
生态浮床对千岛湖水体氮磷净化效果研究
2022-04-25 05:25朱广伟王裕成韩轶才王子聪朱梦圆
环境科学研究 2022年4期
唐 伟,许 海,詹 旭,朱广伟,王裕成,韩轶才,王子聪,朱梦圆
1. 江南大学环境与土木工程学院,江苏 无锡 214122
2. 中国科学院南京地理与湖泊研究所,湖泊与环境国家重点试验室,江苏 南京 210008
3. 杭州市生态环境局淳安分局,浙江 杭州 311700
4. 杭州市生态环境科学研究院,浙江 杭州 310005
湖泊是地表极其宝贵的淡水资源,由于城市化进程加速、人口膨胀、工业快速发展等因素,氮磷富集造成的湖泊富营养化问题成为全球淡水生态系统面临的严峻挑战之一. 湖泊富营养化诱发蓝藻水华,产生藻毒素等有害物质,对人体和动物产生毒害作用,降低人类生活质量,阻碍经济持续发展.因此,迫切需要对湖泊富营养化进行生态修复和有效控制.
生态浮床技术是一种常用的人工强化脱氮除磷工程技术,是控制水体富营养化的一种重要手段,其原理是将水生植物或部分陆生植物固定在浮床上,浮床植物不仅吸收水体中的营养盐,植物根系还为反硝化等脱氮菌群提供天然繁殖场所,提高水体除氮能力,从而达到去除水体氮、磷等营养盐的目标. 生态浮床技术能充分利用水面而无需占用土地,可应用于各种水深条件的水体,且造价低廉,运行管理相对容易,兼具景观效果和经济效益,因此被广泛应用.
20世纪70年代起,生态浮床在国外已有诸多研究,如Audet等报道了水生植物对湖泊沉积物微生物反硝化的促进作用;Sudiarto等以猪场废水为研究对象,研究了浮叶植物大薸()对超高浓度富营养化水体总氮(TN)、总磷(TP)的去除效果,发现TN (151.67 mg/L)、TP (82.77 mg/L)的去除率分别为63.15%、36.15%. 生态浮床技术从20世纪80年代开始在我国应用,已经历30多年的发展,主要应用于城市河道和富营养化湖泊或水库. 如Yang等以模拟天津市湿地水体为研究对象,对比了北方地区常见13种湿地植物在富营养化水体中的氮磷去除效果,发现黄菖蒲(L.)、风车草(L.)、水葱()对TN、TP的去除率分别能达到90%、70%,表现出较好的去除效果;Liu等以北京南郊受污染河道为研究对象,探究2种水生植物对TN (1.86 mg/L)、TP (0.63 mg/L)的水体净化效果,结果表明,千屈菜()、黄花鸢尾 ()对COD、TN、TP的去除率分别达到75%、57%、71%和60%、49%、68%;丛海兵等以扬州市区富营养化河道杨庄河为研究对象,探究3种耐寒陆生植物对营养盐的去除速率,发现黄菖蒲(L.)、西伯利亚鸢尾()和美人蕉()五个月内对TN的平均去除速率分别为763.39、301.81和384.04 mg/(m·d),对TP的平均去除速率分别为86.92、24.91和36.6 mg/(m·d).以上研究多为应用生态浮床修复城市或乡村河道水体,也有学者开展筛选生态浮床植物优势种在富营养化湖泊和水库净化效果中的研究. 如胡晓东等以江苏省6大典型湖泊为研究对象,通过比较不同植物的氮、磷去除效果,筛选出芦苇()、茭草()、穗状狐尾藻()、轮叶黑藻()、野菱()、荇菜() 6种江苏省典型湖泊水生植物优势种; 朱洁等在白洋淀开展了种植沉水植物狐尾藻()生态浮床对湖泊深水区氮磷营养盐净化效果研究,结果表明,水体TN、TP、NO-N(亚硝态氮)去除率分别为40.54%、53.45%、99%. 然而已有研究多集中于生态浮床净化富营养化水体,缺乏对较清洁型水库水体的脱氮除磷效率及其机制研究.
千岛湖位于浙江省淳安县和建德市境内,属亚热带季风气候,四季分明,是长三角地区最大的淡水人工湖和战略水源地,千岛湖的水质安全保障事关重大. 作为一个面源污染负荷高的山区水库,流域气候和水文过程的年际波动对千岛湖水质影响较大. 近20年来,千岛湖水体有富营养化的风险,如2001-2019年TN浓度呈现升高趋势,2017-2019年TP浓度一直未能稳定达标并呈逐年增长趋势,对千岛湖水质构成威胁. 较高的营养盐浓度导致了千岛湖藻类异常生长,如2004年、2005年威坪湾出现了曲壳藻异常增殖,2007年坪山水域出现水华束丝藻异常增殖,2009年、2010年安阳水域出现曲壳藻、鱼腥藻异常增殖等现象. 因此,控制千岛湖水体氮磷浓度十分必要. 然而千岛湖流域多为山地,土地资源紧张,缺乏足够面积的净化湿地. 在氮磷浓度相对高的支汊库湾开发具有一定脱氮除磷能力的生态浮床技术,探究生态浮床技术在深水水库脱氮除磷方面的可行性,能弥补山区流域湿地净化空间不足的缺陷,减轻流域污染压力.
试验选取浮叶植物黄花水龙(Ohwi.)、沉水植物绿色狐尾藻()、挺水植物菖蒲() 3种长三角地区不同类型的典型水生植物为研究对象,通过春秋两季静态模拟试验,探讨了生态浮床对千岛湖水体的氮磷净化效果及其可能机制,筛选出适宜千岛湖生态浮床应用的水生植物,以期为后期生态浮床净化较清洁型水库水体提供参考.
1 材料与方法
1.1 研究区域
千岛湖又名新安江水库,属于大型山谷型深水水库,平均水深为31.13 m,最深处达到100 m,水体透明度、营养盐浓度、温跃层、水面开阔度不断发生变化,形成了复杂的物理、化学和生物环境梯度变化.千岛湖流域包括新安江、武强溪、富春江等30多条入库河流,采样点位于街口镇鸠坑乡一库尾湾,街口镇位于千岛湖西北上游区域,地处新安江水库库尾,是新安江水库最大入湖口.
1.2 试验材料
试验选用20 cm×30 cm×4 cm的聚苯乙烯泡沫板作为生态浮床载体,并在每个泡沫板上设置12个直径32 mm的定植孔,将植物固定器放置其中,植物覆盖率约为50%;选取12个48.7 cm×34.3 cm×25.8 cm的聚乙烯塑料水箱(见图1)作为浮床主体用于盛装试验用水.
通过查阅文献,选取长江中下游生长的3种典型
水生植物(均购置于浙江省金华市金东区竹文园艺基地)作为试验材料,其生物特性见表1.

图 1 生态浮床模拟试验装置示意Fig.1 Schematic diagram of floating bed simulation experiment device

表 1 水生植物种类及生物特性Table 1 Species and biological characteristics of aquatic plants
1.3 试验设置
试验分别于2020年10月22日-11月26日(秋季)和2021年3月27日-5月2日(春季)在浙江省千岛湖国家水质监测站进行. 试验开始前,水生植物先预培养10 d,待植物长势稳定后选取长势较好且质量相近的植物各36株,用于生态浮床模拟试验. 试验用水采自千岛湖上游街口水域,该水域在千岛湖主库区属于营养盐相对丰富的水域. 两次试验初始水样的物理化学指标如表2所示.

表 2 两次试验初始水样水质指标Table 2 Initial water quality index of two experiments
每个水箱中分别装20 L湖水,试验共设置4个处理,其中一个处理为空白对照组(仅放置泡沫板).每种植物设置3个重复,每株植物通过植物固定器固定于浮板上,自然漂浮于水箱中,水位漫过植物根部,为保证静态试验更符合自然效果,试验期间不更换试验用水,通过水位下降高度,换算污染物的去除率.
试验期间,每7 d采集表、中、底层混合水样,采集时间均为10:00. 采集完立即用0.45 μm孔径的玻璃纤维滤膜(Whatman GF/F)进行过滤,过滤后样品采用TOC分析仪测定溶解性有机碳(DOC)浓度,未过滤样品分取、冷冻,用于TN、TP浓度测定(硫酸钾-紫外分光光度法). 使用蠕动泵将表层20 cm以下的水泵入到12 mL空瓶(Labco Exetainer)中,瓶子装满时继续引流并缓慢抽出橡皮管,使液面形成凸状,然后拧紧瓶盖. 每个水箱取3个平行样,采集完后立即用注射器打入50%质量分数的ZnCl溶液终止其微生物反应并密封保存.
采集完水样先用马克笔记录水箱中的水位下降高度,再用便携式水质参数分析仪(Yellow Spring Instruments)测量所需物理指标:水温(WT)、溶解氧(DO)浓度、pH. 两季试验在开始和结束时期分别收集植物,烘干称重,过100 mm×25.4 mm筛网研磨后测量植物体内氮磷含量(意大利元素分析仪,Eurovector EA3000). 每7 d用直尺测量植物株高、根长,用电子天平(常熟意欧电子天平,DT502)称量植物干质量(生物量). 由于黄花水龙和绿色狐尾藻分别为浮叶和沉水植物,根系都较为柔软纤细,因此在测量植物的生物量和植物体内氮磷含量时不考虑地上、地下部分.
1.4 数据处理
污染物去除率()的计算公式见式(1):

式中:为 污染物起始浓度,mg/L;为水样起始体积,L;C为 第次取样的污染物浓度,mg/L;V为第次取样的水样体积,L.
污染物去除效率()的计算公式见式(2):

式中:为污染物去除效率,mg/(kg·d);为单位面积生物量,kg;为时间,d.
′
植物吸收氮去除率()的计算公式见式(3):

式中:为植物吸收氮量,g/kg;为氮去除总量,g/kg.
为了解试验系统中3种水生植物的反硝化脱氮强度,使用Kana等提出的N:Ar方法计算. 因为Ar(氩气)的化学性质是稳定的,而N(氮气)易受到微生物的影响,因此使用N:Ar方法的计算精度远高于单独使用N,N、Ar的测定仪器为膜接口进样质谱仪(Bay instrument, USA),计算方法如式(4)所示:

式中:Δ[N]为N浓度相对于自然条件下的净增量,μmol/L;[N]/[Ar]为经质谱仪矫正过的水样中N与Ar浓度比值;[N]、[Ar]分别为特定温度、盐度条件下N、Ar理论平衡浓度,μmol/L,由标准水样温度和盐度值代入Weiss方程计算得到.
试验数据采用IBM SPSS 26.0软件进行相关性和方差分析,使用单因素ANOVA检验法进行差异显著性分析(<0.05),采用Origin 2018软件作图.
2 结果与分析
2.1 水生植物生长情况
分别将3种水生植物放入塑料水箱中进行静态模拟试验,培养35 d后计算水生植物株高、根长和生物量的变化情况,结果如图2和表3所示. 春、秋季3种植物株高、根长和生物量均有增加,绿色狐尾藻和黄花水龙表现出较显著的生长优势(<0.05),且3种植物长势均为春季好于秋季.

图 2 春秋两季不同植物株高与根长净增量的变化Fig.2 Net increment of plant height and root length of different plants in spring and autumn

表 3 春秋两季不同植物生物量的变化Table 3 Changes in biomass of different plants in spring and autumn kg/m2
春季黄花水龙和绿色狐尾藻株高净增量比菖蒲高20%,秋季绿色狐尾藻株高净增量比黄花水龙高7 cm,是菖蒲株高净增量的7.4倍. 根长净增量差异更显著(<0.05),两季试验绿色狐尾藻根长净增量分别约为黄花水龙的2.2倍、菖蒲的3.2倍. 黄花水龙的单位面积生物量显著优于绿色狐尾藻、菖蒲(<0.05)(见表3). 春季试验黄花水龙、绿色狐尾藻、菖蒲单位面积生物量分别增加260%、100%、17%,秋季黄花水龙和绿色狐尾藻单位面积生物量均增加约45%,而菖蒲仅增加了约13%.
由此可见,春季黄花水龙和绿色狐尾藻表现出更顽强的生命力,更适应千岛湖水体生长,其中黄花水龙生物量增加最多,绿色狐尾藻根长增长最多.
2.2 三种水生植物对TN和TP净化效果
为比较3种水生植物氮磷去除能力,参照式(1)计算出每个处理组TN和TP的去除率,结果如图3所示. 春季黄花水龙和绿色狐尾藻对TN的去除率(简称“净去除率”)相对于空白对照组提高了约50.0%,显著高于菖蒲的TN去除率(36.8%)(<0.05);而秋季3种植物TN净去除率为22.0%~30.0%,差异不显著(>0.05). 春秋两季黄花水龙TP净去除率均显著高于其他2种水生植物(<0.05),其中春季黄花水龙TP净去除率为46.0%,绿色狐尾藻为33.1%,菖蒲为17.1%.

图 3 春秋两季不同处理组TN与TP去除率的变化Fig.3 Changes of TN and TP removal rates in different treatment groups in spring and autumn
为更精确比较3种植物氮磷的去除能力,根据单位面积生物量和时间参照式(2)计算每种水生植物单位生物量单位时间的TN和TP去除效率,结果如表4所示. 由表4可见,绿色狐尾藻、黄花水龙春季对氮磷的去除能力显著强于秋季(<0.05),去除效率约为秋季的2.0倍. 其中春季绿色狐尾藻的TN去除效率为2.89 mg/(kg·d),比黄花水龙〔2.22 mg/(kg·d)〕高23.0%,约为菖蒲〔0.27 mg/(kg·d)〕的11.0倍.

表 4 春秋两季不同水生植物TN、TP去除效率Table 4 TN and TP degradation rates of different aquatic plants in spring and autumn mg/(kg·d)
3种水生植物的TP去除效率和季节性差异均不显著,春秋两季TP的去除效率基本维持在0.01~0.08 mg/(kg·d),以上结果说明绿色狐尾藻对较清洁型水体的适应能力更强.
2.3 植物氮磷吸收情况
3种植物体内氮磷的吸收情况如表5所示. 由表5可见,绿色狐尾藻体内氮磷的吸收量显著高于其他两种水生植物(<0.05). 在氮吸收方面,两季绿色狐尾藻吸收量维持在12.44~15.57 g/kg,约为菖蒲(5.45~8.59 g/kg)、黄花水龙(5.27~8.30 g/kg)的2.0倍.在磷吸收方面,3种植物均表现为春季高于秋季. 绿色狐尾藻的氮吸收量(0.96~1.95 g/kg)最高,黄花水龙(0.51~0.67 g/kg)次之,菖蒲(0.24~0.68 g/kg)最低.

表 5 春秋两季不同水生植物体内氮磷吸收量Table 5 Nitrogen and phosphorus uptake by different aquatic plants in spring and autumn
为探究生态浮床植物氮去除的机制,根据式(3)计算出植物吸收、反硝化与其他作用对氮去除的贡献率,结果如表6所示. 从表6可以看出,植物吸收去除氮的占比为20%~30%,不同水生植物对氮的去除主要依靠的是微生物反硝化与其他作用.
2.4 春秋两季不同处理组溶解性N2增量变化情况
为更进一步探究3种水生植物反硝化作用的差异,参照式(4)计算了3种水生植物溶解性N增量(简称“Δ[N]”)的变化情况,以此来比较3种水生植物的反硝化作用,结果如图4所示. 春秋两季试验中3种水生植物Δ[N]整体都高于空白对照组,且均呈先升后降的趋势,春季试验3种水生植物与空白对照组的Δ[N]差值均高于秋季. 3种水生植物中,绿色狐尾藻的Δ[N]相对于空白对照组增量最多,其中秋季11月12日Δ[N]比空白对照组高13.61 μmol/L,黄花水龙、菖蒲与空白对照组Δ[N]差值(简称“净脱氮差”)分别为3.55和-0.26 μmol/L. 春季4月25日绿色狐尾藻的净脱氮差值为24.63 μmol/L,黄花水龙和菖蒲分别为22.35和15.75 μmol/L. 春秋两季3种水生植物的净脱氮差范围分别为-4.14~24.63 μmol/L(绿色狐尾藻)、0.16~22.35 μmol/L(黄花水龙)、-0.26~15.74 μmol/L(菖蒲). 以上结果说明绿色狐尾藻根系的微生物反硝化作用最强,黄花水龙次之,菖蒲表现最差.

表 6 不同途径脱氮的贡献率Table 6 Contribution rate of nitrogen removal by different ways

图 4 春秋两季不同处理组Δ[N2]的变化Fig.4 Changes of Δ[N2] in different treatment groups in spring and autumn
2.5 水生植物净脱氮量与不同因素相关性分析
为进一步探讨影响水生植物反硝化作用的主要因素,为后期示范工程筛选植物提供参考. 笔者对3种水生植物净脱氮量与5种主要因素的相关性进行了分析(见表7). 结果发现3种水生植物的净脱氮量与各因素之间的相关性存在显著差异,其中黄花水龙和菖蒲净脱氮量与生物量均呈显著正相关(<0.05),绿色狐尾藻净脱氮量与生物量呈极显著正相关(<0.01);绿色狐尾藻脱氮量与根长、温度也均呈显著正相关(<0.05),而黄花水龙和菖蒲净脱氮量与根长、温度均无显著相关性;黄花水龙、菖蒲净脱氮量与DO浓度均呈显著负相关(<0.05),而绿色狐尾藻净脱氮量与DO浓度并无显著相关性;3种植物净脱氮量与DOC浓度均无显著相关性. 结果表明,生物量对3种水生植物的反硝化作用影响最为显著.
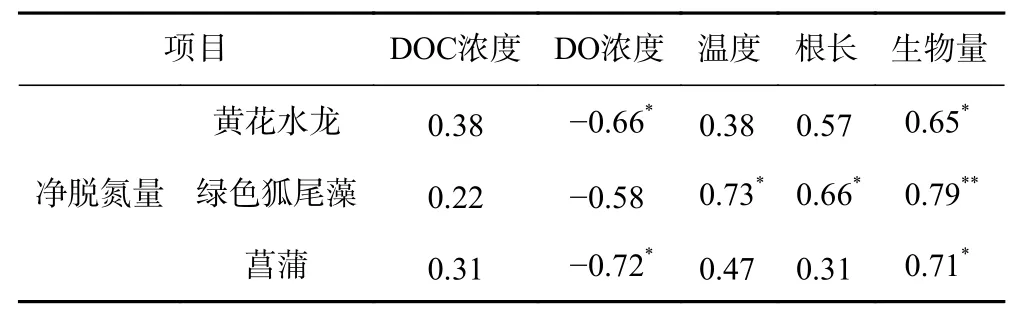
表 7 水生植物净脱氮量与不同因素的相关分析Table 7 Correlation analysis between net nitrogen removal of aquatic plants and different factors
3 讨论
3.1 水生植物净水能力差异分析
在春秋两季试验中发现,春季植物生长情况和氮磷去除效果显著优于秋季,原因与温度有关(秋季温度为10.0~20.1 ℃,春季温度为17.1~28.3 ℃),温度过高或过低都会影响植物生理特性,一定温度范围内,根际微生物活性也会随着温度的升高变强. 试验中,植物生物量的变化情况存在显著性差异(见表3). 植物生物量的大小主要取决于植物自身器官细胞特性、可利用营养盐物质和对环境的适应能力,黄花水龙在生长期间会长出许多向上生长的白色粗棒状不定根,导致其生物量增加最多. 一般来说,植物在污染物中生物量增加表明其对氮磷营养盐胁迫具有很强适应性,这与该试验生物量增加较多的绿色狐尾藻和黄花水龙较强的去氮除磷能力相吻合(见表4).
不同水生植物对水体氮磷去除效果有显著性差异,这种种间差异主要与植物类型及其生理特性有关.绿色狐尾藻属于广泛生长于热带亚热带地区的多年生沉水植物,其根茎发达密集,适宜在16~26 ℃温度条件下生长,且根系不断向水下蔓延,为微生物、细菌提供了良好的繁衍场所,植物根际借助适宜的温度、氧气逐渐形成硝化和反硝化微生物群落,因此绿色狐尾藻对氮磷有较强的净化效果. 绿色狐尾藻ΔN最高(见图4),表明绿色狐尾藻具有相对较强的反硝化脱氮能力,张芳等研究也发现沉水植物溶解性N的释放通量显著高于其他类型水生植物. 黄花水龙对水体氮磷有较明显的去除效果. 黄花水龙的快速繁殖需要从水体中吸收氮磷等营养物质合成组织细胞,但其根际微生物硝化反硝化作用弱于绿色狐尾藻. 黄花水龙属于浮叶植物,根茎不会完全往水下延深,根际区附着的微藻、微生物等物质数量和种类均少于绿色狐尾藻,微生物硝化反硝化作用较低.菖蒲属于不耐寒的挺水植物,最适宜的生长温度为20~25 ℃,因此对氮磷的去除效果低于其他两种植物,且生物量变化不大. 邵凯迪等研究5种水生植物对模拟菜地径流TN去除效果时发现,黄花水龙和绿色狐尾藻对氮的去除效果显著高于其他3种植物,且二者都具有较大生物量.
值得注意的是,3种植物ΔN均呈现先升后降的趋势(见图4),原因可能与水体中氮浓度的下降有关.随着微生物反硝化作用所需的底物硝态氮浓度降低,微生物反硝化作用也随之减弱.
3.2 不同途径脱氮的贡献
生态浮床脱氮途径主要包括植物吸收、氨氮挥发和根际微生物反硝化等作用. 氨氮挥发过程中,当水体pH>8.0、氨和铵根离子比例为1:1时,氨氮挥发造成的氮损失才较为显著. 该试验中pH一直维持在6.8~7.5,因此氨氮挥发造成的氮损失可忽略不计.植物吸收对浮床系统去除水体氮素具有重要意义,植物吸收氮的能力与其根系发达程度、生物量高低及生长代谢能力有关,绿色狐尾藻茂密的根系和较高生物量使得植物体内吸收的氮磷含量最多(见表5).刘少博等研究发现,不同浓度NH-N条件下,绿色狐尾藻对氮的吸收最高(30.7~53.4 mg/g),表现出超高的氮吸收能力. 试验后期植物腐烂衰败重新进入水体导致水体营养盐浓度升高,后期示范工程应用需要采用定期刈割植物的方式带走水体中的氮. 除了植物吸收,微生物反硝化作用也是系统去除氮素的重要途径. 植物根系分泌物能够调节根际区微生物群落的多样性,使得根际区聚集大量硝化反硝化细菌. 该研究中3种水生植物微生物反硝化与其他作用约占氮去除贡献率的70%~80%(见表6),与已有研究报道微生物反硝化作用对氮的去除贡献率为60%~90%相一致. 孙鹏等研究3种植物对氮素富集能力时发现,美人蕉、石菖蒲和伞草氮素累积分别占系统去除率的37.4%、33.3%、27.5%,表明微生物反硝化作用对氮的去除占主导作用.
影响水生植物反硝化作用的主要因素有很多,笔者做了3种水生植物净脱氮量与DOC浓度、DO浓度、温度、根长、生物量5种主要影响因素的相关性分析(见表7). 结果表明,3种水生植物净脱氮量与生物量呈显著正相关,是影响水生植物反硝化作用的最关键因素. 生物量越大不仅能附着更多硝化反硝化细菌、微生物,而且粗壮的枝叶能增加植物覆盖率,减少植物光合作用,间接为反硝化细菌提供更多厌氧区域. 除了生物量,DO浓度在微生物反硝化活动过程中起着重要作用,当氧气大量存在时,电子受体所产生的能量会抑制微生物反硝化作用. 在该研究中黄花水龙和菖蒲净脱氮量与DO浓度呈显著负相关(<0.05),说明水生植物反硝化作用受DO浓度影响较大. Veraart等在研究水生植被对水体反硝化作用的影响时发现,在DO浓度较低的浮叶水生植物区域反硝化速率最高,因此过高的DO浓度会抑制水生植物反硝化作用.
综上,后期示范工程筛选植物时应致力于选取生物量大、耐高低温、根系发达的水生植物.
4 结论
a) 黄花水龙、绿色狐尾藻、菖蒲3种水生植物均能在千岛湖湖水中生长,春季植物生长情况显著优于秋季. 黄花水龙生物量增长最为显著,其次是绿色狐尾藻,菖蒲最少.
b) 绿色狐尾藻的净化水质能力强于黄花水龙、菖蒲. 绿色狐尾藻的TN去除效率为2.89 mg/(kg·d),比黄花水龙高23.0%,是菖蒲TN去除效率的11.0倍. 黄花水龙、绿色狐尾藻的TP去除效率维持在0.07~0.08 mg/(kg·d),菖蒲的TP去除效率仅为0.02 mg/(kg·d).绿色狐尾藻体内吸收的氮、磷含量最多,且反硝化脱氮能力最强.
c) 微生物反硝化作用是该试验生态浮床脱氮的主要途径,生物量是影响该试验水生植物反硝化作用的最关键因素.
d) 通过3种水生植物脱氮除磷能力、反硝化作用、植物长势等方面对比分析,绿色狐尾藻在该次试验中氮磷净化效果最好.
猜你喜欢
今日农业(2022年14期)2022-11-10
今日农业(2022年14期)2022-09-15
今日农业(2022年3期)2022-06-05
国际商业技术(2022年4期)2022-04-21
黄河之声(2020年16期)2020-11-05
今日农业(2020年16期)2020-09-25
环球时报(2020-06-30)2020-06-30
旅游(2018年6期)2018-06-08
知识窗(2017年10期)2017-11-02
中华手工(2016年9期)2016-11-10
