
青花菜、番茄与辣椒根肿菌侵染过程研究
2022-04-26 10:55:36刘玉梅韩风庆李占省
中国蔬菜 2022年4期
谢 琪 刘玉梅 韩风庆 庄 木 李占省
(中国农业科学院蔬菜花卉研究所,农业农村部园艺作物生物学与种质创制重点实验室,北京 100081)
根肿病是一种土传病害,对世界范围内的十字花科作物危害严重,该病可由芸薹根肿菌()侵染植株根系引发,根肿菌在土壤中可存活数年,传播方式呈现多样化。近些年,我国青花菜(L.var.)栽培面积逐年增加(李占省 等,2021),根肿病也开始在部分地区发生且呈扩大趋势,目前已在浙江、云南、湖北、河南、重庆等地发生。根肿病会造成青花菜商品性降低,甚至难以形成正常花球,影响产量和质量,严重时造成绝收。
十字花科根肿病的防治方法有物理防治(撒石灰、石灰氮等)和化学防治(氟啶胺)等,但最根本的方法是选育抗性品种(Webster &Dixon,1991;Wallenhammar et al.,2008)。目前,根肿菌在土壤中侵染十字花科作物的生活史已很清楚(杨晓琴,2015)。十字花科根肿病鉴定方法主要分为两类,一是建立在病圃基础上的鉴定技术,二是离体接种鉴定。两种方法鉴定周期均较长,管理风险大,耗费大量的人力与物力,故探索和建立十字花科根肿病快速鉴定方法将极大地提高鉴定效率(Deora et al.,2012)。
根肿菌生理小种的鉴定方法主要有两种:Williams 鉴定系统和欧洲根肿病ECD 鉴定系统(Williams,1966;Buczackl et al.,1975)。Williams 鉴定系统具有寄主少、简单易操作等优点,在国际上被广泛使用。然而,Williams 鉴定系统只能鉴别出13 个生理小种,且当多个生理小种混杂时,采用Williams 鉴定系统并不能够准确地将优势小种鉴别出来,所以该方法存在一定的局限性(Feng et al.,2016;白宇鹏,2019)。ECD 鉴定系统具有较好的实用性,准确度较高,且能够更加细致地划分生理小种,但ECD 鉴定系统寄主较多,存在难收集、工作量较大和操作程序较为复杂的问题(丁云花 等,2013;朱翠娟,2016)。原玉香等(2017)研究发现,在对同一地区的2 份菌株进行鉴定时,采用Williams 鉴定系统和ECD 鉴定系统鉴定的结果不一致,表明两种鉴定系统对同一地区生理小种的鉴定结果存在差异。近些年,水培接种法作为一种快速鉴定根肿病抗性的技术逐渐显现其优越性,具有操作简单、方便快捷等优点,通过离体接种和染色观察(杜艳 等,2016),可以从细胞学水平上快速鉴定十字花科作物根肿病抗性,应用前景广阔。
相比十字花科作物,根肿病不会对茄果类作物造成危害,探寻及利用该抗性机理选育抗根肿病的十字花科作物新品种具有极其重要的意义和科学价值。因此,揭示并阐明十字花科和茄果类作物在根肿菌侵染过程及其防御能力上的差异,将为创制十字花科抗根肿病种质资源提供新思路。
1 材料与方法
1.1 试验材料
本试验于2019 年7—9 月在中国农业科学院蔬菜花卉研究所进行,种子均为商品种,青花菜品种为中青10 号和中青16 号,番茄()品种为中杂302,辣椒(L.)品种为中椒115,均由本所自主育成。
1.2 试验方法
1.2.1 种子预处理 2019 年7 月将青花菜、番茄和辣椒种子撒播在浸湿的滤纸上,将滤纸放入培养皿中催芽,6 d 后将幼苗移到15 mL 的离心管中,每管加入14 mL 蒸馏水,只放入1 株幼苗用泡沫筛固定,离心管用锡箔纸包裹进行遮光处理,根肿菌侵染过程在水培溶液中进行(图1)。

图1 青花菜、番茄与辣椒水培试验
1.2.2 根肿菌分子辅助鉴定 供试根肿菌菌株来自河南新野青花菜、大白菜肿块,由河南省农业科学院园艺研究所魏小春老师提供并已鉴定为生理小种4 号,将根肿病肿块洗净风干后超低温(-80 ℃)保存备用。采用CTAB 法提取肿块DNA(李占省,2012)。
根据已报道的根肿菌生理小种特异引物(表1)进行PCR 扩增和鉴定(Zheng et al.,2018)。PCR 反应体系:5.0 μL 的2×Plus Master Mix,2.0 μL 的ddHO,2.0 μL DNA,正反引物各0.5 μL,总体积10.0 μL。扩增程序:94 ℃预变性5 min;94 ℃变性30 s,55 ℃退火30 s,72 ℃延伸45 s,35 个循环;72 ℃后延伸5 min,10 ℃保存。采用1%琼脂糖凝胶电泳鉴定,每孔点入5 μL PCR 产物,在150 V 恒压下电泳25 min,后置于自动凝胶成像系统中成像观察。

表1 根肿菌生理小种特异引物信息
1.2.3 根肿菌菌液制备 将-80 ℃冰箱中的根肿病肿块在室温下解冻,称取10.0 g,用刀片将根肿病肿块切成小片后放入研磨机,加入1.0 L 蒸馏水机械研碎成浆液,时间为3 min。将磨好的浆液用8 层尼龙网布过滤,以500 r·min的转速离心5 min,取上清液,最后,用血球计数板和光学显微镜将孢子浓度稀释至4 × 10个·mL,4 ℃保存备用(图2)。

图2 根肿菌菌液制备流程
1.2.4 根肿菌侵染 移至离心管的中青16 号、中青10 号、中椒115 和中杂302 幼苗水培3 d 后,在离心管中加入2 mL 孢子浓度为4 × 10个·mL的根肿菌菌液,侵染处理过程参考张小丽(2014)的方法并进行调整。将接种后的水培苗放入人工气候培养箱中培养。培养条件:16 h 光照,25 ℃;8 h黑暗,20 ℃;相对湿度75%。
1.2.5 镜检观察 在接种后7、12、14 d,利用显微镜对供试对象根细胞进行观察,确定初级、次级侵染时期根肿菌侵染过程。首先,用蒸馏水清洗试验材料根上的附着孢子,然后将根切成1~2 cm 长的小段,用FAA 固定24 h,同时用荧光桃红染液染色15 min,染好的根部组织置于载玻片上用清水冲洗掉多余染液,最后进行镜检观察,分别统计侵染和未侵染的根毛数,计算根毛侵染率。每时间点选取3 个单株分别观察和拍照。
根毛侵染率=(被侵染的根毛数/观察的总根毛数)× 100%
2 结果与分析
2.1 生理小种的分子辅助鉴定
根据已报道的根肿菌生理小种特异引物对河南新野菌株进行PCR 扩增,结果表明,6 对特异引物Novel342-2、Novel407-2、PBRA_007750-2、PBRA_008439-1、PBRA_009348-1 和Actin1 均能扩增出目标条带(图3)。通过Novel342-2、PBRA_009348-1 和Novel407-2 PCR 扩增结果可以确定4 号、7 号和9 号生理小种的存在情况,若特异引物Novel342-2 能扩增出特异产物,则可以鉴定存在根肿菌4 号生理小种;若特异引物Novel342-2 或Novel407-2 不能扩增出特异产物,则可以鉴定存在根肿菌7 号生理小种;若特异引物PBRA_009348-1 能扩增出特异产物,且Novel342-2 或Novel407-2 不能扩增出目标产物,则可以鉴定存在根肿菌9 号生理小种。本试验中,特异引 物Novel342-2、PBRA_009348-1和Novel407-2 均能扩增出特异片段,故可以确定该菌株存在4 号生理小种,不存在7 号和9号生理小种。通过特异引物PBRA_007750-2、PBRA_009348-1 和PBRA_008439-1 的PCR 扩增结果可以判断11 号生理小种的存在情况,若特异引物PBRA_009348-1 未扩增出特异产物而PBRA_007750-2 或PBRA_008439-1 可以扩增出特异产物,则可以推断存在根肿菌11 号生理小种,但本试验中,特异引物PBRA_009348-1 能扩增出特异产物,可推断该供试菌株中不存在11 号生理小种。综上所述,推断该根肿菌只存在4 号生理小种(表2),这与河南省农业科学院园艺研究所原玉香等(2017)的报道结果一致。

图3 根肿菌生理小种PCR 扩增产物

表2 根肿菌生理小种标记鉴定条件及结果
2.2 青花菜、番茄和辣椒根肿菌侵染过程观察比较
水培青花菜根肿菌侵染过程显示,青花菜根部用荧光桃红染液染色后,被根肿菌侵染的根毛呈红色,未被侵染的根毛为无色。图4-a 显示,在接种后7 d,青花菜根毛细胞内有大量分生孢子侵入,表明根肿菌已侵染根毛,此时中青16 号的根毛侵染率为46.62%(图5)。在青花菜根肿菌侵染过程中,中青16 号比中青10 号生长势强,接种后,中青16 号能维持较长一段时间的正常生长,而中青10 号在接种后死亡速度较快,故未能进行观察。图4-a Ⅳ中可以观察到有部分孢子从根毛已进入到根部皮层细胞内,图4-a Ⅵ表明根肿菌已明显侵染根部皮层细胞,根部皮层细胞内有大量的孢子囊形成。随着侵染时间的延长,接种后12 d 比7 d 时的根毛侵染率明显提高25.38%。到接种后14 d 时,根毛侵染率上升减缓,比接种后12 d 时提高了11.40%,根毛侵染率达到83.40%。由此可见,在接种后7~14 d 根肿菌能够快速侵染青花菜,且在不同基因型间存在侵染速率上的差异。本试验中,中青16 号相比中青10 号在接种后能够延长生存时间,滞缓根肿菌侵染速率,但在接种后14 d 时,中青16 号根部皮层细胞内仍形成大量的孢子囊。
番茄品种中杂302 水培接种根肿菌侵染过程显示(图4-b),在接种后7 d 时根肿菌开始侵染根毛,此时根毛侵染率较高,为32.80%(图5),但比青花菜中青16 号的根毛侵染率低。接种后12 d 时仍有部分番茄根毛被根肿菌侵染,相比接种后7 d 的根毛侵染率提高不到10%,部分根肿菌孢子已进入根部皮层细胞,但明显低于中青16 号的侵染率。接种后14 d 时番茄根毛发生明显扭曲,根部皮层细胞可明显观察到侵入的根肿菌分生孢子,此时根毛侵染率为48.21%。从根肿菌的侵染过程来看,番茄品种中杂302 与青花菜品种中青16 号的侵染途径近似:根肿菌先侵染根毛区,再逐渐侵染根部皮层,但在接种后7~14 d 中杂302 的根毛侵染率明显低于同时期的中青16 号;相比青花菜,番茄皮层细胞内仅能形成少量的分生孢子,不能形成孢子囊。

图4 根肿菌侵染中青16 号、中杂302 及中椒115 的根部染色观察
从辣椒品种中椒115 水培接种根肿菌侵染结果可以看出(图4-c),中椒115 根肿菌孢子进入根毛量最少,接种后7 d 时根毛细胞极少被根肿菌孢子侵染,根毛侵染率只有10.56%(图5)。接种后12 d 时根肿菌已侵染根部皮层细胞,但只有极少数分生孢子,没有形成孢子囊,根毛侵染率较低,为12.6%。接种后14 d 时根毛侵染率只上升不到2%。相比中青16 号和中杂302,中椒115 根毛侵染率最低,程度最轻。
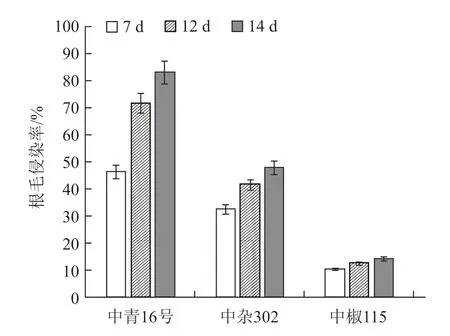
图5 不同品种在不同时期的根毛侵染率
3 结论与讨论
近年来,十字花科作物受根肿病影响和危害越来越大,备受国内外关注。青花菜在国内外广泛种植,2019 年我国种植面积约8.67 万hm(李占省 等,2019)。目前,根肿病已严重危害青花菜主产区的栽培,如浙江台州、云南通海、河南新野等地,危害面积逐年增加。
为了更好地挖掘和创制青花菜抗根肿病资源,本试验以抗根肿病的茄果类蔬菜作物番茄和辣椒为对照,以感病青花菜作为目标研究对象,通过水培法观察根肿菌侵染茄果类和十字花科类作物根部细胞的过程,来揭示对照组和感病组在根肿菌侵染过程中的差异变化,为十字花科选育抗根肿病资源提供一种新思路。本试验为了准确鉴定根肿菌生理小种的种类,结合国内外报道,特选用根肿菌生理小种特异分子标记进行鉴定,与原玉香等(2017)报道的河南新野菌株为4 号生理小种的研究结果一致,表明分子鉴定可作为一种辅助手段实现根肿菌生理小种的快速鉴定。
不同青花菜基因型间以及不同科属作物间根肿菌的侵染过程均存在显著差异,但相同的是番茄、辣椒与青花菜一样,在水培条件下都能够被根肿菌侵染,且其侵染过程与十字花科作物青花菜相似,都是从进入根毛开始,再侵染根部皮层细胞,形成分生孢子或孢子囊,但在根肿菌侵染率方面,三者存在明显差异:青花菜上根肿菌的根毛侵染率明显高于番茄和辣椒。从观察结果来看,虽然后期中青16 号、中杂302 和中椒115 的根毛侵染率均有所增加,但番茄和辣椒明显低于青花菜,与接种后12 d相比,接种后14 d时番茄根毛侵染率提高了6.41%,辣椒根毛侵染率提高了1.80%,而青花菜根毛侵染率提高了11.40%;从进入根毛的根肿菌孢子数量来看,中青16 号也明显多于中杂302 和中椒115。综上所述,青花菜与番茄、辣椒在根肿菌侵染率方面存在明显差异,青花菜根肿菌侵染速率和侵染率最高,其次是番茄,辣椒最低,且番茄、辣椒根部皮层细胞不能形成孢子囊。
本试验中,番茄和辣椒根际分泌物可能发挥了根肿菌防御作用,但在水中根际分泌物可能有别于土壤(Gossen et al.,2019),水中番茄和辣椒的根际分泌物会被稀释,因此根肿菌能够侵染根毛和皮层。土壤栽培环境下根肿病并不会对番茄和辣椒造成危害,从而验证了茄果类蔬菜根际分泌物的重要性,暗示了可以通过在土壤中增施茄果类蔬菜根际分泌物的方式来预防和降低十字花科蔬菜根肿病的危害,这将是一种更加绿色、环保和高效的方法(张婷玉 等,2014;韩旭 等,2015;杨丹丹 等,2018;Fu et al.,2019)。此外,高附加值的十字花科作物或许可以通过嫁接茄果类作物优良砧木来预防根肿病的发生。本试验结果为以上两种方法提供了一定的科学依据和技术支撑,为选育十字花科高抗和免疫根肿病材料提供了新思路。目前,国内外报道的十字花科蔬菜抗根肿病基因主要来自芜菁、萝卜和大白菜(Suwabe et al.,2003,2006;Zhang et al.,2015),其中芜菁报道最多,发现了11 个主效基因或QTL(Piao et al.,2004),茄果类蔬菜尚未见抗根肿病基因的相关报道,其免疫机理有待深入研究与利用。
猜你喜欢
植物研究(2023年3期)2023-05-21 07:51:20
杭州师范大学学报(自然科学版)(2022年2期)2022-04-06 07:48:44
南京农业大学学报(2021年3期)2021-05-24 07:55:20
武夷学院学报(2019年6期)2019-09-06 03:41:48
新农民(2019年29期)2019-02-21 06:35:38
长江蔬菜(2018年2期)2018-04-08 03:02:59
西北植物学报(2018年1期)2018-04-08 01:53:45
农业工程学报(2018年3期)2018-03-01 09:35:17
长江蔬菜(2017年10期)2017-03-25 22:34:12
西南农业学报(2016年6期)2016-04-16 05:12:51
