
蓝藻拟柱孢藻(Cylindrospermopsis)和尖头藻(Raphidiopsis)的分与合
2022-04-14 14:07:10李仁辉程耀耿若真程新良肖鹏张和刘洋马增岭
河南师范大学学报(自然科学版) 2022年3期
李仁辉,程耀,耿若真,程新良,肖鹏,张和,刘洋,马增岭
(1.温州大学 生命与环境科学学院,浙江 温州 325035;2.杭州市淳安生态环境监测站,杭州 311700;3.河南师范大学 生命科学学院,河南 新乡 453007)
蓝藻的现代分类学进入21世纪后发展迅速,除了高阶层分类系统在不断修订之外,新的分类单位的发表数量也是非常之多.以2个常形成水华的蓝藻属的合并为例,从蓝藻分类系统的变化和发展、现代蓝藻分类学的核心以及蓝藻分类系统修订的评估等方面进行论述和介绍.
1 蓝藻的多样性和分类系统
蓝藻的多样性以及进化是早期地球生命研究的一个重要内容.由于蓝藻的原核细胞特性,这种简单的特性反而使蓝藻的分类相当困难[1].蓝藻长期复杂的进化历程使其很难简单地从形态来辨别.在过去的蓝藻分类系统中使用了许多形态特征,如藻体外形、极性、藻丝的分枝类型,藻体和细胞的大小,异形胞和厚壁孢子的有无等来进行高等级的分类,但是这些特征经常在蓝藻的进化过程中多次出现和丢失[2-3].同时蓝藻的分类过程也一直在植物学和细菌学分类方法中争论、比较、妥协和采纳,结果就是蓝藻的分类从早期的主要依照形态学分类发展到现代的多相特征(Polyphasic),或者整合(integrated)特征的分类方法,即通过培养藻种进行形态学、生理学、生化以及分子遗传学特征相结合的综合分类方法[4].正式的蓝藻分类系统从初始建立(1932年)至今也不到90年,特别是近几十年,同时使用植物学分类系统和细菌学分类系统.目前把植物学分类法和细菌学分类法结合起来的方式也很流行,因为这种结合被认为更适用于蓝藻的分类系统.
1932年,捷克的蓝藻分类学家GEITLER提出较为体系化和被认可的形态分类系统,将蓝藻分成3个目[5]:不产内生孢子单细胞的色球藻目(Chroococcales)、产内生孢子单细胞的管孢藻目(Chamaesiphonales)以及所有的丝状体蓝藻藻殖段目(Hormogonales).10年后他本人又将蓝藻的分类系统修订成4目系统[6]:色球藻目(Chroococcales)、皮果藻目(Dermocarpales)、宽球藻目(Pleurocapsales)和藻殖段目(Hormogonales),这一分类系统也持续了近20年.1959年印度蓝藻分类学家DESIKACHARY提出了真分枝的丝状蓝藻从藻殖段目分离出来单独成立真枝藻目(Stigonematales),其后的学者也同意这种分离并且把不分枝的丝状蓝藻也单独成立为一个目(当时叫念珠藻目Nostocales),再到后来提出了不产生异形胞的丝状蓝藻成立颤藻目(Oscillatoriales)[7].所以20世纪70到80年代基本上认可的是蓝藻5目分类系统[8]:色球藻目(Chroococcales)、宽球藻目(Pleurocapsales)、颤藻目(Oscillatoriales)、念珠藻目(Nostocales)和真枝藻目(Stigonematales).同样根据细菌学分类方法也是把蓝藻(蓝细菌)分成和上述蓝藻形态系统的5个目对应的5组(Section I-V)[9].
随着电子显微镜技术和DNA分子序列的应用,给蓝藻的分类系统、发育分析和进化提供了大量新信息.这些新的证据表明,在整个蓝藻分类系统中,5个目中的每个目都不是单系统:色球藻目、宽球藻目和颤藻目在分子系统关系上相互混合;被认为是单系统的具异形胞的念珠藻目也不能和具有分枝的丝状体真枝藻目完全分开,反而组成了一个更大的混合大簇[10].所以,HOFFMANN等[11]在2005年提出了一个新的高层次蓝藻分类系统,把蓝藻纲下面分成4个亚纲:粘菌藻亚纲(Gloeobacteriophycidae)、聚球藻亚纲(Synechococcophycidae)、颤藻亚纲(Oscillatoriophycidae)和念珠藻亚纲(Nostochopycidae),亚纲下面分成6个目的系统.这个新的系统颠覆了很多以前的系统和认知,主要是根据DNA序列的证据,以及细胞的超微结构(类囊体排列的方式)把单细胞的聚球藻目(Synechococcales)和丝状体的假鱼腥藻目(Pseudanabaenales)合在一起归在聚球藻亚纲(Synechococcophycidae),而另外一类单细胞大类色球藻目(Chroococcales)和丝状体类的颤藻目(Oscillatoriales)又合在一起归在颤藻亚纲(Oscillatoriophycidae).这个新的系统基本上实现了目水平上的单系统,但是目以下的科和相当数量的属并不是单系统.
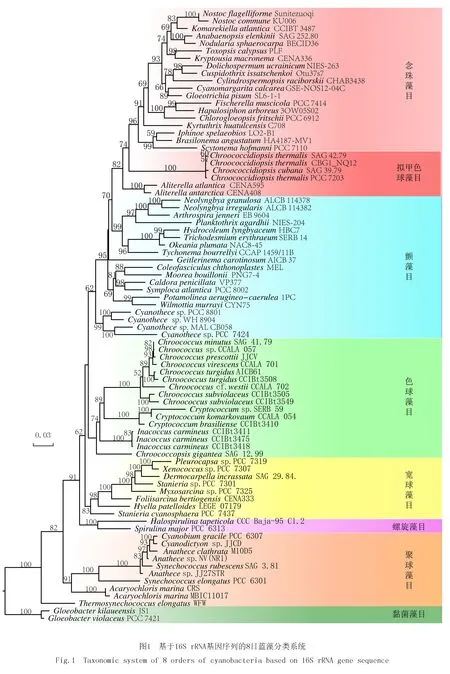
2 现代蓝藻分类系统中属的概念(concept of cyanobacterial genera)的确定
2014年的蓝藻分类系统的特色是以蓝藻的属水平分类为重心的分类系统.KOMREK等[1]根据现代标准提出的蓝藻属的概念包括下面3个方面:1)蓝藻的属在分子系统中要形成独特的一枝,并且同关系最近的属的16S rRNA基因序列的相似度要小于95%;2)蓝藻的属和相近的属要具有形态上可以辨认的或其他生物学特征;3)具有明显的生态习性等生态学差异.通常蓝藻的属应该是在形态、生理生态和分子等综合特征上所代表一个独特的类群.所以,现代蓝藻分类的研究方法都是大力提倡基于多相分类特征的分类(Polyphasic taxonomy).这个蓝藻属的概念及其标准的提出加快了蓝藻新属的建立速度.结果在2014-2020年期间,蓝藻新属的数量又增加了70多个,这些新属都是合法的,是被承认的[12].蓝藻新属的创建大部分的情况还是针对一些种类数目较多的大属,并且已有分子系统证据显示这些大属是多系统的.所以,从这些大的属剥离出许多新的蓝藻属是很常见的操作,例如鞘丝藻属(Lyngbya)剥离,已经成立了9个新属.还有一种分类系统调整的情况,就是2个属或者几个属之间合并成1个属,最主要的原因就是这些属间的DNA分子序列的高度相似性.本文就是要叙述丝状蓝藻拟柱孢藻属(Cylindrospermopsis)和尖头藻属(Raphidiopsis)从单独的属到最近合并成尖头属的变迁.
3 拟柱孢藻属和尖头藻属
3.1 拟柱孢藻属和尖头藻属的介绍
拟柱孢藻属(Cylindrospermopsis)在蓝藻分类上归属于念珠藻目(Nostocales),束丝藻科(Aphanizomenonaceae)(束丝藻科也是2014年新的蓝藻系统提出来的新科,此科包含能形成水华的所有具有异形胞的属,如长孢藻(Dolichospermum),束丝藻(Aphanizomenon),节球藻(Nodularia)等).拟柱孢藻是1972年从项圈藻属(Anabaenopsis)分离出来,以拉氏拟柱孢藻(Cylindrospermopsisraciborskii)(原拉氏项圈藻Anabaenopsisraciborskii)作为模式种的单种属[13].拟柱孢藻属同项圈藻属的区别在于端生异形胞的形成方式不同,即拟柱孢藻的端生异形胞是原生形成(Primary).而项圈藻的端生异形胞是通过藻丝间生的异形胞分裂后的双异形胞中间断裂而成,所以又称次生形成方式(Secondary).自拉氏拟柱孢藻建立后,从亚洲、美洲及非洲又陆续发表了C.philippinensis,C.africana,C.cuspis,C.catemaco和C.curvispora,近年来又从欧洲发表了一些新种,使拟柱孢藻属的种类数达到11个[14-15].尖头藻属(Raphidiopsis)是1929年由FRITSCH和RICH建立,以弯曲尖头藻(R.curvata)为模式种的浮游性丝状蓝藻[16].尖头藻属的形态特点是单藻丝体,藻丝的一头或两头渐尖.它的最特别之处就是藻体在整个生活史阶段不形成异形胞但形成休眠孢子,在最新的蓝藻分类系统中还是将尖头藻属归属到念珠藻目的束丝藻科.
到目前为止,世界上共报道7种尖头藻,除了模式种外,还有地中海尖头藻(R.mediterranea),印度尖头藻(R.indica)和中华尖头藻(R.sinensis)等(表1)[17-19].拟柱孢藻在形态上同尖头藻较为相似,特别是当藻丝上异形胞和休眠孢子没有形成的情况下,更加难以辨别.有趣的是,拟柱孢藻和尖头藻占有几乎完全相同的生态位,也就是说拟柱孢藻和尖头藻常常共存于同一自然水体中.本团队于2001年首次在尖头藻(弯形尖头藻)中发现了产拟柱孢藻毒素以及其衍生物脱氧拟柱孢藻毒素(Deoxyclindrospermopsin)[20-21].最近澳大利亚的学者也发现了地中海尖头藻可形成拟柱孢藻毒素及脱氧拟柱胞藻毒素[22].由于拟柱孢藻和尖头藻所具有的形态、生态习性以及产毒特性的相似性,导致这2个属的区分非常困难,也直接造成了这2个属的分类混乱.
3.2 拟柱孢藻属和尖头藻属的异同性
基于DNA分子序列的研究表明,拟柱孢藻和尖头藻在16S rRNA基因、16S rRNA和23S rRNA基因的间隔区(ITS)以及rpoC1基因的序列都是高度相似且无法将这两属分开[23].这种属间分类界限的不明确性已经使研究者对尖头藻属的合理性产生怀疑:澳大利亚学者认为尖头藻是属于环境中拟柱孢藻的一种形态型[24];希腊学者根据Kastoria湖中拟柱孢藻的形态和16S rRNA基因的证据,认为地中海尖头藻是拉氏拟柱孢藻复杂生活史中的非异形胞时期[25].以色列学者把Kinneret湖的拉氏拟柱孢藻的形态分为6类,其中就有一类是尖头藻的形态[26].STUCKEN等[27]对拉氏拟柱胞藻(C.raciborskiiCS-505)和布氏尖头藻(R.brookiiD9)全基因组序列比较后,发现它们两者共有的2 500多个基因的相似度大于90%.但是,GUGGER团队和本团队的研究结果都表明,尖头藻在结构上不含异形胞,不能进行固氮,所以在缺氮的培养基中不能生长.而拟柱孢藻则可以在无氮培养基中生长[2].全基因组分析结果表明上述的布氏尖头藻(R.brookiiD9)同拉氏拟柱孢藻(C.raciborskiiCS-505)相比,它的基因组里丢失了15 kb的固氮基因簇(nif)和一些重要的辅酶和氨基酸转运蛋白[27].这也验证了GUGGER等[2]的研究,表明他们在非洲分离的尖头藻并不能检测出nifH基因.我们对中国的尖头藻纯藻株的基因检测也没有发现nifH基因[28].

表1 拟柱孢藻和尖头藻合并前后的藻类命名变化
3.3 拟柱孢藻属和尖头藻属的合并过程
从上面的介绍和一些研究结果看出,拟柱孢藻和尖头藻在形态特征、生态学和DNA序列上的相似性很高,所以研究人员一直在寻找一些分子标记(markers)来快速辨别这2个属.除了利用16S rRNA,ITS和rpoC1基因序列外,WU等[29]用了7个基因的整合序列对中国12株拟柱孢藻和9株尖头藻进行分子系统分析,结果也表明拟柱孢藻和尖头藻是一个相互混合的簇(cluster).为解决藻株使用量的问题,LI等[23]利用从中国分离的44株拟柱孢藻和16株尖头藻,使用16S rRNA,cpcBA-IGS,rpoC1和ITS-L基因序列分别进行分子系统分析,发现前3个基因(16S rRNA,cpcBA-IGS,rpoC1)还是无法分开这两个属,但是ITS-L可以把这60个中国藻株分为明显的两个簇,分别对应拟柱孢藻属和尖头藻属.并且发现了ITS-L的7个碱基(RAGAAACT)可作为尖头藻的保守序列,是鉴别尖头藻的分子标记.这个结果非常振奋,是拟柱孢和尖头藻分子鉴别的重要进展,但是,这个结果只是使用中国的大量藻种,并没有涉及世界其他区域的藻种.
2018年,AGUILERA等[30]通过对阿根廷浅水湖泊分离的两株地中海尖头藻(R.mediterranea)藻种的多相分类研究,并且使用了大量世界各个地(包括许多中国的藻株)的拉氏拟柱孢藻和部分尖头藻藻种的16S rRNA,16S-23S ITS和cpcBA-IGS的基因序列,进行分子系统学分析.这种大量藻株分析的结果还是同样表明,拟柱孢藻和尖头藻在分子系统中无法分开,特别是ITS的二级结构也显示2个属高度一致,并且特别强调ITS-L并不能区分中国以外的藻株,7个特征碱基(RAGAAACT)并不是尖头藻所特有的,许多欧洲的拉氏拟柱孢藻种也含有这7个碱基.最重要的是,AGUILERA等人利用本研究的结果,把拟柱孢藻和尖头藻2个属合并为1属.因为尖头藻属成立时间比拟柱孢藻属要早,具有优先权,所以最后2个属合并成尖头藻属,模式种还是弯形尖头藻.至此,合并后的尖头藻属是单系统了,目前在藻类最有权威的种属库Algaebase中共有18个种类[31].这个属的合并是合法的,是被承认的.
4 拟柱孢藻属和尖头藻属合并后的影响和评价
最近几十年蓝藻分类系统在各个分类阶层都在不断变化,在属水平上更加突出.但是拟柱孢藻属和尖头藻属的合并可以算得上是蓝藻分类史上的一个较大的事件,虽然这只是在属水平的调整,这2个属特别是拟柱孢藻属是近年来受关注度非常高的蓝藻.它是水华蓝藻,又是产毒和入侵蓝藻.它们的合并对于蓝藻水华研究中使用哪个名字成为大家首先遇到的问题.现在已经有很多拟柱孢藻的研究接受这个新的命名,投稿过程中也有部分审稿人要求作者将拟柱孢藻改为尖头藻.我们再次强调,这种合并是合法的,在形式上是按照植物学命名法进行的,现在也是被承认的.但是对于这种合并或者分类处理是否合理,学者们可以评价和讨论.我们认为这种合并不是很恰当,因为它首先就没有遵循KOMREK等[1]提出的蓝藻属的概念.按上述介绍,一个蓝藻属应该是在分子系统,形态学,以及生物学和生态学等综合方面的独立体.现在拟柱孢藻和尖头藻的合并只是强调了DNA分子序列和生态位等的高度相似性,而对藻丝中异形胞的形成与否这么重要的生物学结构特性完全忽略,这是不合适的.异形胞的特征在蓝藻的分类系统中经常影响目水平以上的分类阶层,为什么这种属水平的合并却遭到忽略?原因可能是很多场合下特别是野外条件下,异形胞不形成而无法观测到.为此,KOMREK[12]指出了现代DNA测序技术以后建立的新的蓝藻属和蓝藻种的诸多问题,其中就有拟柱孢藻和尖头藻的合并问题.理由也是对决定性的特征(异形胞形成)的忽略,还有合并研究的材料尖头藻使用的并不是模式种,而拟柱孢藻属有10多个种类,而合并研究中只是用了拉氏拟柱孢藻一个种的藻株,这种合并缺乏整个属的代表性.但是这些评论和批评目前还不能改变这种合并的合法性,除非找到新的证据和新的markers来证明这2个属的明显差异.期待蓝藻分类学在不断发展的过程中,新方法和新技术带来更多令人振奋的结果,为深入认识自然和服务人类做出贡献.
猜你喜欢
可再生能源(2022年8期)2022-08-17 06:37:52
法医学杂志(2022年1期)2022-06-21 01:23:34
大自然探索(2019年2期)2019-03-01 02:23:30
生态学报(2018年14期)2018-08-29 11:08:32
中国环境科学(2018年7期)2018-07-26 09:03:50
法医学杂志(2016年5期)2016-11-21 01:44:43
分析测试学报(2015年9期)2015-12-17 16:44:27
中国环境科学(2015年6期)2015-11-19 08:39:58
小资CHIC!ELEGANCE(2014年9期)2015-03-16 09:56:41
水生生物学报(2015年1期)2015-02-28 15:59:57
