
油棕的花序和花组织形态解剖结构观察
2022-04-11 05:53张大鹏李东霞
西南林业大学学报 2022年2期
石 鹏 王 永 张大鹏 李东霞
(中国热带农业科学院椰子研究所/国家热带棕榈种质资源圃/农业农村部热带油料科学观测实验站/海南省热带油料作物生物学重点实验室,海南 文昌 571339)
花是被子植物的生殖器官,被子植物的花一般含有花柄、花托、花萼、花瓣、雄蕊和雌蕊。花的正常发育对于植物繁殖后代至关重要,解剖结构观察可以看到植物器官微观结构,花解剖结构观察对于理解花发育和功能具有重要意义。国外对棕榈科和油棕的花序和花起始过程研究得比较透彻。棕榈科包含大约2 400个物种[1],其花序大多从叶腋基部着生。油棕只有1个茎尖生长点,成熟油棕树大约每2周产生1个新的叶原基,叶片从起始到脱落大约需要2~3年[2],而花序则在叶腋中不断形成。作为典型的单子叶植物,棕榈科植物通常为三基花,萼片和花瓣分开[3]。油棕花发育过程复杂,其中雄花序发育过程中只具有单独的雄花,而雌花序早期则产生有一朵雌花和两朵雄花组成的两性花,后期两朵雄花败育形成只有一朵雌花的单性花[4]。油棕(Elaeis guineensis)是棕榈科油棕属的多年生乔木,油棕的果实含油率可达50%,是世界上最重要的热带木本油料作物[5]。然而,成熟油棕花序和雌雄花的形态和解剖结构还不清楚。因此,开展油棕雌雄花解剖结构观察有助于了解其生殖器官发育过程,对于授粉后果实和种子形成过程的认识具有重要意义。近年来国内对油棕花的研究主要集中在花序性别分化[6]、花粉活力和柱头可授性的测定[7]、花粉形态观察[8]等方面。针对油棕花器官的发育,杨静等[6]对油棕花序性别分化过程进行了较为细致的观察,他们发现油棕雌、雄花序的发育过程有所不同:雌花存在1个两性期阶段,由1个有功能的雌花和两侧的伴随雄花组合而成,伴随雄花选择性败育后,雌花即成为单性花;而雄花不会经历两性期阶段。李元元等[7]发现不同果壳类型油棕花粉活力和柱头可授性存在差异。李元元等[8]发现不同果壳类型油棕花粉显微结构均为近正三角形,都有3条萌发孔沟。张大鹏等[9]发现冷冻切片适用于油棕幼嫩叶片的快速显微观察。石蜡切片是组织学、发育生物学研究的主要实验方法,利用石蜡可与植物组织很好的结合的原理,制成石蜡切片,在显微镜下观察,能清晰地反映机体的结构和形态变化[10]。取油棕雌雄花序,对雌、雄花外部形态进行观测,结合冷冻和石蜡切片技术观察雌、雄花的显微结构。通过观察雌雄蕊在形态上与解剖结构上的特征,了解生殖发育基础,为进一步研究油棕种子形成过程奠定基础。
1 材料与方法
1.1 试验材料
本试验于2015年11月取7年树龄油棕树的雌雄花,油棕树位于中国热带农业科学院椰子研究所的国家热带棕榈作物种质圃(油棕分区)(北纬19°31′40″~19°31′58″,东经110°45′54″~110°46′4″),地块位于海南省文昌市,土壤为滨海沙地(表土pH值4.4~4.8,有机质含量约1.5%,全氮含量约0.3 g/kg,有效磷含量约0.05 g/kg,速效钾含量约0.04 g/kg),气候为热带季风岛屿型气候(月平均气温15.6~32.8 ℃,月平均降水量33.6~352.1 mm)。
1.2 试验方法
1.2.1树和花序观察
选取有雌雄花开放的油棕树和雌雄花序,利用Nikon D7100照相机拍照。择取处于开花状态中的雌雄花,分别放在Leica M50体视显微镜下,适当调节焦距观察雌雄花外部形态,拍下清晰的图片。
1.2.2冷冻切片观察
将新鲜取回的雌雄花用于冷冻切片,在样品托上倒少量的冰冻包埋剂OCT,放在提前预冷冷冻切片机凝固后形成一个衬垫,把需要观察的样品放在衬垫上,继续用包埋剂浇注样品,直到样品被完全包裹住,迅速放入Leica CM1900冷冻切片机后固化。固化后,用预冷的刀片修块,再切片,厚度为10~16 μm,取片后观察或染色后在Leica DM 2500倒置生物显微镜下观察并拍照,具体方法参考文献[9, 11]。
1.2.3石蜡切片观察
首先选取还未完全开放的雌花,将萼片剥掉,之后将雌蕊全部剥出,随后马上用解剖刀把柱头和子房分开,立刻放入装有卡诺固定液(V(无水乙醇)∶V(乙酸)=3∶1)的高型称量瓶,真空抽气固定后,经95%乙醇、85%乙醇,每级20~25 min,之后转入70%乙醇中备用。历经整染、水洗与返蓝、脱水、透明、加碎蜡、透蜡、包埋、修蜡块与切片、展片、粘片及烘干、脱蜡与透明、封片、烘片、观片等环节。切片厚度为14~16 μm,在Leica DM 2500倒置生物显微镜下观察并拍照,具体方法参考文献[11-12]。
2 结果及分析
2.1 花序和花的器官形态结构
油棕花雌雄异序同株,每个叶腋内可着生花序,即花序腋生。雌雄花序均为肉穗花序,花苞未开裂时,雌雄花序形态相似,称为佛焰苞。近头状的为雌花序(图1a,1b),而多个指状的小穗则构成了雄花序(图1a,1g)。
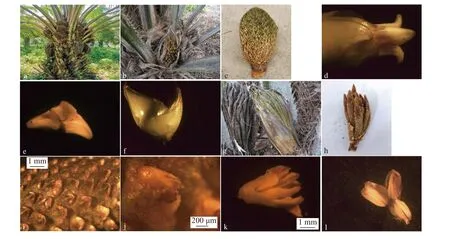
图 1 油棕花序与花形态结构Fig. 1 Inflorescence and flower structure of E. guineensis
雌花序长度30 cm左右,上可着生近1 000朵雌花,每个小穗上能着生5~40朵雌花,许多小花在小穗轴上螺旋排列(图1b,1c)。成熟雌花序的花序柄长20~30 cm。雌花由2个苞片包裹,雌花长约2~3 cm。苞片内含6片卵形或卵状长圆形花瓣和1枚雌蕊,花瓣长约2 cm,宽约1 cm。苞片和花瓣均呈淡黄色,花瓣呈覆瓦状排列(图1d,1f)。油棕雌蕊由3个心皮组成(图1d,1e),花柱较短,雌蕊的基部膨大部位是子房。
雄花序的每一个指状花穗长为10~20 cm,其直径为0.8~1.5 cm,雄花着生于小穗上,每个小穗有近1 000个雄花。穗轴顶端呈突出的尖头状,成熟雄花序轴下的花序柄长为30~40 cm(图1h)。雄花由3片苞片包裹,内含6枚雄蕊,雄花长约3~4 mm,宽1.5~2.0 mm。成熟的雄花有3片花萼3片花瓣,覆瓦状排列。萼片革质,呈船状,雄花的萼片长约4 mm、宽约1 mm,苞片的顶端稍尖,有棱(图1j,1k,1l);雄花的形态结构与雌花相仿,每个雄花有管状雄蕊6枚,雄蕊由花药和花丝组成(图1l)。
2.2 油棕雌蕊的形态及内部结构
油棕雌蕊的柱头表面3裂(图2a,2b),每个柱头裂片的中央皆有一个由乳突细胞组成的带状柱头沟,授粉前裂痕会逐步增大,授粉后又会逐步合拢(图2b,2c)。这一生理特性大大增加了授粉的成功率。柱头表面会形成不同大小的乳突,其分泌的物质有利于粘落在柱头表面的花粉萌发,由此推断柱头类型为湿型柱头(图2c)。花柱基部中间空腔周围有染色较深的圆点为花柱道,是花粉萌发后进入胚珠的通道,属于开放型花柱(图2d)。花柱连通柱头和子房,在花柱底部稍微膨大的部分为子房(图2c,2d)。子房中3心皮3室,每个子房室中只有1个胚珠,胚珠长约200 μm,宽约100 μm(图2d,2e)。一般只有1个胚珠能发育成种子,其他2个败育。
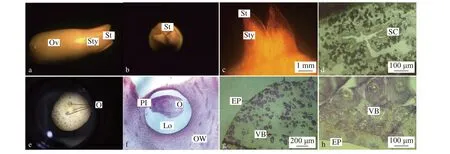
图 2 油棕雌蕊形态结构Fig. 2 Pistil structure of E. guineensis
花柄是植株养分进入花蕊的唯一通道,在观察花柄横切片中,发现油棕雌花的花柄主要由表皮和维管束组成(图2f),与植物茎的组成结构相似。维管束遍布花柄,相间排列,各维管束之间相隔3~4层薄壁细胞(图2g),与雄花花柄维管束结构相似。
2.3 油棕雄蕊的形态及内部结构
花药和花丝是油棕雄蕊的组成部分,花丝顶端呈长囊状的结构为花药,花药长约3 mm,宽约0.5 mm(图3a,3b),花药内含大量花粉粒。在显微镜下观察发现,每个成熟花药有4个椭圆形的花粉囊,淡黄色,左右对称分开,花粉囊之间以药隔相连,花药横切面近似蝶形(图3c)。花粉囊直径约100 μm,花粉粒直径约15 μm。花药的壁主要由表皮、纤维层和绒毡层等细胞组成,表皮偏薄(约5 μm),纤维层偏厚(约10 μm),绒毡层细胞致密且颜色较深(图3c,3d)。药室内有大量花粉,都以单粒的形式存在,其形状近似三角形或椭圆形(图3c,3d)。在花粉中可观察到极少量形态较小或其他形状的花粉,可能是发育不良的花粉。

图 3 油棕雄蕊形态结构Fig. 3 Stamen structure of E. guineensis
雄花指状小穗中间为小穗轴(直径约2 mm),周边着生雄花(图3e)。花柄外侧呈椭圆形的细胞排列紧密,为韧皮部,中间细胞排列疏松,为木质部,另外还散布大量圆形气腔(图3f)。维管束(直径约100 μm)呈椭圆形,较大的空腔为导管,较小的空腔为筛管,导管将水分和无机盐运输到花中,使其得到充足的水分和养料(图3g)。花柄维管束中的纤维增加了细胞群和组织之间的支撑力,发达的木纤维增强了花柄的强度,使花朵得到充分的支撑,不易脱落(图3g,3h)。
3 结论与讨论
油棕雌雄同株异序,花序腋生,肉穗花序(佛焰花序),雌雄花序均由2层苞片包裹,花序由众多小穗组成。雌花有6片花瓣,内含1枚雌蕊,湿型柱头,开放型花柱,三心皮三室,每个子房室中有1枚胚珠,花柄中散布大量维管束。雄花有3片萼片3片花瓣,内含6枚雄蕊,每个花药含4个花粉囊,中间被药隔相分,花粉形态近似三角形或椭圆形,花柄主要包含表皮、基本组织和维管束,其中遍布大量维管束和纤维。油棕在雌雄花序性别上与椰子、槟榔和椰枣等棕榈科植物存在差异,油棕雌雄同株异序,而椰子和槟榔雌雄同株同序,花序上部为雄花,下部为雌花[13-14],椰枣则为雌雄异株。这些差异是物种长期进化的结果,也是初步判断物种类别的表型证据[15]。
在李元元等的报道中,油棕花粉外形呈近正三角形[8],本研究观察到的花粉形态有近三角形、椭圆形的正常花粉,也有因发育不良的个体较小花粉,发育不良的花粉可能会引起花粉败育。花粉粒由外壁、内壁、营养细胞和生殖细胞构成,淀粉粒、线粒体在花粉发育的过程中,提供了营养和能量,如果这些细胞器和结构发育不全,可能造成花粉不能正常萌发。徐丽萍等[16]和陆彦等[17]都在花粉中观察到了这些细胞器和结构。李东霄等[18]发现小麦中淀粉粒的积累与花粉育性相关。上述相关的研究结果印证了植物界中存在的共性,可以推断,油棕花粉粒可能也存在上述相关特征。以后可进一步观察发育不良的花粉粒内部结构,找到导致油棕花粉败育的原因。
油棕花柄中发达的维管组织,构成了发达的纵向养分运输系统,保证了植株的水分和营养能及时向花部供应。胡露洁等[19]发现濒危水生植物莼菜的花柄维管组织结构与根茎类似,油棕可能也类似。张颖等[20]在观察锦绣杜鹃花柄时指出,花柄在开花期会积累大量的淀粉粒,以满足子房发育膨大的需要,随着子房逐渐膨大,花柄中的淀粉粒也会逐渐向子房转移。因此,可以根据淀粉粒的在花柄中的量初步推断子房的发育状况,但在油棕的花柄中是否也有类似特征,需要进一步研究。另外,赵庆芳等[21]发现5种棕榈科植物花序轴解剖结构存在种间差异,可以作为棕榈科植物分类的辅助证据。油棕雌雄花柄解剖结构是否可以作为区分椰子、槟榔和椰枣等其他棕榈科植物的证据,还需要进一步研究。
综上所述,油棕花序和花的形态特征符合棕榈科植物的基本特征,为肉穗花序,花单性,三基花,每个子房1个胚珠,但在花序性别和花解剖结构上与椰子、槟榔和椰枣等棕榈科植物存在差异,可以作为区分其他棕榈科植物的特征。雌雄花柄主要包含表皮、基本组织和维管束,大量维管束为花的正常发育提供充足养分。本研究还不能解释花粉败育、无籽果实等油棕生殖器官异常现象的生物学机制,宜采用亚显微技术和高通量多组学技术作进一步的深入研究。
猜你喜欢
安徽农业科学(2022年18期)2022-10-13
金沙江文艺(2022年4期)2022-04-26
江苏农业科学(2020年21期)2020-03-08
湖北农业科学(2019年20期)2019-12-11
中国瓜菜(2019年8期)2019-09-19
儿童故事画报·自然探秘(2019年10期)2019-01-14
安徽农业科学(2018年27期)2018-05-14
现代农业研究(2017年10期)2018-01-02
Coco薇(2015年12期)2015-12-10
农村百事通(2009年8期)2009-05-25
