
基于SSR分子标记的早熟棉种质资源遗传多样性分析
2022-04-09 03:04路战远张建中陈立宇程玉臣杨建强张向前
北方农业学报 2022年1期
咸 丰,路战远,张建中,陈立宇,程玉臣,杨建强,张向前,苏 和
(1.内蒙古自治区农牧业科学院,内蒙古呼和浩特 010031;2.阿拉善盟农业技术推广中心,内蒙古巴彦浩特 750306)
棉花是世界上重要的经济作物之一,不仅是纺织业主要的天然纤维原料作物,也是动物饲用蛋白以及人类食用油的重要资源。我国是棉花生产大国,也是消费大国,但自给率仅为70%左右,进口率仍高达30%,极大地制约着我国棉花产业的可持续发展[1]。随着我国棉花新品种培育速度的加快以及骨干亲本的集中使用和转基因技术的应用,棉花品种间的遗传差异越来越小[2-4]。因此,如何充分利用现有棉花种质资源进行育种,实现育种目标性状有突破性进展已迫在眉睫。
分子标记是基于DNA 多态性的遗传标记,可以直接反映个体在DNA 水平上的差异,具有许多其他标记无法比拟的优势[5-6],其中,SSR分子标记(又称微卫星DNA),是一类由几个核苷酸为重复单位组成的串联重复序列[7],具有共显性、多态性强、可复制性强、基因组覆盖范围广等特点[8]。SSR 标记作为一种理想的分子标记,在棉花种质资源多样性评价和遗传研究中也有较多应用,BOURGOU 等[9]利用SSR分子标记分析了当地传统棉花种质的遗传多样性,并与现有品种的多样性进行了比较。贾子昉等[10]利用SSR分子标记分析了300 份棉花种质资源的遗传多样性,为进一步开展育种工作提供了理论依据。石建斌等[11]利用SSR分子标记对58 份棉花种质资源进行遗传多样性分析,结果表明,这些资源聚类与其地理生态来源无关,而与材料的亲缘关系相关性较高。卫泽等[12]通过表型聚类和SSR分子标记聚类研究了国内外57 份棉花种质的遗传多样性,各聚类结果表明,同一国家的品种遗传差异较小,国家间品种差异较大,但也存在明显的相互渗透,多数品种没有表现出明显的地域差异。高伟等[13]对60 份四倍体棉种材料进行遗传多样性分析,结果显示,从1 050对SSR 引物筛选出的95 对SSR 引物均能在60 份材料间扩增出稳定明显的多态性条带,共检测出660 个片段,其中多态性片段584 个,占88.5%,每个位点的等位基因为2~12 个,平均每对引物6.1 个。
鉴于内蒙古自治区农牧业科学院棉花研究中心早熟棉种质资源来源广泛,资源丰富,为弄清现有早熟棉种质资源的亲缘关系,本试验利用SSR分子标记对44 份早熟棉种质资源进行遗传多样性分析,以期为棉花种质资源鉴定、育种亲本选择、新品种改良培育、重要基因挖掘等工作奠定基础。
1 材料和方法
1.1 试验材料
供试的44 份早熟棉种质资源,均由内蒙古自治区农牧业科学院棉花研究中心收集、保存和种植(表1)。
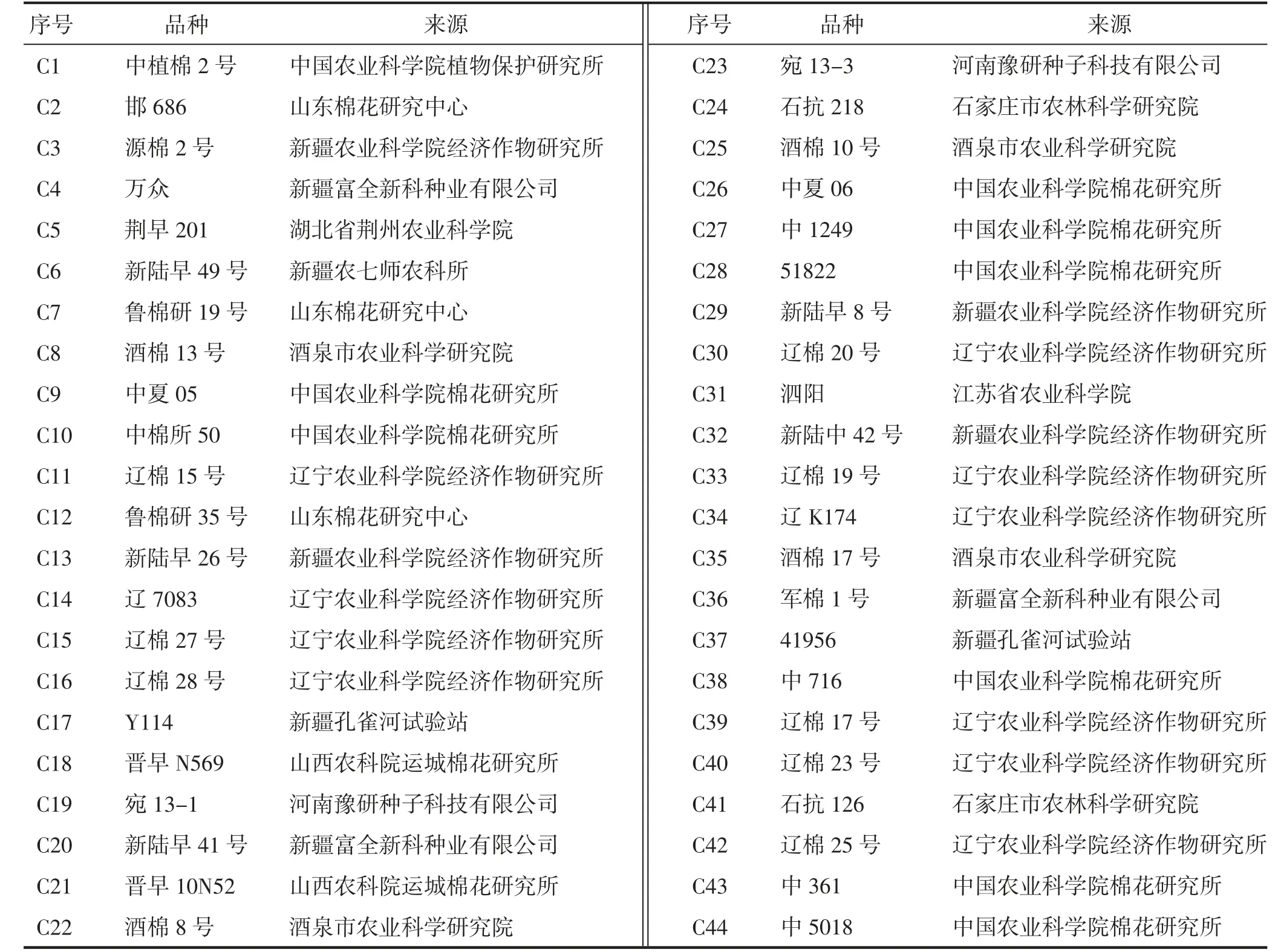
表1 供试早熟棉种质资源的编号、名称及来源
1.2 试验方法
1.2.1 棉花基因组DNA 的提取
试验于2019年春季在棉花研究中心实验室进行,每个品种育苗10 株,待2~3 片真叶时,每份材料随机选择新鲜幼嫩叶片4~5 片,液氮速冻后置于-80 ℃冰箱保存备用。采用改良的CTAB 法提取棉花叶片基因组DNA[14],用紫外分光核酸测定仪检测DNA 浓度及质量,将DNA 稀释到80 ng/μL,于-20 ℃保存备用。
1.2.2 SSR 引物多态性扩增
所用SSR 引物序列见表2,参考张玉翠等[15]、潘兆娥等[16]的试验设计,由上海生物工程有限公司合成。PCR反应体系:2×Taq Master Mix(康为世纪)10 μL,DNA 模板1 μL,10 mmol/L 引物各0.5 μL,ddH2O 补至20 μL。PCR反应程序为:94 ℃预变性5 min;94 ℃变性30 s,48~54 ℃退火30 s,72 ℃延伸90 s,共35 个循环;72 ℃延伸10 min,4 ℃保存。采用6%非变性聚丙烯酰胺凝胶电泳分离检测。

表2 SSR 引物及其序列
1.2.3 数据处理
数据记录时,只统计3 次重复试验中稳定出现的条带,将任一扩增条带作为一个等位位点,按条带的有/无分别赋值1/0,建立二元数据矩阵。利用NTSYS-pc V2.10 软件中的Qualitative date 函数计算品种间的遗传相似系数(Jaccard 系数);利用非加权成对群算术平均数法(unweight pair group method using arithmetic averages,UPGMA)进行聚类分析;最后,采用Treeplot 模块生成树状聚类图。
2 结果与分析
2.1 SSR 引物多态性分析
本试验用筛选出的31 对条带清晰且扩增多态性较好的引物,对44 份早熟棉种质资源进行遗传多样性分析,在目标群体中检测出72 条多态性条带,每对引物扩增1~6 条,平均能够扩增出2.25 条。其中,NAU1200、CIR216、BNL3140、NAU859、DPL0442、NAU5099,6 对SSR 引物仅检测到1 个等位基因,而MUCS101 和NAU3110 检测到6 个等位基因(图1)。

图1 NAU1200(a)、CIR216(b)、MUCS101(c)和NAU3110(d)引物对44 份早熟棉材料扩增的电泳谱
2.2 早熟棉亲缘关系的聚类分析
依据UPGMA 法对供试的44 份早熟棉种质资源进行遗传多样性聚类分析(图2),供试44 份早熟棉种质资源间的遗传相似系数为0.35~0.80,平均为0.58。其中,C35(酒棉17 号)与C38(中716)相似系数最小,仅为0.35;相似系数较小的还有C43(中361)与C31(泗阳)、C3(源棉2 号)与C40(辽棉23 号),分别为0.37、0.38,C43(中361)与C30(辽棉20 号)、C14(辽7083)与C25(酒棉10 号)均为0.39;C13(新陆早26 号)与C40(辽棉23 号)、C42(辽棉25 号)与C28(51822)、C42(辽棉75 号)与C30(辽棉20 号)均为0.40,表明材料间的遗传差异较大,亲缘关系较远。相反,C20(新陆早41 号)与C43(中361)、C9(中夏05)与C11(辽棉15 号)的相似系数最大,均为0.80,表明材料间的遗传相似性较高,基因组间差异较小,亲缘关系较密切。此外,追溯各材料的来源及其遗传背景,发现来源于河南豫研种子科技有限公司的C19(宛13-1)和C23(宛13-3)也表现出较近的亲缘关系,表明选育背景具有较大相似性。聚类结果相对较客观地反映了44 份材料间的亲缘关系。
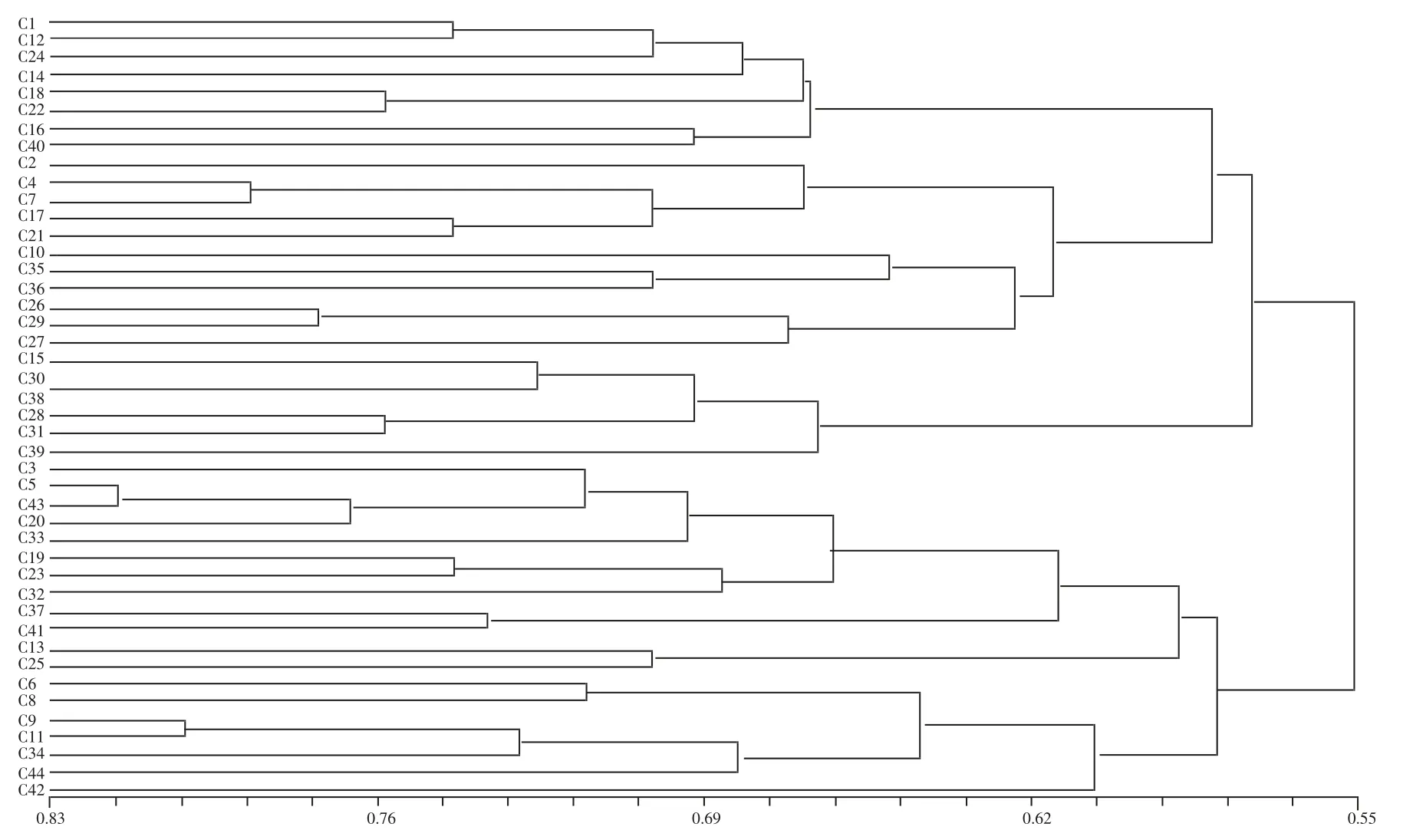
图2 早熟棉种质资源的UPGMA 聚类结果
3 讨论与结论
对现有早熟棉材料的遗传多样性进行聚类分析,明确种质资源的遗传基础和分类,有助于棉花的育种研究。同时运用分子生物学的方法从基因水平上分析种质资源的多样性,可以充分挖掘棉花种质资源的丰富基因。潘兆娥等[16]通过构建棉花参比种质,高效快速地筛选出319 对核心SSR 引物,为棉花品种间遗传多样性的评价奠定了基础。贺道华等[17]利用分布于棉花全基因组的132 个SSR 标记对精选的92 个棉花品种(系)进行聚类分析,结果显示,92 个品种之间的成对相似系数平均为0.729 2±0.135 0。董章辉等[18]利用22 对SSR 多态性引物共扩增出30 条多态性条带,遗传多样性分析表明,83 份早熟抗虫棉材料的相似系数为0.73~0.98。陈浩东等[19]利用68 对SSR引物,对15 份短季棉材料进行了聚类分析,共检测出178 个有效等位位点,平均每对引物检测到2.62 个,各材料基因型间遗传相似系数为0.45~0.79。王欣怡等[20]利用78 对SSR 多态性引物在120 个新疆陆地棉材料中检测到392 个等位位点,其中多态性位点324 个,多态性比例达82.7%;聚类分析显示,120 个陆地棉品种遗传相似系数变化范围为0.50~0.96,平均为0.73。翟书伟等[21]利用39 对SSR 多态性引物在26 份棉花材料中检测到99 个多态性位点,遗传相似系数为0.73 时,可聚为4 类。本试验利用筛选的31 对SSR 引物对早熟棉品种进行聚类分析,结果表明,44 份早熟棉品种间的遗传相似系数分布在0.35~0.80,平均为0.58,与前人相比遗传相似系数较低,也表明我国早熟棉遗传多样性高,遗传背景差异较大,遗传基础相对较广,但部分材料基本接近,可能是由于育种研究者在选育新种质的工作中育种目标过于单一,一些棉花遗传背景多样性逐渐丧失[22-23]所致。
综上所述,本试验利用31 对SSR 引物在44 份早熟棉品种中检测出72 条多态性条带,平均每对引物检测到2.25 条。聚类分析表明,被测材料间的遗传相似系数为0.35~0.80,平均为0.58,初步揭示了现有44 份早熟棉资源间的遗传背景差异及其相互亲缘关系,为下一步合理高效选择杂交亲本以及进行杂交选育等提供了理论依据。
猜你喜欢
今日农业(2022年14期)2022-09-15
今日农业(2022年13期)2022-09-15
计算机应用与软件(2022年6期)2022-07-12
自然灾害学报(2022年2期)2022-05-10
昆明医科大学学报(2022年3期)2022-04-19
昆明医科大学学报(2022年2期)2022-03-29
中国种业(2022年1期)2022-01-27
中国种业(2022年1期)2022-01-27
昆明医科大学学报(2021年5期)2021-07-22
中国学校体育(2021年10期)2021-04-26
