
柠条堆肥与耕作深度对连作黄瓜土壤细菌群落组成与代谢功能的影响
2022-04-13 07:29张凯歌兰挚谦付玉芳王晓卓张雪艳
植物营养与肥料学报 2022年3期
张凯歌,兰挚谦,付玉芳,王晓卓,张雪艳
(宁夏大学农学院,宁夏银川 750021)
柠条(Caragana microphylla)作为防风固沙、保持水土的功能作物被广泛种植于我国西北地区,根据生长情况需要定期进行平茬,据统计仅宁夏全区每年产生超过80万t柠条残茬废弃资源[1]。柠条残茬堆肥化处理作为资源再利用的有效途径越来越被人们广泛关注。Zhang等[2]研究发现,柠条堆肥氮素和有机质含量高,在长期连作的黄瓜土壤中连续施用可显著增加根际土壤碳矿化,促进黄瓜根系发育。但柠条堆肥的高效施用尚缺乏系统性的研究。
有机肥常以底肥方式施入农田,不合理的施用伴随着对土层过度的侵扰。免耕作为一种保护性耕作方式,减少了对土壤团粒结构的破坏,促进了生态环境的稳定性[3],但长期免耕会导致土壤固、液、气失衡,需要深翻打破犁底层,协调土壤中的水、肥、气、热,以改善土壤环境,促进土壤微生物群落多样性和稳定性[4–6]。耕作方式对农田土壤环境的影响在小麦、玉米、棉花等大田作物上多有研究[7],而设施土壤上的研究较少。
因此,本研究以柠条堆肥与不同耕作深度配合,探讨对土壤养分和微生物多样性等关键指标的影响,以期为柠条残茬废弃资源高效再利用,及设施农田生态可持续发展提供理论依据和数据支撑。
1 材料与方法
1.1 试验材料与设计
试验在宁夏园艺产业园4号日光温室进行,土壤质地为粉壤土(含砂粒23.5%,粉砂52.8%和黏粒3.7%),土壤速效氮、磷、钾含量分别为10.92、38.52、49.95 mg/kg,有机质含量为12.83g/kg。该区域海拔 1110.14 m,位于东经 105°53′—106°36′,北纬 38°26′—38°48′之间,属于中温带干旱气候,平均温度6.28℃~13.1℃,年降雨量为129~600 mm。本试验于2018年2月至2020年1月连续进行了4茬,供试黄瓜品种为‘博美626’(天津德瑞特种业有限公司)。试验以鸡粪+翻耕15 cm为对照(CK),施用柠条堆肥分别配合免耕(T0)、翻耕15 cm(T15)、翻耕 35 cm (T35)、翻耕 45 cm (T45),共 5 个处理。柠条、玉米秸秆、羊粪以质量比1∶2∶6混合作为堆肥原料,碳氮比 25∶1,堆肥50 天后即为柠条堆肥。柠条堆肥的理化性状为:容重0.25 g/cm3、总孔隙度 79.2%、全氮 29.70 g/kg、全磷 18.60 g/kg、全钾18.60 g/kg、有机碳475 g/kg。鸡粪全量氮、磷、钾含量依次为30.80、13.40、2.32 g/kg,有机碳含量为312 g/kg。5个处理的氮素投入量相等,鸡粪施用量为 22.5 t/hm2,柠条堆肥施用量为 23.2 t/hm2,连同磷酸二铵450 kg/hm2和复混肥(N∶P∶K=20∶20∶20) 450 kg/hm2,在黄瓜移栽前一次性配合耕作作底肥施用。黄瓜采用高畦栽培,双行种植,株距33 cm,行距70 cm。为防止处理间水分横向运移,小区之间用泡沫板进行隔离,泡沫板埋入深度为80 cm。每个小区面积9 m2,每个处理3次重复随机排列,进行统一追肥和灌水。
1.2 土壤样品的采集与分析
在第4个栽培茬的盛果期(定植后2.5个月),采用5点取样法在每个处理每个重复的栽培畦中部(避开温室最北部和最南部)取0—20 cm土层样品。采用四分法将混合的土壤装入自封袋后用冰盒带回实验室,新鲜土壤中剔除石子等杂物后,土壤过2 mm筛后,保存于–80℃冰箱中,用于土壤微生物群落多样性分析。另一部分土壤风干磨碎过1 mm筛后用于土壤养分含量的测定。
1.2.1 土壤养分 土壤pH采用电位计法(上海雷磁,PHS-3E),EC值采用电导法(上海雷磁,DDS-307),有机质含量采用重铬酸钾—硫酸氧化法;全氮含量采用H2SO4消化—半微量凯氏定氮法;速效氮含量采用K2SO4浸提—流动分析仪进行测定[8]。
1.2.2 微生物群落多样性 采用Biolog-ECO技术,称取相当于10 g烘干土于250 mL锥形瓶中,加入 90 mL 灭菌去离子水,震荡 1 h (200 r/min,4℃),静置30 min后,取上清液用灭菌去离子水稀释1000倍,吸取 150 μL加至 Biolog-ECO 板的微孔中,25℃下暗室培养,每24 h在590 nm下测定吸光值,以每孔平均颜色变化率 (average well color development,AWCD) 作为微生物活性的有效指数,其公式为:
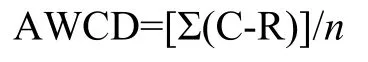
式中,C为微平板含有31种碳源每孔读数,R表示对照微孔读数,n表示96孔板中碳源底物的种类。
1.2.3 土壤微生物DNA的提取及测序 微生物宏基因组测序由上海派森诺生物科技有限公司完成。土壤样品微生物总DNA利用快速提取试剂盒 (MP Biomedicals, Santa Ana, CA, USA)提取。利用 1% 琼脂糖凝胶电泳检测抽提的基因组 DNA 质量,用超微量分光光度计 (NanoDrop 1000,USA) 测定提取的DNA 浓度。采用通用的正向引物 338F (5′-ACTCC TACGGGAGGCAGCA-3′) 和反向引物 806R(5′-GGACTACHVGGGTWTCTAAT-3′) 对细菌 16S rRNA基因V3~V4区进行PCR扩增。PCR扩增程序如下:初始变性 2 min (98℃),变性 30 s (95℃),退火 30 s (55℃),延伸 45 s (72℃),进行 27 个循环,然后稳定延伸 10 min (72℃)。用 2% 琼脂糖凝胶电泳对 PCR 扩增产物进行检测。对扩增产物切胶回收,用 QuantiFluorTM 荧光计进行定量。将纯化的扩增产物进行等量混合,连接测序接头,合格后采用Illumina Miseq 平台 (Illumina, Inc., San Diego, CA,USA) 对DNA片段进行双端(Paired-end)测序。利用Vsearch软件对所有样品序列进行去引物片段,在98%相似度水平对去重后的序列聚类后输出分类操作单元 (operational taxonomic units,OTUs),使用QIIME2在Greengenes数据库进行比对得到 OTUs 的分类学信息,得出物种对应的基因丰度和分类学水平上各样品中物种的丰度,从而构建相应分类学水平上的丰度谱,并计算Chao1丰富度估计量、Shannon多样性指数和Simpson指数及Pielou’s 指数。
1.3 数据统计
使用 SPSS 24 软件,进行单因素 ANOVA 分析;用Origin 2018制作相关图表。利用qiime2云平台 (https://view.qiime2.org/)进行 α 多样性分析 (包括Chao1、Shhanno、Simpson 等指数),使用 R 软件对环境因子、细菌群落和功能、微生物碳代谢功能指标进行冗余分析 (RDA分析)。
2 结果与分析
2.1 不同处理对黄瓜产量的影响
4个柠条堆肥处理的黄瓜产量与CK无显著差异,黄瓜产量随耕作深度增加而下降,免耕和浅耕处理(T0和T15)的黄瓜产量显著高于深翻(T45)处理(图1)。

图1 不同处理的黄瓜产量Fig. 1 Cucumber yield of different treatments
2.2 不同处理对土壤理化性质的影响
施肥和耕作深度的改变影响了土壤pH、EC值、碳氮比和速效氮含量(图2)。柠条堆肥处理中土壤pH随耕作深度增加而上升,T0、T15和CK处理差异不显著;T0、T15和T35处理土壤EC值和速效氮含量显著高于CK,而T45处理速效氮含量显著低于CK处理。与CK相比,柠条堆肥配合免耕和浅耕显著降低了土壤碳氮比,而T35和T45处理的碳氮比保持稳定。
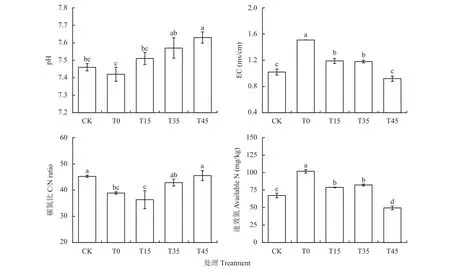
图2 不同处理下土壤pH、EC、碳氮比和速效氮含量Fig. 2 Soil pH, EC, C/N ratio and available nitrogen content under different treatments
2.3 不同处理对土壤细菌群落多样性的影响
各处理 Good’s coverage 指数均大于 0.96,表明本研究测序深度满足分析要求(表1),柠条堆肥各处理的 Good’s coverage 指数均高于 CK。细菌群落丰富度、多样性和均匀度指数均随耕作深度增加呈先上升后降低的趋势,T35处理最高。除T0外,其他处理 Shannon、Simpson 和 Pielou’s 指数均高于 CK,且深翻处理均显著高于T0 (P<0.05),说明柠条堆肥施用和深翻处理均有利于农田微生物群落多样性发展。
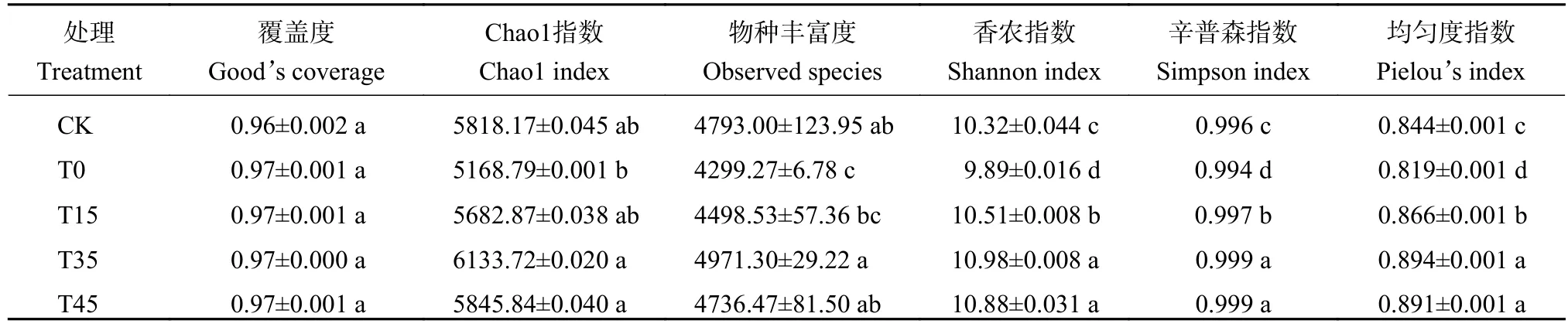
表1 不同处理土壤细菌群落丰富度、多样性和均匀度指数Table 1 Soil bacteria species richness, diversity, and evenness index under different treatments
放线菌门(Actinobacteria,40.77%)、变形菌门(Proteobacteria,26.26%)、绿弯菌门 (Chloroflexi,10.50%)相对丰度大于10%,为各处理中土壤优势菌门。放线菌门(Actinobacteria)的相对丰度随耕作深度的增加显著下降,而变形菌门(Proteobacteria)、绿弯菌门(Chloroflexi)、酸杆菌门(Acidobacteria)、芽单胞菌门(Gemmatimonadetes)的相对丰度与放线菌门相反,T35处理的酸杆菌门(Acidobacteria)、杆菌属菌门(Patescibacteria)、Rokubacteria门相对丰度最高;T15较CK显著提升了变形菌门(Proteobacteria)、绿弯菌门(Chloroflexi)、Deinococcus-Thermus门、杆菌属菌门(Patescibacteria)的相对丰度度;T0处理土壤放线菌门(Actinobacteria)、厚壁菌门(Firmicutes)和Deinococcus-Thermus门的丰度显著高于T15处理(分别提高26.38%、295.83%和52.29%,图3a)。
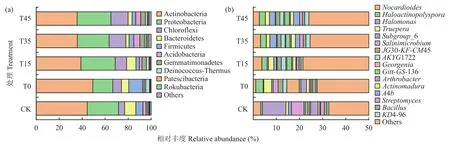
图3 不同处理土壤样品细菌在门(a)、属(b)水平上的物种分布组成Fig. 3 Composition of bacteria at the phylum (a) and genus (b) levels in the soil under different treatments
5个处理土壤中诺卡氏菌属(Nocardioides)和晕轮放线菌属(Haloactinopolyspora)占主导地位。T0和T45处理较CK显著降低了诺卡氏菌属(Nocardioides)的相对丰度,其他处理与CK无显著差异;T0、T15、T35和T45处理较CK提高了晕轮放线菌属(Haloactinopolyspora)和特吕珀菌属(Truepera)的相对丰度,但显著降低了盐单胞菌(Halomonas)和新疆盐坑微菌(Salinimicrobium)的相对丰度。
由图4所示,在UPGMA聚类树上,施用柠条堆肥的4个处理(T0、T15、T35与T45)为一个大类,施用鸡粪的处理(CK)为另一个大类,表明有机肥种类是造成土壤细菌群落结构差异的主要因素;柠条堆肥处理中,免耕(T0)与其他3个耕作处理土壤细菌群落结构分属不同亚类,耕作制度也是影响细菌群落结构的重要因素;翻耕深度的增加改变了菌群结构,T45和T35处理分为一个小类,表明细菌群落关系最相近,其次与T15处理为一个亚类,三者细菌群落较为相近;从聚类分析图中发现,不同处理菌群与CK的距离随着耕作深度的增加而增加,说明施肥类型和耕作深度交互作用造成细菌群落结构差异的增加。

图4 不同处理UPGMA聚类分析图Fig. 4 Graphics of UPGMA clustering analysis in different treatments
2.4 不同处理对微生物功能多样性的影响
图5显示了细菌丰度前19位的KEGG基因代谢通路条目。这19条代谢通路按照功能可划分为代谢、遗传信息处理、环境信息处理和细胞进程4大类。代谢功能与土壤物质循环密切相关,其中氨基酸代谢、辅助因子和维生素代谢、萜类和聚酮化合物代谢、其他氨基酸代谢、类脂化合物代谢相关的微生物相对丰度大小顺序均为T35>T45>T15>T0>CK。

图5 不同处理土壤细菌代谢通路Fig. 5 Bacterial metabolic pathways in the soil under different treatments
细菌碳代谢功能对土壤物质循环和作物生长至关重要,细菌群落对6大碳源的利用程度如图6所示。6大碳源中,土壤微生物对多聚物利用程度最高,对羧酸类化合物利用程度次之,对碳水化合物的利用程度最低。虽然不同碳源的利用程度受耕作深度的影响不同,柠条堆肥各处理对6种碳源的利用总的看来均显著高于鸡粪处理(CK)。免耕处理(T0)对氨基酸、胺类化合物和芳香类化合物的利用率均高于翻耕处理,而翻耕处理对羧酸类化合物和多聚物的利用程度高于免耕。

图6 不同处理下6大碳源的利用情况Fig. 6 The utilization of six carbon sources under different treatments
2.5 环境因子、细菌群落和功能、微生物碳代谢的冗余分析
施肥和耕作主要通过改变环境因子影响土壤微生物。环境因子、细菌群落、细菌功能和微生物碳代谢间的冗余分析(RDA分析)结果(图7)显示,pH、EC和速效氮对细菌群落结构和微生物群落碳代谢功能影响显著(P<0.01),丰度排在前9名的细菌也与土壤碳代谢紧密相关,前两轴解释的环境因子、细菌群落、细菌功能和微生物碳代谢总变异分别为58.24%、58.266%、43.671%和80.76%。绿弯菌门、酸杆菌门、芽单胞杆菌门、羧酸类化合物代谢和萜类化合物和聚酮化合物代谢与土壤pH呈显著正相关,酸杆菌门、芽单胞杆菌门、其他氨基酸代谢、异生物素生物降解和代谢和能量代谢与土壤碳氮比呈正相关,EC值和速效氮含量有利于厚壁菌门丰度和氨基酸代谢的提升,绿弯菌门和芽单胞杆菌门与羧酸类化合物代谢呈显著正相关,厚壁菌门和放线菌门与胺类化合物和芳香类化合物呈显著正相关。
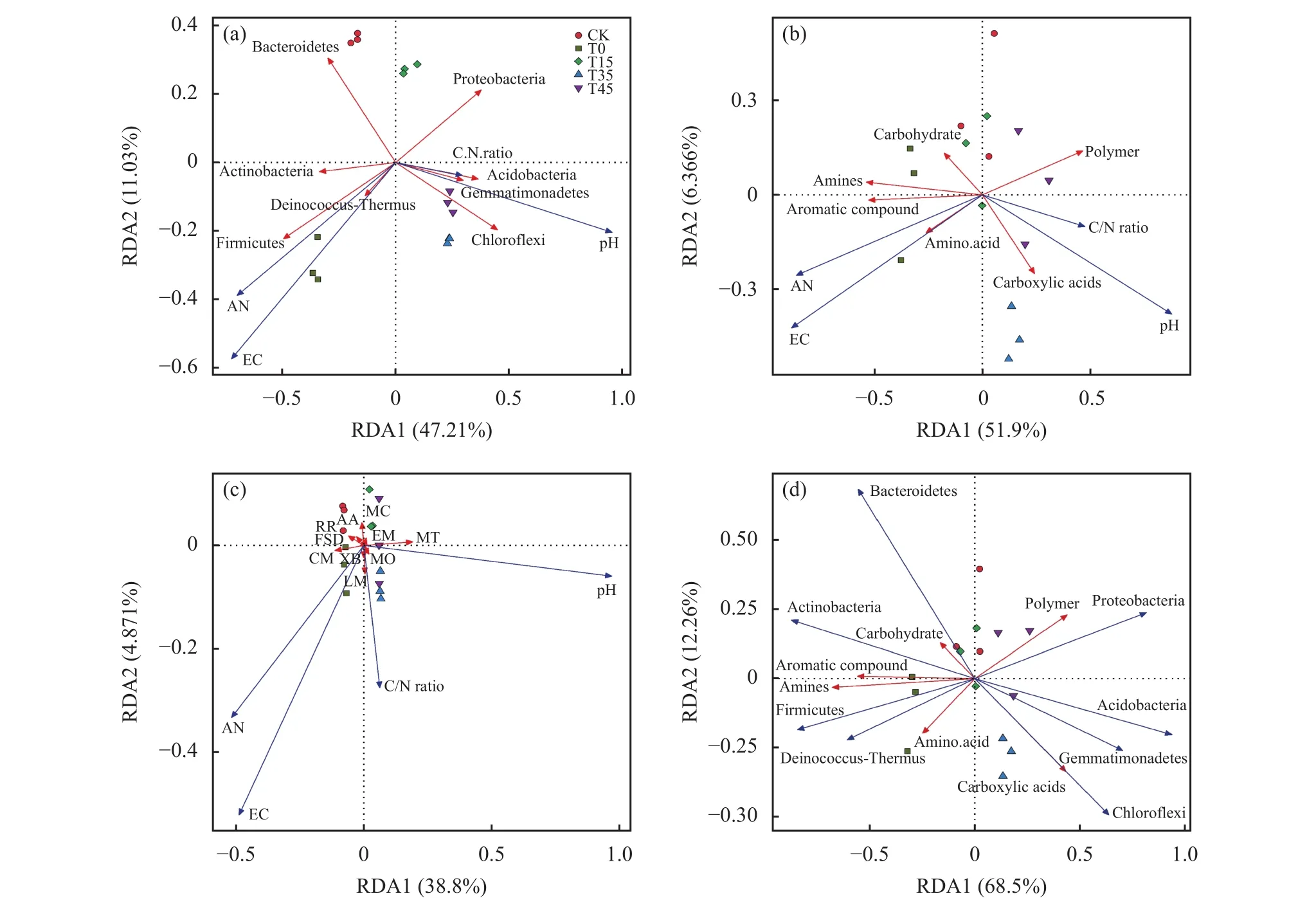
图7 环境因子(a)、细菌群落组成(b)、细菌功能(c)及微生物碳代谢(d)之间的冗余分析Fig. 7 Redundancy analysis among environmental factors, bacterial community composition, bacterial function,and microbial carbon metabolism
3 讨论
本研究发现与传统鸡粪相比,柠条堆肥的施用并未显著降低黄瓜产量。粪肥的长期施用会造成土壤酸化,从而影响土壤养分循环及作物生长[9]。本研究发现柠条堆肥连续施用较传统鸡粪提高了土壤pH,这是因为柠条堆肥施用后向土壤中输入了更多中碱性物质,提升了土壤的缓冲能力[10]。同时,结合深翻可调节土壤耕层结构和土壤碳氮比,降低氮循环效应,从而促进有机肥分解消耗质子来抑制土壤酸化[11–12]。耕作深度增加降低了速效氮的含量,可减少表层土壤养分的聚集,降低土壤次生盐渍化的风险。
微生物群落多样性是表征农田土壤微生物群落功能的发挥和土壤特征及功能的重要指标[13]。本研究发现在相同翻耕深度下柠条堆肥施用较传统鸡粪提高了土壤细菌群落的多样性,说明柠条堆肥含有更丰富的营养物质,促进了土壤微生物的生长。路颖等[14]研究也发现,性质、质量和分解速率差异的不同类型有机物输入到土壤中,会对土壤微生物群落多样性产生显著影响。有研究表明耕作方式也是影响微生物多样性的重要因素[15]。本研究发现柠条堆肥各处理中,耕作处理(T15、T35和T45)较免耕处理(T0)均提高了土壤细菌丰富度、多样性和均匀度指数,T35处理最高。
本研究发现各处理样品中放线菌门、变形菌门、绿弯菌门为土壤细菌群落的优势菌门,这与贾远航等[16]研究的结果一致,这说明不同土壤的微生物群落存在很多共性和一定的环境适应性。尽管如此,施肥和耕作对土壤微生物群落结构组成造成了一定的影响,肥料类型为主导因素。本研究中,放线菌门的相对丰度最大,被视为土壤碳氮循环主要功能细菌[17]。有研究表明,放线菌门的相对丰度与土壤团粒结构的大小呈负相关[18]。相同耕作深度下,柠条堆肥处理放线菌门的相对丰度低于鸡粪处理,这可能与土壤结构的变化有关。柠条堆肥为土壤微生物的物质循环提供了原料、能源和良好的生存环境,产生的有机无机胶结物质促进了团粒结构的形成,从而降低了放线菌门的相对丰度。同时,在柠条堆肥各处理中,免耕处理(T0)放线菌门、厚壁菌门的相对丰度高于耕作处理(T15、T35和T45),这可能是放线菌门和厚壁菌门作为富营养菌在有机营养输入条件下更多地依赖于不稳定的碳源[19],免耕处理为放线菌门和厚壁菌门提供了更为充足的营养,其他菌群受外源输入养分影响较小。耕作深度的增加改善了土壤耕层结构,促进了变形菌门、绿弯菌门、酸杆菌门和芽单胞杆菌门的相对丰度的提升。
本研究对细菌功能预测发现,代谢为细菌群落的主要功能,柠条堆肥各处理主要代谢通路基因丰度均高于CK,T35处理对氨基酸、碳水化合物等物质的合成促进效果最好。细菌碳代谢功能对土壤物质循环和作物生长至关重要[20]。本研究中柠条堆肥各处理碳源利用程度均高于CK,这可能是柠条堆肥作为豆科植物残茬堆肥富含有机质,为微生物的生长提供了充足的有效碳源。耕作深度也显著影响了碳源的利用,微生物对氨基酸、胺类化合物、碳水化合物和芳香类化合物的利用程度随着耕作深度的增加而降低,而深翻处理提高了羧酸类化合物和多聚物的利用。
施肥和耕作分别通过直接和间接方式影响微生物群落[21];一方面可以直接为微生物创造适宜的生存环境,另一方面通过调控环境因子改变微生物群落。有研究表明,不同处理间细菌群落组成差异主要取决于土壤性质[22]。本研究发现土壤pH、EC和速效氮含量对土壤细菌群落组成、功能和碳源利用均有显著的影响,其中pH是影响土壤群落结构的主要驱动因子[23–24]。绿弯菌门、酸杆菌门、芽单胞杆菌门与土壤pH呈显著正相关,柠条堆肥施用和耕作深度增加通过提高土壤pH来促进这些菌群丰度的增加。细菌群落的变化和生物功能紧密相关[25–26]。本研究发现,前9丰度的细菌显著影响了土壤碳代谢,绿弯菌门和芽单胞杆菌门与羧酸类化合物代谢呈显著正相关,厚壁菌门和放线菌门与胺类化合物和芳香类化合物呈显著正相关。变形菌门和厚壁菌门作为富营养菌,可能会受到EC和速效氮等养分指标的调控,从而影响相关功能的表达,此外,EC值过高对大多数菌群结构和功能发挥呈负向影响。
4 结论
柠条堆肥对黄瓜的增产效果与鸡粪没有显著差异。柠条堆肥配合免耕可提升土壤EC值和速效氮含量,配合深翻(35 cm、45 cm)可显著提高土壤碳氮比和土壤细菌群落多样性。放线菌门、变形菌门、绿弯菌门为土壤细菌群落的优势菌门,柠条堆肥能提高土壤优势菌门相对丰度,T35和T45能够显著提高变形菌门和绿弯菌门相对丰度。同时,柠条堆肥各处理也有效提高了主要代谢通路相对丰度,有利于菌群功能发挥;相对鸡粪,柠条堆肥有助于细菌碳代谢能力的提升,结合深翻进一步提高了对羧酸类化合物和多聚物的利用。RDA分析表明,环境因子、细菌群落和菌群功能间主要处在共生关系,pH是影响土壤群落结构的主要驱动因子,可通过柠条堆肥和适当深翻(T35)来维持良好土壤环境,作为调节土壤微生物群落、维持农田生态可持续发展的重要方式;同时,可通过优化有机肥使用量及有机无机配比研究进一步提高产量。
猜你喜欢
科学技术与工程(2022年26期)2022-11-01
现代园艺(2022年19期)2022-09-21
中国农学通报(2022年14期)2022-06-01
油气田环境保护(2022年2期)2022-05-09
内蒙古林业(2021年7期)2021-12-31
现代农村科技(2021年5期)2021-12-07
小读者(2021年19期)2021-11-07
乡村科技(2021年16期)2021-10-21
世界竹藤通讯(2020年5期)2020-11-11
当代陕西(2019年7期)2019-04-25
