
金银花U6启动子的克隆及转录活性分析
2022-04-08 05:44唐志强李小丽许小涵蒲高斌
山东科学 2022年2期
唐志强,李小丽,许小涵,蒲高斌,2*
(1.山东中医药大学,山东 济南 250355;2.山东省中药质量控制与全产业链建设协同创新中心,山东 济南 250355)
金银花为忍冬科植物忍冬LonicerajaponicaThunb.的干燥花蕾或带初开的花,有清热解毒、疏散风热等功效。目前,中国新冠肺炎疫情虽然整体得到了有效的控制,但是国内个别地区仍有反复,国际形势依然非常严峻。金银花为清热类药物,通过发挥清热解毒、疏风散热等功效,起到防御病毒的作用,成为各个省份使用频率较高的中药之一,社会需求也随之不断加大[1]。病虫害是导致金银花产量不高、质量不稳的主要原因,因此急需培育出优良种质来提高金银花的产量、质量和抗病虫害能力,满足社会需求。
基因编辑是改良品种、创新种质的新方法,已在多个物种中成功应用。CRISPR/Cas9基因组编辑技术因为其高效简便等优点,已经成为目前功能基因研究及种质改良的主要工具。在水稻中过表达OsPPDK基因,能够在种子形成时期提高叶片的光合效率,促进衰老叶片的N活化,在延缓叶片衰老的同时提升水稻的持绿性,为增加水稻产量打下了基础[2]。Wang等[3]利用CRISPR/Cas9技术生成携带TaMLO-A1等位基因突变的转基因小麦植株,提高抗白粉病能力的同时增加了小麦的产量。Do等[4]通过编辑大豆FAD2-1A和FAD2-1B基因,将油酸含量增加了80%,亚油酸含量下降到1.3%~1.7%,改善了种子油脂的性状,创造出高油酸大豆。U6RNA是一种小的非编码RNA,广泛参与mRNA前体的剪接成熟[5]。U6RNA所对应的U6启动子通常被用来驱动sgRNA的转录,是CRISPR/Cas9系统中的重要组成部分。目前,U6启动子已在多种植物的CRISPR/Cas9系统中使用,包括苹果[6]、藜麦[7]、大豆[8]、地钱[9]等。藏旭阳等[10-11]利用海岛棉的U6启动子构建GbU6-1P启动子并且利用PEG瞬时转化法将核心片段转入原生质体中,筛选出GbU6-7P启动子,以GGB为靶序列构建CRISPR/Cas9基因编辑载体并成功转入棉花的原生质体中,实现了对棉花内源靶基因GGB的定点编辑。王怡婷等[12]以大豆YT9启动子构建的CRISPR/Cas9基因编辑体系能够靶向敲除GmPDS11和GmPDS18双基因,实现大豆靶向基因编辑功能。Pompili等[13]利用拟南芥U6启动子驱动sgRNA的转录,对苹果中火疫病相关基因进行定点突变,以植株接种病菌后的发病情况来判断基因编辑的效果,统计了苹果的转基因和基因组编辑效率。
近年来,越来越多的金银花功能基因被相继克隆出来,为金银花的基因编辑奠定了基础。乔永刚等[14]从金银花中筛选出34个AP2转录因子,DREB和RAV均能响应低温胁迫,随着处理时间和组织部位的不同,表达量也存在差异,在一定程度上阐明金银花AP2转录因子响应低温胁迫的机制。Yan等[15]发现在不同浓度的盐胁迫下,HQT和PAL家族基因转录水平升高,绿原酸含量升高,药材生物量积累和质量都得到提高。Qi等[16]采用BLAST法和HMM法从忍冬转录组测序结果中得到CYP450基因,通过qRT-PCR筛选出5个与绿原酸生物合成相关的CYP450s酶进行探究,克隆得到3个LjC4Hs基因和2个LjC3Hs基因,有效地促进了绿原酸的生物合成。目前为止U6启动子已应用于多种植物,但是关于金银花U6启动子转录活性的研究还未见报道。金银花基因组中含有多种U6启动子,这些启动子的活性也并不相同,并且U6启动子具有物种偏好性,外源的启动子在金银花中可能难以高效启动转录,因此有必要克隆金银花自身的U6启动子,以便于开展金银花的基因编辑。
1 材料与方法
1.1 材料
本研究所用的金银花华金6号和烟草均由山东中医药大学中药资源研究室种植;大肠杆菌感受态细胞购自全式金生物技术有限公司;农杆菌感受态细胞购自上海唯地生物技术有限公司;基因组DNA提取试剂盒、纯化试剂盒和5minTMTA/Blunt-Zero Cloning Kit购自南京诺唯赞生物技术有限公司;质粒小提试剂盒购自天根生化科技(北京)有限公司;启动子测序由铂尚生物技术(上海)有限公司完成;DNA聚合酶采用北京聚合美生物科技有限公司生产的新型蓝色染料高保真Taq酶mix;染色液5-溴-4-氯-3-吲哚葡萄糖苷X-Gluc和10×PBS缓冲液购自北京索莱宝科技有限公司。
1.2 方法
1.2.1 启动子的克隆
取山东中医药大学药圃中生长良好的华金6号金银花叶片,采用植物基因组提取试剂盒提取金银花基因组DNA。利用设计好的引物(表1)和DNA聚合酶进行扩增,PCR程序为:98 ℃ 3 min;95 ℃ 30 s、55 ℃ 30 s、72 ℃ 1 min,32个循环;72 ℃ 5 min。反应体系为:DNA模板2.5 μL、正向引物和反向引物各1.25 μL、DNA聚合酶25 μL、水20 μL,共50 μL。用1%琼脂糖凝胶电泳检测正确后,胶回收试剂盒回收并纯化目的片段。回收完成后与pCE2 TA/Blunt-Zero载体连接、转化、一次活化,对重组质粒采用PCR方法鉴定,鉴定正确的质粒进行测序。用DNAMAN软件对测序结果进行分析,序列对比正确后进行二次活化。

表1 本研究使用的引物序列
1.2.2 U6启动子表达载体构建


图1 金银花不同截短U6启动子驱动GUS的融合表达载体构建
1.2.3 原生质体的制备
取无菌烟草叶片1 g,剪成约0.5 cm2的碎片,加入10 mL酶解液(纤维素酶1%,离析酶0.5%,甘露醇0.6 mol/L,MES 20 mmol/L,KCl 20 mmol/L,55 ℃水浴10 min后加入BSA 0.1%和CaCl21 mmol/L),26 ℃,60 r/min,避光酶解4 h[17]。酶解后的叶片过70 um细胞滤网,700 r/min离心5 min,弃上清后用W5(NaCl 154 mmol/L,CaCl2125 mmol/L,KCl 5 mmol/L)洗涤3次,加入1 mL MMM缓冲液(MgCl215 mmol/L,MES 0.1%,甘露醇 0.5 mmol/L),于4 ℃保存。烟草原生质体在4 ℃中保存的时间约为3 d[18]。
1.2.4 农杆菌侵染原生质体
取1 mL二次活化后的菌液,4 000 r/min离心5 min,弃上清;加入1 mL LB液体培养基悬浮菌体,静置2 min后离心,弃上清;加入1 mL MS液体培养基,悬浮菌体,28 ℃,200 r/min培养1~2 h;加入1 mL原生质体,25 ℃培养30 min,离心,弃上清,加入1 mL MS液体培养基悬浮后,28 ℃放置24 h。
1.2.5 原生质体染色和计数
在原生质体中加入1 mL的染色液(X-Gluc母液100 mL+50 mmol/L的PBS 900 mL;X-Gluc母液:5-溴-4-氯-3-吲哚葡萄糖苷 X-Gluc用N-N-二甲基甲酰胺配成20 mmol/L),37 ℃染色12 h。12 h后使用75%乙醇脱色3次,滴入血球计数板,镜检。
2 结果与分析
2.1 启动子克隆
从华金6号金银花中分别克隆出Chr01 U6-F1、Chr02 U61-F1、Chr02 U61-F2、Chr02 U62-F1和Chr02 U62-F2启动子片段,回收纯化后与T载体连接,PCR扩增后根据1%琼脂糖凝胶电泳检测结果(图2),将正确的质粒送去测序。测序结果符合预期。

注:M—D2000; 1—Chr01 U6-F1;2—Chr02 U61-F1;3—Chr02 U61-F2;4—Chr02 U62-F1;5—Chr02 U62-F2。
2.2 启动子序列分析
本研究克隆了金银花中3个候选的启动子,发现其都含有RNA聚合酶Ⅲ结合、识别所需要的TATA框和USE元件(图3),这表明克隆的3个金银花U6启动子可能都具有转录活性。

注:箭头为U6RNA的转录起始位点。
2.3 表达载体的构建
利用Hind Ⅲ和BamH Ⅰ双酶切构建好的T载体(图4),经验证每个启动子片段大小正确,回收、纯化目的片段并成功与PBI 121载体大片段相连(图5)。

注:M—D2000。
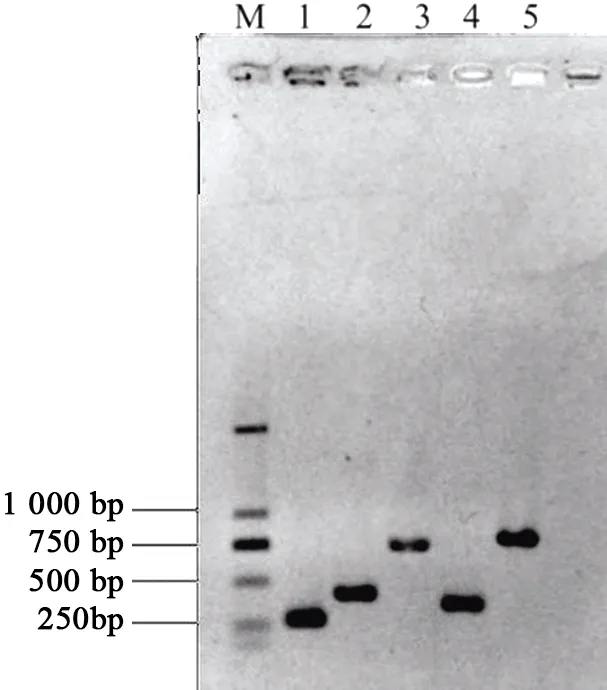
注:M—D2000;1—Chr01 U6-F1;2—Chr02 U61-F1;3—Chr02 U61-F2;4—Chr02 U62-F1;5—Chr02 U62-F2。
2.4 启动子转录活性分析
农杆菌侵染原生质体后,利用染色液对原生质体进行染色。本研究以CaMV35S启动子为阳性对照,以农杆菌为阴性对照,通过染色的结果可发现Chr01 U6-F1启动子具有较高的转录活性,Chr02 U62-F1启动子的转录活性低于Chr01 U6-F1启动子,Chr02 U61-F1、Chr02 U61-F2和Chr02 U62-F2启动子染色结果均未出现蓝色,表明这些启动子不具备转录活性或转录活性极低(图6)。

注:A—烟草原生质体;B—Chr01 U6-F1启动子转录后的原生质体染色;C—Chr02 U61-F1启动子转录后的原生质体染色;D—Chr02 U61-F2启动子转录后的原生质体染色; E—Chr02 U62-F1启动子转录后的原生质体染色;F—Chr02 U62-F2启动子转录后的原生质体染色;G—CaMV35S转录后的原生质体染色;H—农杆菌侵染后的原生质体染色。
3 小结与讨论
本研究从金银花全基因组测序结果中筛选出多个候选的U6启动子,利用BLAST和PCR验证,最终确立Chr01 U6-F1、Chr02 U61-F1、Chr02 U61-F2、Chr02 U62-F1和Chr02 U62-F2这5种U6启动子进行后续研究。通过构建不同长度启动子驱动GUS的植物融合表达载体和农杆菌瞬时转化等方法,最终筛选出Chr01 U6-F1这一长度为255 bp的高效转录启动子。在筛选出的启动子中Chr02 U62-F1的转录活性低于Chr01 U6-F1,其余启动子转录活性较低或不具有转录活性。针对Chr02 U62-F1和Chr02 U62-F2这两个同一启动子5′端不同截短染色情况差异提出假设,猜测其5′端有抑制因子或随着序列长度增加,组织转染难度也相应提高。研究表明,U6启动子在具备必要元件时即可拥有转录活性,而序列过长可能会存在抑制因子,使较长序列的U6启动子的转录活性受到抑制[19]。卞书迅等[20]在包含必要的USE和TATA盒前提下,把苹果10号染色体上的U6启动子从5′端进行不同长度的截短,发现长度为275 bp的U6启动子转录活性最高,而长度为1 527 bp和959 bp的U6启动子转录活性较低。卞书迅提出苹果U6启动子的5′末端存在一些抑制因子,抑制了这些启动子的转录活性。雷建峰等[21]从棉花中克隆得长度分别为1 166、1 119、1 134、1 214和1 176 bp的5种GbU6启动子,经过筛选,最终确定了转录活性较高,序列长度为1 134 bp的GbU6-5P,为构建棉花CRISPR/Cas9基因组编辑载体系统提供了有效的启动子。在番茄中,U6启动子被截取到200 bp左右时仍具有较高的转录活性[22],而在拟南芥中,U6启动子被截取到79 bp时还可用于基因编辑体系[23]。这些研究结果进一步验证了启动子5′端有抑制因子或随着序列长度增加组织转染难度也相应提高的猜想。
综上所述,本研究克隆并鉴定得到了一个具有高效转录活性的U6启动子,为在金银花中建立CRISPR/Cas9基因组编辑系统提供了理论依据,为后续对金银花功能基因研究、种质改良、新品种培育、提高抵抗生物及非生物胁迫能力奠定了坚实的基础。
猜你喜欢
环球时报(2022-09-20)2022-09-20
今日农业(2021年15期)2021-11-26
今日农业(2020年24期)2020-12-15
新农业(2020年21期)2020-11-19
小学生作文(低年级适用)(2019年12期)2020-01-18
江苏农业科学(2018年21期)2018-12-05
华人时刊(2016年13期)2016-04-05
小资CHIC!ELEGANCE(2015年15期)2015-09-01
特产研究(2015年1期)2015-04-12
现代检验医学杂志(2015年4期)2015-02-06
