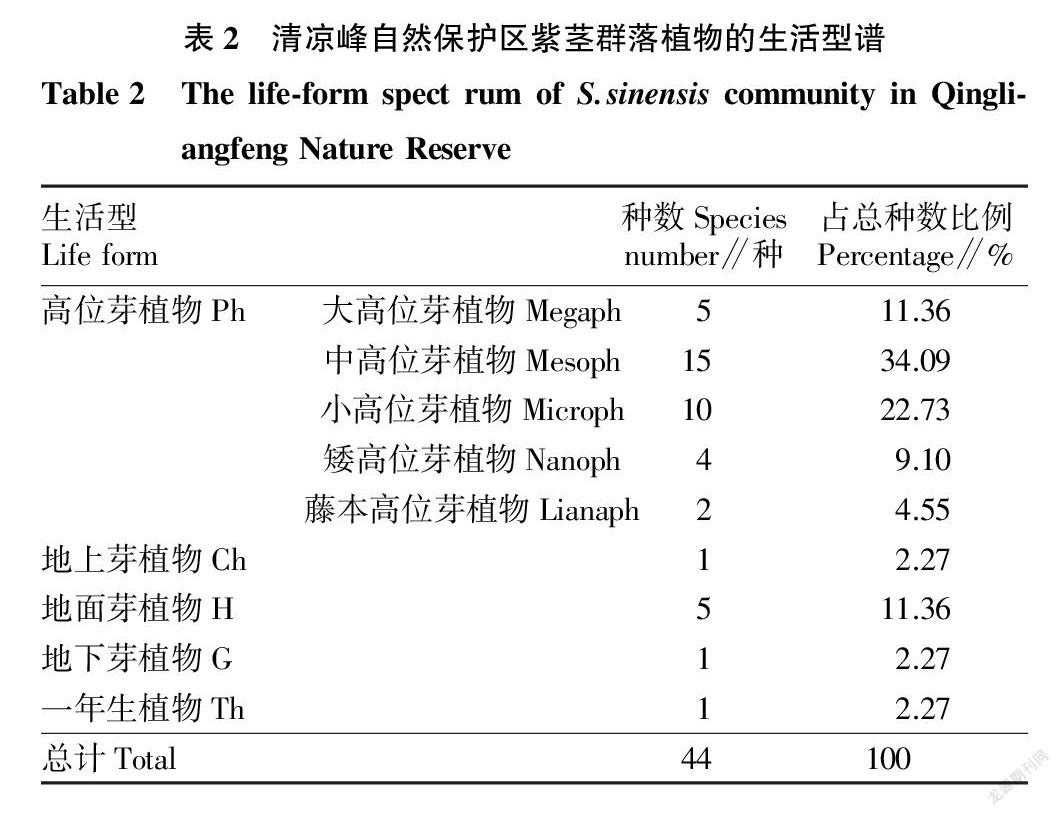
2.3 群落生活型比例 根据丹麦植物学家Raunkiaer的生活型分类系统,绘制清凉峰紫茎样地维管束植物生活型谱 [22]。结果表明(表2),该样地高位芽植物种类最多,共36种,占总种数的81.83%,其中种类数量表现为中高位芽植物>小高位芽植物>大高位芽植物>矮高位芽植物>藤本高位芽植物;其次是地面芽植物,占11.36%;地上芽、地下芽和一年生植物均占2.27%。
2.4 群落植物叶片性质 参考丹麦植物学家Raunkiaer提出的叶级分类方式将样地内出现的植物叶级划分为4个等级:微型叶(0.26~2.25 cm 2)、小型叶(2.26~20.25 cm 2)、中型叶(20.26~182.25 cm 2)和大型叶(182.26~1 640.25 cm 2),绘制清凉峰紫茎所在群落样地维管束植物叶相谱图 [22]。结果表明,就叶的大小而言,该群落的植物以小型叶为主,占总种数的54.55%,中型叶占31.82%,微型叶占9.09%,大型叶占4.54%;就叶的类型而言,单叶占77.27%,复叶占22.73%;就叶的质地而言,叶为革质占43.18%,厚革質占2.27%,纸质占47.73%,草质占6.82%;就叶缘而言,非全缘叶略多于全缘叶(表3)。此外,样地中阔叶树种类占总种数的比例高达72.73%,针叶树占4.55%;在阔叶树中常绿树种占43.33%,落叶树种占56.67%。
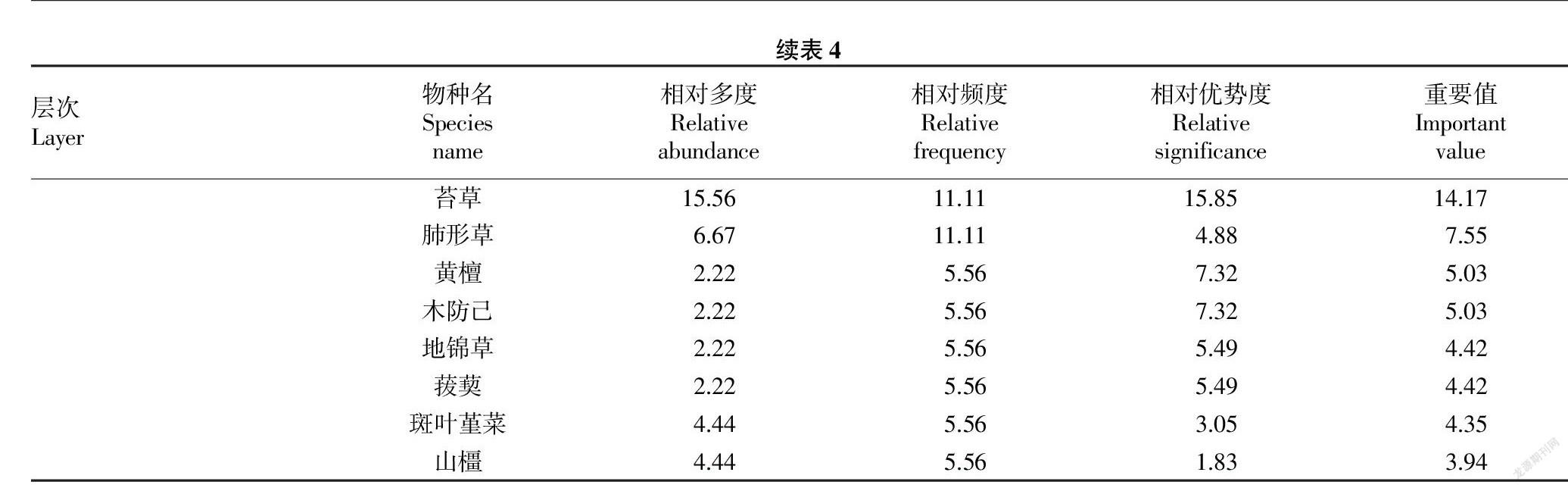
2.5 群落垂直层次结构 该群落垂直结构可分为乔木层、灌木层和草本层。乔木层有19科26属32种植物,可分为2个亚层,第一亚层高8.60~18.50 m,胸径15.50~35.90 cm,优势种为短柄枹栎(Quercus glandulifera)、黄山松(Pinus taiwanensis)、紫茎等;第二亚层高3.50~8.50 m,胸径2.30~15.40 cm,优势种为雷公鹅耳枥(Carpinus viminea)、小叶青冈(Cyclobalanopsis myrsinifolia)、细叶青冈(Cyclobalanopsis myrsinaefolia)、格药柃(Eurya muricata)等。根据物种重要值可知(表4),有6种乔木层树种的重要值均大于7.00%。灌木层有8
科12属13种植物,株高1.50~3.50 m,其中重要值大于7.00%的灌木树种为杜鹃(Rhododendron simsii)、吴茱萸叶五加(Acanthopanax evodiaefolius)、黄丹木姜子(Litsea elongata)、紫茎、宜昌荚蒾(Viburnum erosum)5种。草本层有12科13属13种植物,其中包括2科2属2种藤本植物,为木防己(Cocculus orbiculatus)、菝葜(Smilax china);重要值大于7.00%的草本层物种为淡竹叶(Lophatherum gracile)、吴茱萸叶五加、苔草(Carex spp.)、肺形草(Tripterospermum chinense)。
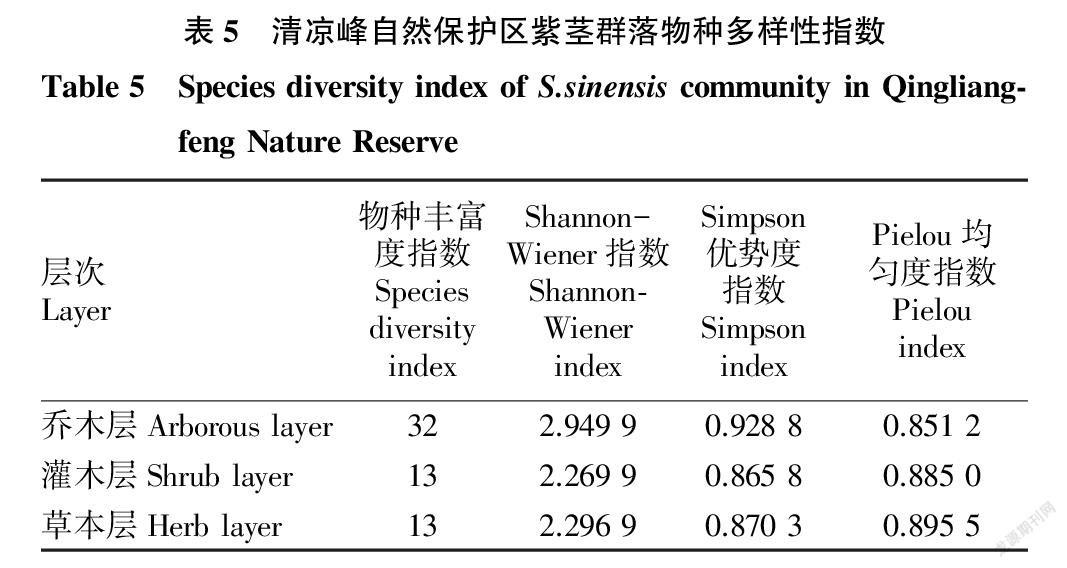
2.6 物种多样性指数比较 物种多样性(community diversity)属于反映植物群落结构和功能的重要指标之一。为了减小误差,以物种重要值(IV)作为测度Shannon-Wiener指数、Simpson指数和Pielou指数的依据。结果表明(表5),该群落不同层次的物种多样性表现为乔木层>草本层>灌木层。其中,乔木层的Shannon-Wiener指数和Simpson优势度指数高于灌木层和草本层,而Pielou指数稍低于灌木层和草本层;灌木层和草本层的各项指数相差较小。
2.7 群落径级结构 径级结构是植物群落稳定性和生长发育状况的重要表征指标 [23]。统计样地内所有乔木DBH每递增2 cm所含个体数量占总数的百分比,再以同样的方法统计紫茎种群。结果表明(图1),当DBH≤8 cm时,随着DBH的增大,样地内乔木个体数量总体呈递增趋势;当DBH>8 cm时,随着DBH的增大,样地内乔木个体数量呈递减趋势。样地内所有乔木径级结构呈类倒“J”型(偏正常型),但个体数量集中在4 cm≤DBH≤10 cm;紫茎种群呈峰型(中径级个体储备型),个体主要集中在14 cm≤DBH≤22 cm,且在6 cm≤DBH≤12 cm存在个体缺失现象,由此表明,紫茎幼树出现典型的缺失现象,且幼苗株数明显偏少。
3 讨论
群落稀有种的分布取决于物种种群特征、植物区系交汇以及人为或自然干扰 [23],而濒危种的出现与群落稀有种出现的原因部分重合。紫茎主要分布于海拔600~1 900 m的阳坡、河谷和沟谷地带,总体数量偏少,植株分布分散,野生种群处于濒危状态。调查群落的紫茎在乔木层处于稳定优势地位,但由于紫茎种群天然更新受阻,年龄结构不合理,衰退趋势明显,样地仅有2株种子更新的紫茎幼树,其余均为同一株紫茎母树的根蘖植株;而在样地外道路侧方土壤贫瘠裸露的地段,沿道路长约110 m,分布生长有13株紫茎实生幼苗和幼树,长势良好,株高为52.52~128.42 cm,地径6.22~12.64 mm。在紫茎生长分布的群落下层,由于乔木层郁闭度较高、透光不足,加之灌木层和草本层以及枯枝落叶层的架空阻碍作用,幼苗建成机会较小。根蘖苗与紫茎母树争夺空间和环境资源的能力较弱,特别是群落下层生长光强明显不足,结果根蘖繁殖难以成为紫茎的主要繁殖方式,实生幼苗缺失又极大地影响了紫茎种群结构的稳定性 [24]。由此,可以认为紫茎种子萌发过程需要接触土壤,幼苗幼树乃至种子发芽需要一定的侧方光照,或者林窗及疏林地段的光照条件。因此,凋落物层属于紫茎种子发芽的重要生境,草本层和灌木层属于紫茎幼苗幼树生长发育的重要生境,甚至对于整个群落的更新具有重要意义 [25]。
进一步分析紫茎径级分布可知,紫茎种群呈中径级个体储备型,径经在4 cm≤DBH≤12 cm存在个体缺失现象,而调查群落内其他乔木树种的个体集中于紫茎种群个体缺失的径级区间内,导致在幼树阶段其他乔木树种对紫茎施加不对称的种间竞争排斥。随着径级的增大,紫茎个体数量大量减少,说明紫茎的天然更新不良与群落的种间竞争排斥有一定的关系,群落上层结构影响紫茎小径级个体的分布与生长,可能属于群落的自疏与他疏作用 [25]。
群落的组成特征可以通过群落物种库的大小、非生物环境、种间相互作用等加以判断 [26],则濒危植物的种群更新过程主要受森林群落乔、灌、草层次遮蔽、非生物环境的过滤和种间竞争排斥的影响。木本植物从种子发芽到幼苗建成的过程中受到群落内动态光照、凋落物、水分等因素的综合影响,紫茎生长过程中存在常绿落叶阔叶混交林和针阔叶混交林累积的凋落物遮盖地表的情况,阻挡种子接触土壤,影响种子发芽和幼苗建成,导致实生幼苗缺失。此外,阔叶群落下层凋落物在土壤中分解释放化感物质,极有可能是紫茎天然更新的潜在抑制因素 [27]。
群落内植物间竞争将导致生态位压缩,物种的独立性和相互依存性随时间和空间的改变而发生变化 [28]。如果可以建立群落动态模拟模型,量化物种间的相互作用和共存过程 [29-30],深入分析群落内紫茎种子扩散、发芽到幼苗建成等重要阶段的性状表现,对进一步研究紫茎濒危机制具有重要意义。当然还需要加强野生植物资源的调查和就地保护,在群落竞争评估的基础上对影响紫茎生长的竞争树种进行科学合理的修枝或择伐,并采取人工扩繁栽培试验促进紫茎就地保育进程,所采取的措施对于珍稀濒危植物紫茎种群的持续繁衍具有重要作用。
4 结论
通过调查分析清凉峰自然保护区珍稀濒危植物紫茎群落的结构特征,得如下结论:
(1)该群落样地内共有维管束植物29科38属44种,被子植物27科36属42种,裸子植物2科2属2种; 27科被子植物只含1属1种的有22科,占总科数的75.86%,说明该群落物种的科、属组成相对分散。
(2)群落植物地理区系组成结果属于温带成分(52.78%)的属略多于热带成分(41.66%),表明该群落呈现一定热带向温带的过渡性特征和亚热带性质。
(3)群落类型属于常绿落叶阔叶混交林,且混生一定数量的黄山松,高位芽植物占总种数的81.83%;群落以单叶、革质、中小型叶为主,非全缘叶略多于全缘叶。
(4)群落垂直结构可分为乔木层、灌木层和草本层,乔木层优势种为短柄枹栎、雷公鹅耳枥、紫茎和黄山松,灌木层优势种为杜鹃、吴茱萸叶五加和黄丹木姜子,草本层优势种为淡竹叶。
(5)群落各层盖度大小表现为乔木层>灌木层>草本层;灌木层植物盖度较草本层大,但种类少,物种多样性大小表现为乔木层>草本层>灌木层。
(6)群落径级结构显示中、小径级(2 cm≤DBH≤22 cm)的乔木占91.38%,天然更新良好,属增长型;样地紫茎种群的小径级植株和幼苗幼树极少甚至缺失,表明天然更新严重受阻,属衰退型。
参考文献
[1]
傅立国.中国植物红皮书——稀有濒危植物:第1册[M].北京:科学出版社,1992.
[2] SPONGBERG S A.A review of deciduous-leaved species of Stewartia (Theaceae)[J].J Arnold Arbor,1974,55(2):182-214.
[3] 张若惠.浙江珍稀濒危植物[M].杭州:浙江科学技术出版社,1994:1-17.
[4] 刘兴剑,孙起梦.美木紫茎入园林 观花赏干两相宜[J].植物杂志,1999(5):3-5.
[5] 蔡晟,刘学全,张家来,等.鄂西三峡库区大老岭珍稀树木群落特征研究[J].应用生态学报,2000,11(2):165-168.
[6] 李捷.山茶科紫茎属和舟柄茶属的系统学研究[J].植物分类学报,1996,34(1):48-67.
[7] 李密密.山茶科及紫茎属的系统发育和生物地理研究[D].杭州:浙江大学,2011.
[8] 左艺芳.紫茎繁殖技术体系的研究[D].合肥:安徽农业大学,2007.
[9] 左艺芳,黄成林.观赏树种紫茎扦插繁殖的研究[J].安徽农学通报,2007,13(7):94-95,203.
[10] 胡正华,于明坚,余建平,等.浙江古田山自然保护区紫茎林特征研究[J].中国计量学院学报,2002,13(3):199-204.
[11] 钱海源,唐战胜,余建平,等.浙江古田山珍稀植物紫茎群落的组成与结构特征[J].贵州农业科学,2019,47(1):152-157.
[12] 董瑞瑞,唐战胜,陈建华,等.珍稀濒危植物紫茎群落树种的种间联结性[J].安徽农业科学,2018,46(5):127-129,153.
[13] 周庆,李晓芳,骆畅,等.黔东南一个紫茎群落的研究[J].贵州科学,2018,36(5):46-50.
[14] 吴泽群.珍稀濒危植物紫茎Stewartia sinensis的种子生理特性和光合生理特性研究[D].杭州:浙江農林大学,2019.
[15] WAN J,ZANG Y,XIAO D A,et al.Stewartiacids A-N,C-23 carboxylated triterpenoids from Chinese Stewartia and their inhibitory effects against ATP-citrate lyase and NF-κB[J]. RSC advances,2020,10(6):3343-3356.
[16] 方精云,王襄平,沈泽昊,等.植物群落清查的主要内容、方法和技术规范[J].生物多样性,2009,17(6):533-548.
[17] 宋朝枢.浙江清凉峰自然保护区科学考察集[M].北京:中国林业出版社,1997.
[18] 马克明,张洁瑜,郭旭东,等.农业景观中山体的植物多样性分布:地形和土地利用的综合影响[J].植物生态学报,2002,26(5):575-588.
[19] 王杰,王計平,张华新,等.滦河河口地区植物区系分析及多样性研究[J].山西农业大学学报(自然科学版),2020,40(4):111-120.
[20] 吴征镒.中国种子植物属的分布区类型[J].云南植物研究,1991(增刊IV):1-139.
[21] 陈金磊,方晰,辜翔,等.中亚热带2种森林群落组成、结构及区系特征[J].林业科学,2019,55(2):159-172.
[22] RAUNKIAER C.The life forms of plants and statistical plant geography[M].New York:Oxford University Press,1932.
[23] 叶万辉,曹洪麟,黄忠良,等.鼎湖山南亚热带常绿阔叶林20公顷样地群落特征研究[J].植物生态学报,2008,32(2):274-286.
[24] 刘海洋,金晓玲,薛会雯,等.壶瓶山自然保护区珍稀濒危植物珙桐群落的研究[J].中南林业科技大学学报,2011,31(4):31-36,41.
[25] 徐敏,骆争荣,于明坚,等.百山祖北坡中山常绿阔叶林的物种组成和群落结构[J].浙江大学学报(农业与生命科学版),2007,33(4):450-457.
[26] 练琚愉,陈灿,黄忠良,等.鼎湖山南亚热带常绿阔叶林不同成熟度群落特征的比较[J].生物多样性,2015,23(2):174-182.
[27] FACELLI J M,PICKETT S T A.Plant litter: Its dynamics and effects on plant community structure[J]. Botanical review, 1991, 57(1):1-32.
[28] LORTIE C J,BROOKER R W,CHOLER P,et al. Rethinking plant community theory[J]. Oikos, 2004, 107(2):433-438.
[29] WIEGAND T,URIARTE M,KRAFT N J B,et al. Spatially explicit metrics of species diversity,functional diversity, and phylogenetic diversity:Insights into plant community assembly processes[J].Annual review of ecology evolution and systematics, 2016, 48(1):329-351.
[30] CAPIT N J A,CUENDA S,ALONSO D.Competitive dominance in plant communities:Modeling approaches and theoretical predictions[J/OL].Journal of theoretical biology,2020,502[2021-03-14].https://doi.org/10.1016/j.jtbi.2020.110349.
猜你喜欢
浙江农林大学学报(2022年6期)2022-12-07
热带农业工程(2022年3期)2022-08-16
云南化工(2021年7期)2021-12-21
热带农业科学(2016年11期)2017-01-21
热带农业科学(2016年11期)2017-01-21
绿色科技(2016年21期)2016-12-27
现代农业科技(2016年20期)2016-12-20
科技创新导报(2016年21期)2016-12-17
现代经济信息(2016年13期)2016-06-17
湖南农业(2016年3期)2016-06-05
