
基于COI基因鉴定云南省43种蝇及其遗传关系分析
2022-04-06 02:37:46邵仕芳何鹏飞吴毅歆何月秋
西南农业学报 2022年2期
邵仕芳,何鹏飞,吴毅歆,何月秋
(1. 云南农业大学,昆明 650201;2.微生物菌种筛选与应用国家地方联合工程研究中心,昆明 650217)
【研究意义】蝇类属于双翅目昆虫,习性较为复杂,一般是昼间活动,适应力极强,陆生水生均有,且危害极为广泛,对农业、畜牧业产生极大的危害。例如:实蝇科及果蝇科的斑翅果蝇(Drosophilasuzukii)利用产卵棒插入瓜果蔬菜内部将卵产在瓜果蔬菜内,幼虫以果肉为食,对水果和瓜类蔬菜危害大,经济损失严重[1-4]。除斑翅果蝇外,果蝇科、蝇科、丽蝇科等科的大部分物种以腐烂的水果或植物体为食,产卵于果实表面或者缝隙中,雌虫繁殖能力强,幼虫蛀食成熟的果实,对水果或蔬菜经济危害大;蝇类可机械性传播伤寒、痢疾等多种疾病[5-14];此外,嗜尸性蝇类还具有法医学意义[5-9]。目前对蝇类害虫的鉴定主要基于成虫形态的鉴定,但对复杂的形态学特征和相似的种类鉴定有困难。麻蝇多数种类形态相近[5,12],主要依赖雄蝇尾器鉴定,雌性麻蝇没有统一的形态分类标准。在口岸日常检疫截获的蝇类多以幼虫、卵、蛹等虫态及大量残缺不全的虫体。传统分类很难准确鉴定物种。活体的幼虫、蛹和卵等饲养成成虫的周期长,会大大提高鉴定成本,直接影响口岸检疫部门的效率和质量。【前人研究进展】DNA条形码是分类学中辅助物种鉴定的新技术,是利用线粒体细胞色素C氧化酶I基因(COI)的特定区段进行物种分子鉴定的方法。2003年由加拿大动物学家Paul Hebert提出,该基因片段已经在果实蝇属[4]、嗜尸性麻蝇[5]、蝇类[6]、丽蝇科[10]、双翅目[12]等类群中,证明了其鉴定物种的有效性。与传统形态学的方法相比,DNA条形码技术能对昆虫各生命形态,以及对保存不完整的昆虫标本进行准确的物种鉴定,具有明显优势。对形态学特征差异性不明显,或成虫虫体不完整,依靠传统的形态学鉴定物种受限较大,而对不完整的昆虫标本或在幼虫、卵、蛹等阶段形态差异不大的,利用DNA条形码进行鉴定就具有明显优势。【本研究切入点】对诱捕到的蝇类、先经形态学物种分类,再利用COI基因片段进行基因鉴定,再依据进化树遗传距离等确定快速有效鉴定蝇类的方法。【拟解决的关键问题】对来自云南省河口县、大理市银桥镇、剑川县、蒙自市和元江县的蝇类标本,采用形态学和基因水平上的技术手段,能准确鉴定其蝇类科、属、种的归类,为后续快速鉴定提供依据。
1 材料与方法
1.1 试验材料
于2017年10月至2018年10月,在云南省的河口县、大理市、剑川县、蒙自市和元江县,应用微生物菌株Y2、432、460发酵液诱捕得到双翅目蝇类,昆虫标本保存于微生物菌种筛选与应用国家地方联合工程研究中心。
1.2 昆虫总DNA提取
将诱捕到的双翅目蝇类昆虫依据形态进行归类,利用平衡酚-Tris饱和酚法[8,10],每1个类型提取2~3个个体的DNA。
1.3 PCR扩增与测序
选取通用DNA条形码COI基因对所有类型样本的DNA进行扩增。引物为:ffly-1:5′-TATGTACTACCATGAGGACAAATATC-3′,ffly-2:5′-ATTACACCTCCTAATTTATTAGGAAT-3′(上海捷瑞生物技术有限公司合成)。PCR反应的总体积为40 μL,其中含有1.5 μL DNA模板,引物(10 pmol/L)各2 μL,4 μL 10×EasyTaqBuffer,4 μL Mg2+(25 mmol/L),3.2 μL dNTP,0.4 μLTaqDNA 聚合酶(10 U/μL)。PCR扩增条件为94 ℃预变性4 min;94 ℃变性40 s,52 ℃退火40 s,72 ℃延伸1 min,40个热循环;最后在72 ℃延伸10 min,空白对照为ddH2O代替模板。扩增产物用1%的琼脂糖凝胶电泳检测,目的片段为mtDNACOI基因中650 bp序列。
1.4 序列处理与分析
将获得的序列用DNAMAN软件阅读,利用MEGA 6.06进行比对,统计所有种类COI基因部分序列的长度、GC 含量、多态位点、简约信息位点等参数。对不同蝇类之间COI序列遗传差异进行分析。利用NJ 法构建系统发育树,采用自展法重复检测,自展参数设置为1000次,验证结果。
2 结果与分析
2.1 双翅目昆虫种类
经1年时间,在云南省河口县、大理市、剑川县、蒙自市和元江县5地共诱捕到蝇类7科21属43种(表1),其中果蝇科3属11种,实蝇科1属6种,丽蝇科4属7种,厕蝇科1属5种,蝇科5属6种,麻蝇科3属3种,花蝇科5属5种。

表1 诱捕到的双翅目蝇类昆虫种类

续表1 Continued table 1
2.2 COI序列特征
43种蝇的线粒体COI基因5’端长度为650 bp的条形码序列分析(表2)表明:该序列编码211个氨基酸。其中,果蝇科、实蝇科、丽蝇科、厕蝇科、蝇科、麻蝇科、花蝇科7个科COI基因片段的A+T平均含量分别为65.6%、64.1%、68.9%、67.7%、68.9%、67.0%、68.5%,表现出明显的A+T碱基偏倚性。
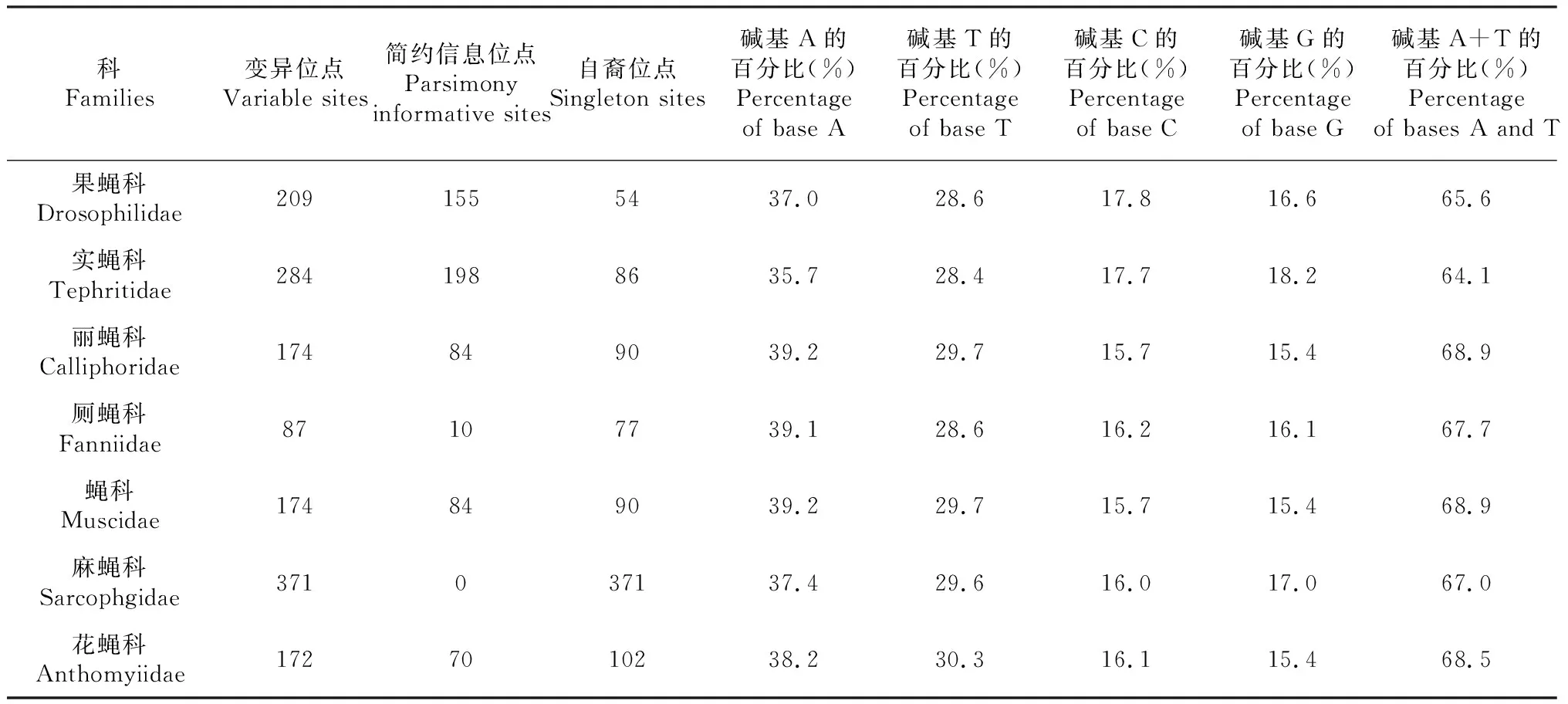
表2 双翅目蝇类不同科的昆虫COI序列特征
2.3 蝇类遗传关系分析
根据研究获得的蝇类COI基因片段序列,对蝇类不同科进行种间遗传距离计算(表3)。11种果蝇种间遗传距离为0.013~0.190,平均0.151;种内遗传距离为0.002~0.012,平均0.006。种间遗传距离是种内遗传距离的25.2倍。其中D.suzukii和D.subpulchrella遗传距离为0.013,明显小于其余果蝇之间的遗传距离,根据形态学看,这2种果蝇极其接近,两者为近源姐妹种。
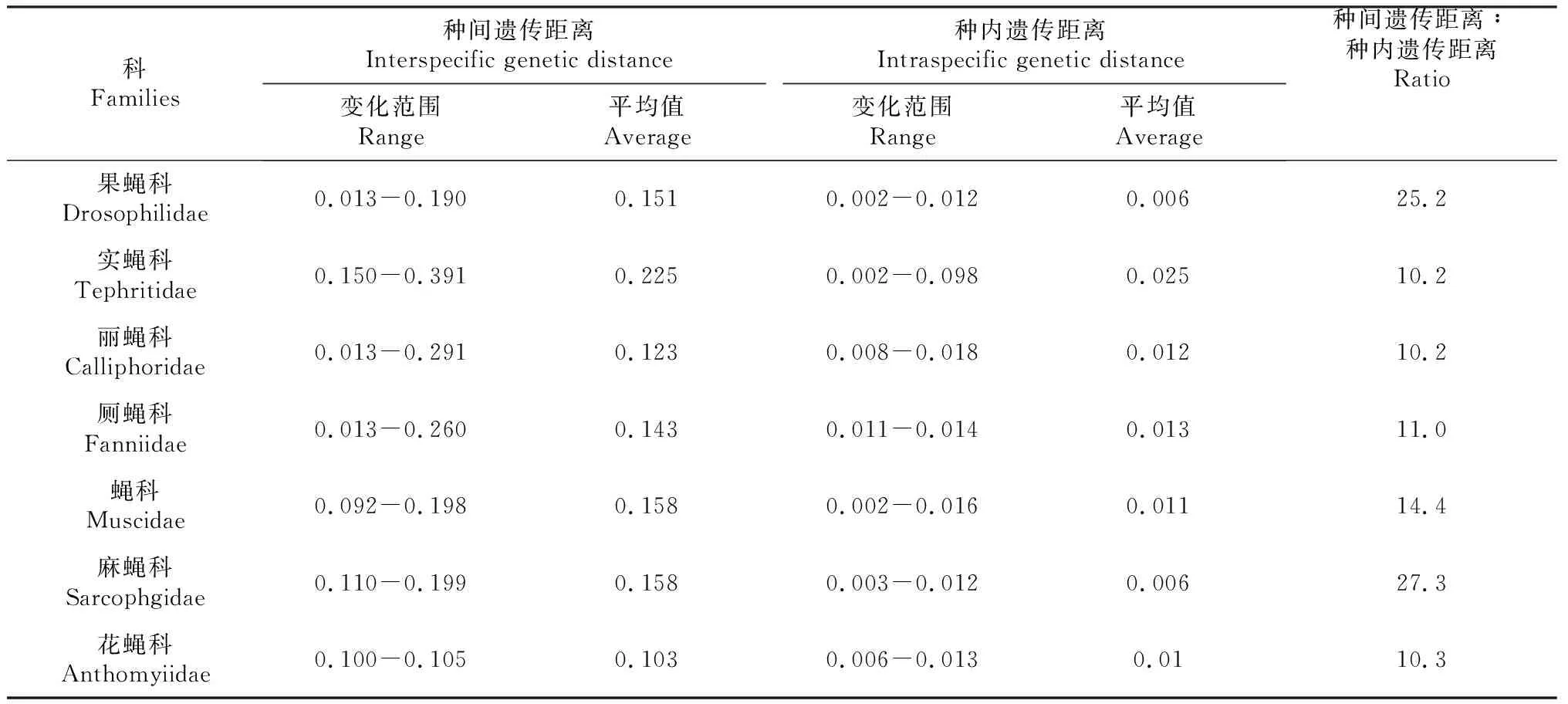
表3 蝇类各科种间及种内遗传距离
对6种实蝇种间及种内遗传距离分析表明,它们种间遗传距离为0.150~0.391,平均0.255;种内遗传距离为0.002~0.098,平均0.025。种间遗传距离是种内遗传距离的10.2倍。研究结果与双翅目其他类群的较为一致。
7种丽蝇种间遗传距离为0.013~0.291,平均0.123;种内遗传距离为0.008~0.018,平均0.012。种内遗传距离是种间遗传距离的10.2倍。绿蝇属的铜绿蝇与丝光绿蝇的遗传距离为0.013,小于其余丽蝇科的种间遗传距离。
5种厕蝇种间遗传距离为0.013~0.260,平均0.143;种内遗传距离为0.011~0.014,平均0.013。种内遗传距离是种间遗传距离的11.0倍。夏厕蝇与元厕蝇遗传距离为0.013,接近种内平均遗传距离。这一结果与其形态学分类一致,夏厕蝇与元厕蝇在形态学上极为相似。
对6种蝇种间及种内遗传距离分析表明,它们种间遗传距离为0.092~0.198,平均0.158;种内遗传距离为0.002~0.016,平均0.011。种间遗传距离是种内遗传距离的14.4倍。刺足齿股蝇与中华毛庶芒蝇、东方芒蝇遗传距离分别为0.092和0.094;东方芒蝇与中华毛庶芒蝇遗传距离为0.096。遗传距离相较于其他蝇科的种间遗传距离更小。
对麻蝇科的黑尾麻蝇、野亚麻蝇和白头亚麻蝇研究显示,其中白头亚麻蝇和野亚麻蝇,黑尾麻蝇和野亚麻蝇、白头亚麻蝇和黑尾麻蝇的遗传距离分别为0.100、0.105、0.103,平均遗传距离为0.103;野亚麻蝇、白头亚麻蝇和黑尾麻蝇的种内遗传距离分别为0.011、0.006、0.013,种内平均遗传距离为0.010,种间遗传距离是种内遗传距离的10.3倍。
5种花蝇种间遗传距离为0.110~0.199,平均0.158,种内遗传距离为0.003~0.012,平均0.006,种间遗传距离是种内遗传距离的27.3倍。
2.4 系统发育树的构建
由图1可知,从科水平上看,并未像形态学一样形成7个大的独立分支,实蝇科、果蝇科、厕蝇科、蝇科和丽蝇科大部分物种聚集成一支,各科间分支上,部分物种存在一定的同源交叉,尤其是花蝇科、丽蝇科、蝇科3科物种同源交叉明显。从属水平上看,果蝇科3个属各聚集成一簇,各属均形成各自的单系,与形态学结果一致;厕蝇科的厕蝇属单独聚集成一簇;蝇科的芒蝇属和腐蝇属单独聚集成一簇,符合形态学分类要求。从种的水平上看,同一种不同样本大多聚集在一起,大多数置信度在90%以上,各种之间区别开来,丝光绿蝇与铜绿蝇聚成一簇,2种存在同源性交叉,印证了二者种间遗传距离较小(0.013)的原因,斑翅果蝇与其姊妹种D.subpulchrella聚集在一起支持率达94%,印证2个种亲缘关系极其相近,夏厕蝇和元厕蝇聚集在一起,聚集置信度为91%。
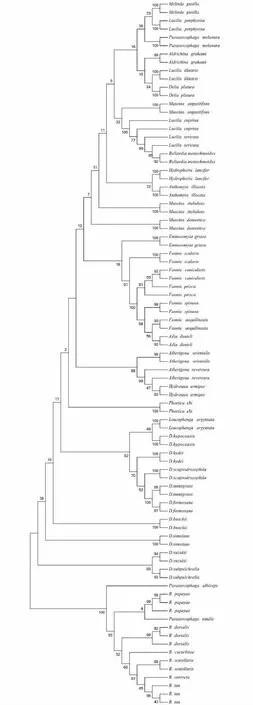
分支上数值示自展检验置信度,标尺示遗传距离The numbers at the nodes indicate the bootstrap value. Scale bar indicates the genetic distance图1 依据COI基因序列采用邻接法构建的云南省43种蝇类系统发育树Fig.1 Neighbor-joining tree for 43 species of flies from Yunnan province, China based on COI gene sequences
3 讨 论
3.1 COI基因鉴定蝇类昆虫的优点
应用微生物发酵液能诱捕到7科21属43种双翅目蝇类昆虫,表明该类诱捕剂具有广泛诱蝇能力。利用DNA条形码对双翅目系统发育学的研究,主要集中在各科的水平上,如实蝇科[2,4]、果蝇科[3,15]、丽蝇科[7,9]等,而针对各科之间发育学研究并不多,为此利用DNA条形码COI基因对双翅目的果蝇科、实蝇科、丽蝇科、厕蝇科、蝇科、花蝇科、麻蝇科43种蝇类的种间及科间系统发育进行了初步探讨。采用DNA条形码分析时,种内、种间遗传距离是进行物种鉴定的重要依据。种间遗传距离大于种内遗传距离是利用DNA条形码鉴别物种的条件之一,Hebert等[18]提出种间遗传距离要达到种内遗传距离的10倍。研究显示,7科21属43种蝇类种内遗传距离都很小,而种间遗传距离较大,果蝇科、实蝇科、丽蝇科、厕蝇科、蝇科、花蝇科和麻蝇科等各科的种间平均遗传距离分别是种内遗传距离的25.2、10.2、10.2、11.0、14.4、27.3、10.3倍,科间鉴定结果符合要求和具有可行性。
结合建树方法评估COI序列的鉴定能力,研究结果表明:果蝇科中的斑翅果蝇与其姊妹种D.subpulchrella种内遗传距离均为0.005,种间遗传距离为0.013,且进化树上两者聚集在一起,Zhang等[13]和Conner等[14]的研究也有相同结果。蝇科中,紫绿蝇与亮绿蝇种间遗传距离为0.089,2个物种在进化树中聚集在一起,形成2个支系,种内同源性为98.2%~100%,利用COI基因可以将2个物种准确鉴定。铜绿蝇与丝光绿蝇同源性高(种间遗传距离为0.013),在进化树上不能完全分开,国内外研究结果也相似[16-17]。厕蝇科中,元厕蝇与夏厕蝇种间遗传距离为0.013,在进化树上聚集在一起,均为夏厕蝇种团。说明对果蝇科、丽蝇科、厕蝇科中一些近缘种利用COI基因鉴别存在缺陷。
3.2 COI基因鉴定蝇类昆虫的局限性
对于同源性较强的物种,利用COI基因序列鉴定会有一定的局限性,最大的难点是没有统一的分子标签来建立统一的鉴定标准。研究显示部分物种的种间和种内差异不明显,在进化分支上存在重叠,在种间水平的鉴定分类上利用COI基因序列存在一定的限制,尤其是对于亲缘关系较近,形态相似的物种。对此,可以采取扩大基因选取的范围、尝试多个分子标记相互印证,采取分子和形态学鉴定相结合是一个有效的方法。分类学家已将不同种形态学上给予足够的分类标准,但是对形态相似或者残体的物种,依靠形态学无法鉴定的,就要利用COI基因序列的差异性并与形态学相结合,可以达到对蝇类准确鉴别的目的。
利用COI基因片段,鉴定了云南省河口县、蒙自市和元江县应用微生物诱剂诱捕到的双蝇目昆虫7科22属43种。然而在科水平上,7个科的各个物种样本并未完全聚集成一支,部分物种存在同源交叉,尤其是亲缘关系较近的物种,系统进化树分支与形态学分类不完全一致,说明利用COI基因片段对蝇类科水平的鉴定存在一定的局限性。从种水平上看,除果蝇科、丽蝇科、厕蝇科部分种的鉴定不明确之外,其余物种均能较好区分,但种及种下水平或近缘物种鉴定存在同源交叉,鉴定仍然存在一定困难。因此,利用DNA条形码针对不同种及种下水平之间的分子发育关系尚待研究。
4 结 论
采取COI基因序列的差异性与形态学相结合的方法,鉴定了云南省河口县、大理市、剑川县、蒙自市和元江县5地应用微生物诱剂诱捕到的双翅目蝇类昆虫有7科22属43种,基本明确了各蝇类间的遗传关系,为利用COI基因鉴定双翅目蝇类昆虫奠定了科学依据。
猜你喜欢
学苑创造·A版(2023年10期)2023-11-04 13:14:04
植物研究(2023年5期)2023-09-09 08:01:22
大自然探索(2023年11期)2023-03-01 09:04:36
河南师范大学学报(自然科学版)(2022年5期)2022-08-08 14:07:42
学苑创造·A版(2022年3期)2022-03-29 23:32:16
学苑创造·A版(2019年6期)2019-07-11 01:07:39
中国继续医学教育(2015年2期)2016-01-06 01:36:16
杂草学报(2015年2期)2016-01-04 14:57:55
振动、测试与诊断(2014年6期)2014-03-01 01:14:50
中国海洋大学学报(自然科学版)(2014年12期)2014-02-28 12:22:02
