
天山云杉林下优势草本植物化学计量内稳性特征
2022-04-02 06:10阿里木买买提卡哈尔曼恰依扎旦常顺利
西北林学院学报 2022年2期
阿里木·买买提,李 翔,卡哈尔曼·恰依扎旦,常顺利
(1.新疆林业科学院 森林生态研究所,新疆 乌鲁木齐 830063;2.哈密市生态修复保护中心,新疆 哈密 839000;3.新疆大学 资源与环境科学学院,绿洲生态教育部重点实验室,新疆 乌鲁木齐 830046)
生物生长过程的实质是生物体最基本的结构功能性物质C、N、P、K 等的积累与其相对比例的调节[1-2]。生态化学计量学是从元素组成的角度探讨生态过程和生态作用中化学元素平衡的理论[3-5],为研究生态系统中土壤—植物—环境三者之间相互作用提供了新手段。内稳态理论作为生态化学计量学主要理论,阐述了生物在变化的环境中具有维持体内元素组成在相对狭窄的范围内稳定的能力[6-8]。在国外化学计量内稳性研究多集中于藻类、浮游动物和草本植物[9],对高等植物的研究较少,在物种水平上的研究更为缺乏[10];国内内稳性研究多集中于草原和湿地生态系统中的草本植物[11-12],针对森林生态系统中的草本植物研究甚少。李玉霖等[13]、刘洋等[14]研究发现内稳性的高低受植物自身器官和环境因子共同影响,影响因子的差异使得不同植物神态化学计量内稳性对环境的响应不一致。
森林生态系统中草本植物对促进能量流动和物质循环具有不可忽视的作用[15],草本植物对于环境的变化比乔木层更为敏感,更容易在较短时间内反映环境带来的影响[16]。天山云杉是天山山脉的地带性植被,在水源涵养、保持水土等生态功能方面发挥着重要作用,其林下分布着季相变化明显的草本层。目前,对草本层物种组成和区系特征方面已开展较多研究[17-18],但对草本植物的养分适应机制研究较少。因此,本研究以天山北坡中部天山云杉林下的优势草本植物天山羽衣草(Alchemillatianschanica)为研究对象,比较不同海拔天山羽衣草的C、N、P、K含量及计量比的差异性,并结合土壤养分分析判断该区域草本植物生长过程中的营养限制因子,揭示其生态适应的化学计量学机制,为合理制定天山北坡生态系统保护与修复措施提供理论参考。
1 材料与方法
1.1 研究区概况
研究区位于天山北坡乌鲁木齐县中国森林生态系统研究网络(CFERN,China forest ecology research net)天山森林生态系统定位研究站(87°07′-87°28′E,43°14′-43°26′N)。该区属温带大陆性气候,年总辐射量达5.85×105J/(cm2·a),年均气温约为2~3 ℃,历年极端最高温为30.5 ℃,极端最低温为-30.2 ℃,年降水量400~600 mm,雨季集中在6-8月,最大积雪深度为65 cm,年蒸发量980~1 150 mm,年均相对湿度65%,无霜期89 d,≥10 ℃积温1 170.5 ℃。该区植被类型以天山云杉(Piceaschrenkiana)纯林为主,森林覆盖率达60%,郁闭度0.4~0.8。林下草本主要有天山羽衣草(A.tianschanica)、老鹳草(Geraniumrotundifolium)、羊角芹(Aegopodiumpodagraria)等,林下土壤为山地灰褐色森林土。
1.2 样品采集
2020年8月,以天山北坡中部不同海拔的天山羽衣草为研究对象,选择4个海拔区域(Ⅰ:2 000~2 200 m、Ⅱ:2 200~2 400 m、Ⅲ:2 400~2 600 m、Ⅳ:2 600~2 800 m),在每个区域内随机设置10个采样点,各采样点水平距离大于100 m。在每个采样点布设1 m×1 m的样方进行草本调查,调查项目包括株数、高度、盖度及群落高度。调查结束后采集样方天山羽衣草地上部(枝叶)和根系带回实验室进行养分分析。在每个样方随机选取3个点,去除地表枯落物和植物残体后,用直径为5 cm的土钻取地表0~20 cm的土壤,将每个样方的土壤分别混合,室温下自然风用于土壤养分测定。
1.3 样品分析
将采集的植物样品在105 ℃烘箱中杀青30 min,然后65 ℃下烘干至恒重,粉碎后备用。植物有机碳测定采用K2Cr2O7-H2SO4氧化法;全N的测定采用H2SO4-HCLO4消化-靛酚蓝分光光度法;全P的测定采用H2SO4-HCLO4消化-钼锑抗比色法;全K的测定采用H2SO4-HCLO4消化-火焰光度。
采集的土壤样品自然风干后,经研磨过0.150 mm筛备用。土壤有机碳的测定采用K2Cr2O7氧化-外加热法;全N的测定采用半微量开氏法;全P的测定采用酸溶-钼锑抗比色法;土壤全K的测定采用NaOH碱熔-火焰光度计法[19]。
1.4 数据处理
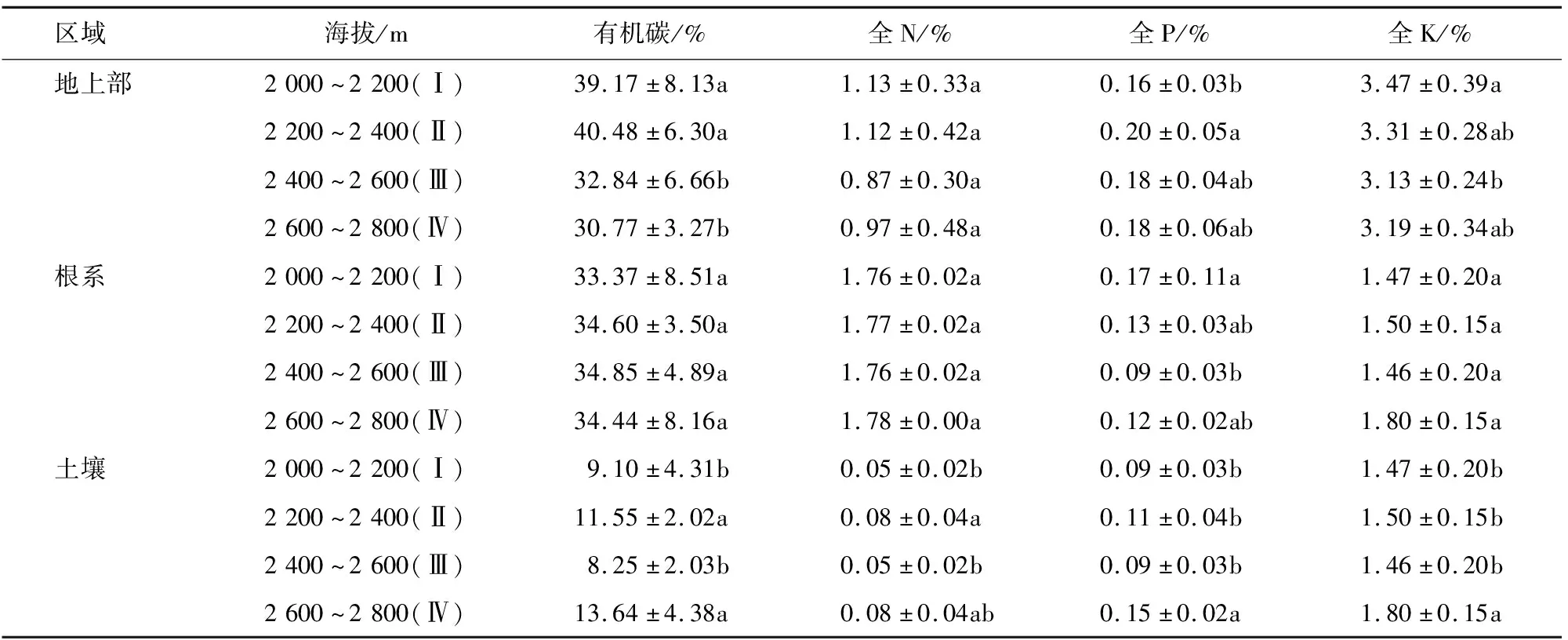
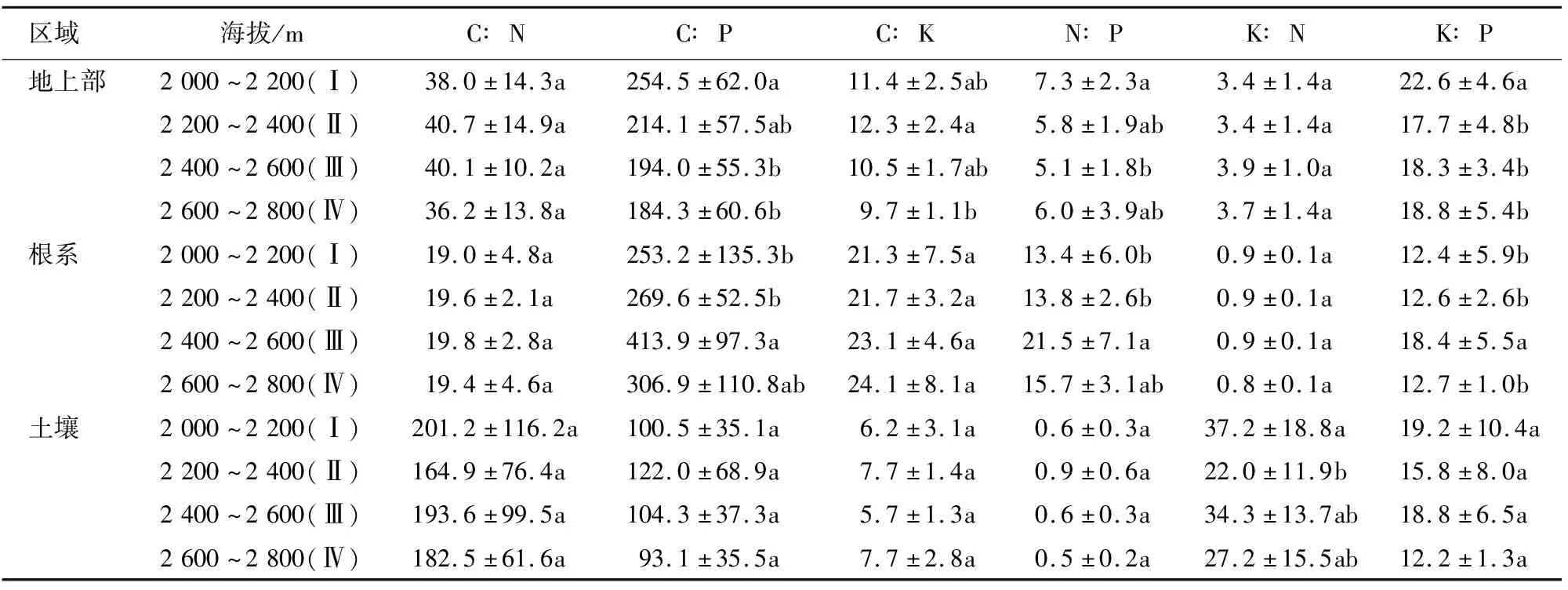
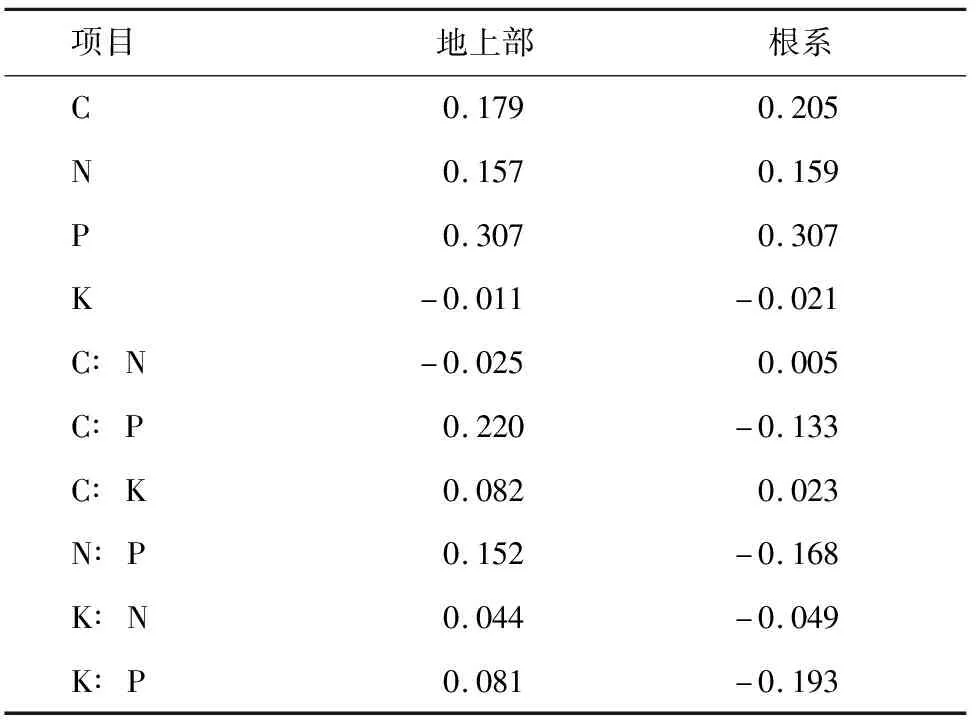
内稳性指数用内稳性模型y=cx1/H通过回归分析计算[20]。式中,x为土壤中C、N、P、K含量(%)及其比值,y为植物地上部对应的C、N、P、K含量(%)及其比值,c为常数,H为植物内稳性指数,H>4为稳态型、2 采用Excel 2007和SPSS 17.0进行数据分析和处理。植物与土壤化学计量比均采用百分含量比表示。采用单因素方差(one-way ANOVA)分析比较不同海拔天山羽衣草和土壤养分化学计量内稳性的差异(P<0.05);采用Pearson相关分析方法计算天山羽衣草与土壤生态化学计量特征之间的相关系数。 不同海拔区域,天山羽衣草地上部、根系和土壤的养分含量存在差异(表1)。低海拔区域(Ⅰ和Ⅱ)天山羽衣草地上部养分含量高于高海拔区域(Ⅲ和Ⅳ)。其中,C含量变化范围为30.77%~39.17%,区域Ⅰ和Ⅱ C含量显著高于Ⅲ和Ⅳ;N含量变化范围为0.87%~1.13%,不同海拔区域差异不显著;P含量区域Ⅱ显著高于区域Ⅰ;K含量区域Ⅰ显著高于区域Ⅲ。天山羽衣草根系养分含量高海拔区域(Ⅲ和Ⅳ)高于低海拔区域(Ⅰ和Ⅱ)。其中,不同海拔高度C、N和K含量差异均不显著;P含量区域Ⅰ显著高于Ⅲ。土壤C含量变化范围为8.25%~13.64%,区域Ⅱ和Ⅳ显著高于Ⅰ和Ⅲ;土壤N含量区域Ⅱ显著高于Ⅰ和Ⅲ;土壤P含量变化范围为0.09%~0.15%,土壤K含量变化范围为1.46%~1.80%,区域Ⅳ的P、K含量均显著高于Ⅰ、Ⅱ和Ⅲ。 表1 不同海拔天山羽衣草和土壤C、N、P、K养分含量 天山羽衣草地上部C∶N随海拔升高呈先上升后下降的趋势,不同海拔之间的差异不显著(表2)。C∶P随海拔上升逐渐下降,区域Ⅰ的C∶P显著高于Ⅲ和Ⅳ;区域Ⅱ的C∶K显著高于Ⅳ;区域Ⅰ的N∶P显著高于Ⅲ;不同海拔区域K∶N差异不显著;K∶P随海拔上升呈现先降低后升高的趋势,区域Ⅰ的K∶P显著高于其他海拔区域。天山羽衣草根系C∶N、C∶K和K∶N在不同海拔区域之间差异均不显著。C∶P、N∶P和K∶P在区域Ⅲ均为最大值,且区域Ⅲ的C∶P和N∶P显著高于区域Ⅰ和Ⅲ,区域Ⅲ的K∶P显著高于Ⅰ、Ⅱ和Ⅲ。不同海拔区域C∶N、C∶P、C∶K、N∶P和K∶P差异不显著。 表2 不同海拔天山羽衣草及土壤中养分计量比 天山羽衣草地上部、根系养分含量及计量比与土壤中对应养分含量及计量比相关性均不显著(表3)。天山羽衣草地上部与根系C、N、P含量与土壤中对应的C、N、P含量呈正相关,其中天山羽衣草P含量与土壤相关系数较大;K含量与土壤K含量呈负相关。天山羽衣草地上部养分含量计量比与土壤对应计量比除C∶N外,均呈正相关性。根系养分含量计量比与土壤养分对应计量比C∶P、N∶P、K∶N、K∶P呈负相关。 表3 天山羽衣草不同器官养分特征与土壤养分含量特征的相关系数 分析天山羽衣草地上部和根系养分元素内稳性指数,结果表明(表4):天山羽衣草地上部C、N、P、K及其计量比均为稳态性,且元素计量比内稳性高于元素本身,C∶N和N∶P内稳性较高。根系P和K∶P均为弱稳态性,C、N、C∶P、C∶K和N∶K为稳态性,N元素内稳性最高。天山羽衣草地上部、根系中K元素、根系C∶N和K∶N不具内稳性。 表4 天山羽衣草地上部和根系内稳性 3.1.1 C、N、P、K生态化学计量特征的变化 刘新圣等[21]研究表明随海拔升高可引起植物水热配比的变化,进而影响植物的生理过程。本研究中天山羽衣草地上部和根系养分含量与新疆草原植物和中国草地生态系统植物相比,C和N含量降低,K含量升高。天山羽衣草地上部养分含量表现为海拔2 000~2 400 m区域大于海拔2 400~2 800 m区域,且养分含量随海拔变化差异显著;根系的养分含量表现为:高海拔区域(2 400~2 800 m)大于低海拔区域(海拔2 000~2 400 m),养分含量随海拔变化差异不显著。低海拔区域,水热条件良好,有利于增加植物体内营养物质的积累;高海拔区域,水热条件变差温度降低,植物光合作用减弱,营养物质的积累减少。植物地下部分作为养分库器官,受温度和水分影响较小,因而在不同海拔梯度之间差异不显著[22]。本研究中天山羽衣草地上部养分含量与根系相比,除N含量以外,均为地上部大于根系,与何亚婷等[23]的研究结论基本一致。叶片是植物与外界进行物质和能量交流的主要器官,营养元素在地上部中的分配高。而N含量相反则,可能是由于植物体内的N主要是通过植物的根系从土壤中吸收然后向各器官运输[24]。 申静霞等[25]研究表明温度和降水是植物生长的重要生态因子,高温湿润的环境使得土壤有机质积累相对容易;而随海拔升高,水热条件逐渐降低,凋落物养分循环减弱,使得土壤养分含量呈现下降的趋势。但本研究中土壤养分含量随海拔波动性变化,海拔Ⅱ和Ⅳ的养分含量高于海拔Ⅰ和Ⅲ,与聂明鹤等[26]在小尺度的研究结果一致,可能是样地内物种组成以及地形等其他因素产生一定的影响。 3.1.2 C、N、P、K生态化学计量比特征的变化 生态化学计量比值可用于揭示植物的营养状况和区域生境的养分供给能力[27]。在水热条件良好的低海拔区域,植物生长发育快,养分利用效率较高。本研究中天山羽衣草地上部C∶N和K∶N在不同海拔之间差异不显著,C∶P、C∶K、N∶P、K∶P呈低海拔区域大于高海拔区域的规律。根系C∶N、C∶K和K∶N在不同海拔梯度之间差异均不显著,而C∶P、N∶P和K∶P在海拔2 400~2 600 m为最大值,反映了P对于根系生长的重要性。本研究中天山羽衣草地上部的N∶P变化为5.1~7.3,地上部分生长受到N限制;根系的N∶P变化为13.4~21.5,根系生长主要受到P限制。与刘万德等[28]研究结论一致,植物生长初期受N限制,而在演替后期主要受P限制。 土壤元素化学计量比反映了土壤释放矿化养分的能力[29]。本研究中虽然不同海拔土壤C∶N、C∶P、C∶K、N∶P和K∶P差异不显著,但仍表现为低海拔区域大于高海拔区域的趋势。因此,认为低海拔良好的水热配比条件有利于土壤养分的矿化。土壤N∶P的变化为0.5~0.9,与其他土壤N∶P研究相比该比值偏小[30-31],造成这一结果的原因可能来自气候与植物的双重影响。已有的针对中国不同气候土壤C、N、P生态化学计量特征的研究表明,N∶P在不同气候区的变异较大[32-33],本研究区域土壤N∶P具有地域特点。此外,因植物与土壤的反馈作用,植物会影响土壤N和P的贮存与矿化。 3.1.3 天山羽衣草内稳性特征 土壤为植物提供了生长必需的养分,植物体内养分特征与土壤养分具有一定相关性。但在本研究中,天山羽衣草地上部和根系养分特征均与对应土壤养分特征相关性不显著。 天山羽衣草C、N、P、K元素计量比的内稳性高于元素本身,说明植物在生长过程中按一定比例调控自身营养元素,C、N(稳态型)内稳性高于P(弱稳态型)。李贵才等[34]研究表明,植物N含量比P含量具有更高的自我调控系数,表现出较强的化学内稳态。同时,天山羽衣草地上部与根系化学元素内稳性相比,根系C、N、C∶P和C∶K内稳性更高。石贤萌等[35]对哀牢山中山湿性常绿阔叶林2种优势幼苗研究表明,幼苗根和茎的N内稳性比叶片更高;贡璐等[6]在对塔里木河绿洲棉花的研究中发现,苗期根系N、P元素内稳性均比叶片高。 天山北坡云杉林下天山羽衣草地上部和根系C、N、P、K含量分别为30.77%~39.17%、0.87%~1.13%、0.16%~0.20%、3.13%~3.47%和33.37%~34.85%、1.76%~1.78%、0.09%~0.17%、1.46%~1.80%。 天山羽衣草地上部养分含量低海拔区域(Ⅰ和Ⅱ)高于高海拔区域(Ⅲ和Ⅳ);根系养分含量高海拔区域(Ⅲ和Ⅳ)高于低海拔区域(Ⅰ和Ⅱ);土壤养分含量海拔Ⅱ、Ⅳ高于海拔Ⅰ、Ⅲ。 天山羽衣草地上部C∶P、C∶K、N∶P、K∶P计量比低海拔区域均大于高海拔区域;根系C∶N、C∶K和K∶N计量比在不同海拔区域之间差异均不显著,C∶P、N∶P和K∶P均随海拔升高先增大后减小;不同海拔区域间土壤元素计量比差异不显著。 天山羽衣草地上部N∶P计量比为5.1~7.3,本研究认为N元素为天山羽衣草地上部生长限制元素;根系N∶P计量比为13.4~21.5,P元素为天山羽衣草根系生长的限制元素。 天山北坡云杉林下优势草本植物天山羽衣草C、N、P、K计量比的内稳性高于元素本身;C、N(稳态型)内稳性高于P(弱稳态型)。 综上所述,天山羽衣草地上部和根系养分含量及其化学计量比内稳性存在一定变化规律,但与温度和降水等环境因子的定量关系如何仍有待进一步研究。2 结果与分析
2.1 不同海拔天山羽衣草及土壤养分含量变化特征
2.2 不同海拔天山羽衣草及土壤养分计量比
2.3 天山羽衣草内稳性特征

3 结论与讨论
3.1 讨论
3.2 结论
猜你喜欢
军事文摘(2022年14期)2022-08-26
纺织学报(2022年8期)2022-08-26
散文诗(2021年22期)2022-01-12
黄河之声(2021年12期)2021-10-25
动漫星空(兴趣百科)(2020年9期)2020-09-28
学生天地(2020年6期)2020-08-25
意林·全彩Color(2019年11期)2019-12-30
环球时报(2019-05-23)2019-05-23
奥秘(2015年2期)2015-09-10
吐鲁番(2014年2期)2014-02-28
