
西瓜后绿突变体光合特性分析
2022-04-01 11:26高美玲包秀萍刘秀杰刘继秀
西北农林科技大学学报(自然科学版) 2022年3期
徐 铭,高美玲,2,郭 宇,包秀萍,刘秀杰,刘继秀,高 越
(1 齐齐哈尔大学 生命科学与农林学院, 黑龙江 齐齐哈尔 161006;2黑龙江省抗性基因工程与寒地生物多样性保护重点实验室,黑龙江 齐齐哈尔 161006;3齐齐哈尔市园艺研究所, 黑龙江 齐齐哈尔 161006)
叶色突变是植物最常见的突变之一,现已在多种植物中被发现,目前报道的叶色突变体有白化、黄化、浅绿、白翠、绿白、黄绿、绿黄和条纹共8种类型[1]。
关于叶色突变体的大部分报道集中在大田作物中,其中除少部分叶色突变为细胞质遗传外,大部分叶色突变为细胞核遗传[2]。如已在水稻中发现了140种叶色突变体,分布在12个染色体上,其中有少部分叶色突变体已被克隆[3-5]。在葫芦科作物黄瓜中已经发现了17种叶色突变体,其中有4种致死性的叶色突变(al、cd、gc和pl)和13种能稳定遗传的叶色突变(yp、yf、g、yc-1、yc-2、ygl、lg-1、lg-2、v、v-1、vvi、vyl、YL)[6-10],甜瓜(ypl、MT)[11-12]、西瓜(YL、I-dg、dg)[13-17]也陆续出现叶色突变体的报道。叶色突变的形成原因不一,主要原因包括叶绿体发育受阻、叶绿素合成途径受阻、叶绿素降解等,这都可以导致植物叶片叶绿素含量降低,从而使叶片颜色无法呈现绿色[18]。叶色突变已应用在杂种鉴定中,可作为叶色标记分辨出杂交品系中的自交种[13]。后绿突变体具有转绿特性,对植株光合能力影响较小,因此后绿性状也适合作为叶色标记应用于雄性不育系和优良品系[19]。
齐齐哈尔大学西甜瓜育种课题组于田间发现一个后绿突变西瓜品系(命名为63),田间种植表明该叶色突变能稳定遗传。本研究以该后绿突变品系63为试验材料,由于缺少该突变体的野生型品系,故选用绿叶品系Liu-2作为对照,通过测定其光合色素含量、光合参数及荧光动力学参数并进行比较分析,以期初步了解此后绿突变体的光合特性,为进一步探究西瓜的叶色后绿机制提供理论依据。
1 材料与方法
1.1 供试材料
本试验选用田间发现的西瓜后绿突变体63,以正常绿叶品系Liu-2为对照,取2个品系各20粒完整饱满的种子,温汤浸种并催芽,播种于育苗穴中,待两叶一心时定植于黑龙江齐齐哈尔市曙光村大棚,株距14 cm,行距20 cm,常规田间管理。如图1所示,在植株伸蔓期时,以西瓜叶色后绿突变体63第三节位(Ⅰ时期)黄化叶片、第九节位(Ⅱ时期)绿色叶片为试验材料,用于后绿突变体转绿前后光合色素含量、光合参数、荧光动力学参数的测定。

a.63第三节位叶片;b.63第九节位叶片;c.Liu-2第三节位叶片;d.Liu-2第九节位叶片;e.63植株生长形态,其中Ⅰ、Ⅱ分别表示63植株第三节位和第九节位叶片。比例尺=1 cma.63 3rd node;b.63 9th node;c.Liu-2 3rd node;d.Liu-2 9th node.e.63 Plant growth pattern,Ⅰ and Ⅱ indicate the 3rd node leaf and 9th node leaf of 63 plant.Scale:1 cm
63子叶出土时呈淡黄色,随后萌生的真叶也呈淡黄色,第二片真叶展平后,原本黄化的子叶整体转绿。随着植株生长发育,后续萌生出的所有新叶均呈淡黄色,但随着叶片生长发育自叶缘向内转绿,这种后绿的特征伴随该品系的整个生长期,并且该突变体能够正常结实。2019和2020年其果实品质性状见表1。

表1 后绿突变西瓜63果实的相关性状Table 1 Fruit related traits of virescent mutant 63
1.2 光合色素含量测定
用乙醇提取法[20]进行叶绿素提取。在63和Liu-2的Ⅰ、Ⅱ两时期各取10片叶片为1组(共4组),将每组叶片剪碎混匀,称取0.2 g加入10 mL体积分数95%的乙醇避光浸泡48 h,期间进行多次振荡,重复3次。
采用可见光分光光度计测量663,645 和470 nm波长的吸光值并计算叶绿素a(Chl a)、叶绿素b(Chl b)、总叶绿素(Chl)和类胡萝卜素(Caro)含量,计算公式为:
Chl a=(12.7A663 nm-2.69A645 nm)×V/(W×1 000);
Chl b=(22.9A645 nm-4.68A663 nm)×V/(W×1 000);
Caro=(1 000A470 nm-3.27 Chl a-104 Chl b)×V/(229×1 000×W);
Chl=Chl a+Chl b。
式中:Chl a、Chl b、Caro、Chl分别表示叶绿素a、叶绿素b、类胡萝卜素、总叶绿素含量(mg/g),A663nm、A645nm、A470nm分别表示相应波长下的OD值,V表示提取液体积(mL),W表示叶片质量(g)。
1.3 光合参数测定
使用Li-6400测定光合参数。选择晴朗上午测定63和Liu-2 Ⅰ、Ⅱ两时期的净光合速率(Pn)、气孔导度(Gs)、胞间CO2浓度(Ci)、蒸腾速率(Tr),重复测定3次取平均值。
1.4 叶绿素荧光动力学参数测定
使用Li-6400测定荧光动力学参数。对63和Liu-2 Ⅰ、Ⅱ两个时期的叶片进行标记,并使用锡箔纸严密包裹,经过30 min以上暗处理后,测定初始荧光产量(F0)、最大荧光产量(Fm)及最大光化学效率(Fv/Fm,Fv=Fm-F0),随后对标记叶片进行30 min以上的光活化,测定或计算光下最小荧光产量(F0′)、光下最大荧光产量(Fm′)、实际光化学效率(Fv′/Fm′,Fv′=Fm′-F0′)、实际光化学量子效率(ΦPSⅡ)、光化学淬灭系数(qP)和电子传递效率(ETR)。重复3次取平均值。
1.5 数据分析
使用Execl表格进行数据整理,并使用SPSS工具进行数据分析,使用One-way ANOVA进行数据间差异显著性分析。
2 结果与分析
2.1 后绿突变体63光合色素含量的变化
由表2可知,在西瓜叶片的Ⅰ和Ⅱ时期,正常绿叶品系Liu-2叶片中的Chl a、Chl b、Caro、Chl及Chl a/Chl b差异不显著,Caro/Chl显著降低了2.7%;而后绿突变体63叶片中的Chl a、Chl b、Caro及Chl在两个时期间均存在极显著差异,Ⅱ时期较Ⅰ时期分别增加177.7%,251.9%,92.0%和171.1%,而Caro/Chl极显著降低了29.2%。可见光合色素含量在正常绿叶品系的幼叶和功能叶之间无明显差异,而在后绿突变体中却有极显著变化。

表2 Ⅰ和Ⅱ时期正常绿叶品系Liu-2及后绿突变西瓜63生长过程中光合色素的变化Table 2 Changes in photosynthetic pigments during growth of normal green leaf inbred line Liu-2 and virescent mutant watermelon 63 in periods Ⅰ and Ⅱ
2.2 后绿突变体63光合参数的变化
由表3可知,与Ⅰ时期相比,正常绿叶品系Liu-2 Ⅱ时期叶片的Pn增加了133.1%;差异达极显著水平;Ci含量降低16.0%,差异达显著水平;Gs增加41.0%,差异也达显著水平;Tr增加7.3%,差异不显著。说明对正常绿叶品系而言,西瓜幼叶至功能叶的转变,能够在一定程度上提高叶片的光合能力。表3表明,与Ⅰ时期相比,后绿突变西瓜63 Ⅱ时期叶片的Pn、Gs和Tr分别增加了1 213.7%,49.6%和17.3%,差异均达极显著或显著水平;而Ci含量降低19.3%,二者间也存在极显著差异。上述结果说明,在Ⅱ时期,正常绿叶品系Liu-2和后绿突变体63的功能叶较幼叶光合能力均有所提升,但后绿突变体63的Pn提升了1 213.7%,正常绿叶植株增加相对较小。

表3 Ⅰ和Ⅱ时期正常绿叶品系Liu-2及后绿突变西瓜63光合特性的变化Table 3 Changes in photosynthetic characteristics of green leaf inbred line Liu-2 and virescent mutant watermelon 63 in periods Ⅰ and Ⅱ
2.3 后绿突变体63荧光动力学参数的变化
由表4可知,与Ⅰ时期相比,Ⅱ时期正常绿叶品系Liu-2的F0减小了5.2%,二者差异不显著,而Fm、Fv/Fm、Fv′/Fm′、ΦPSⅡ、qP、ETR分别增加了8.0%,3.1%,34.0%,77.2%,32.4%和77.2%,差异均达到极显著水平。与Ⅰ时期相比,Ⅱ时期后绿突变西瓜63的F0、Fm分别增加78.1%和124.1%,差异均达到极显著水平,且增加幅度均明显大于Liu-2,同时Fv/Fm、Fv′/Fm′、ΦPSⅡ、qP、ETR较转绿前分别增加6.6%,25.3%,70.0%,34.5%和68.5%,差异也均达到极显著水平。
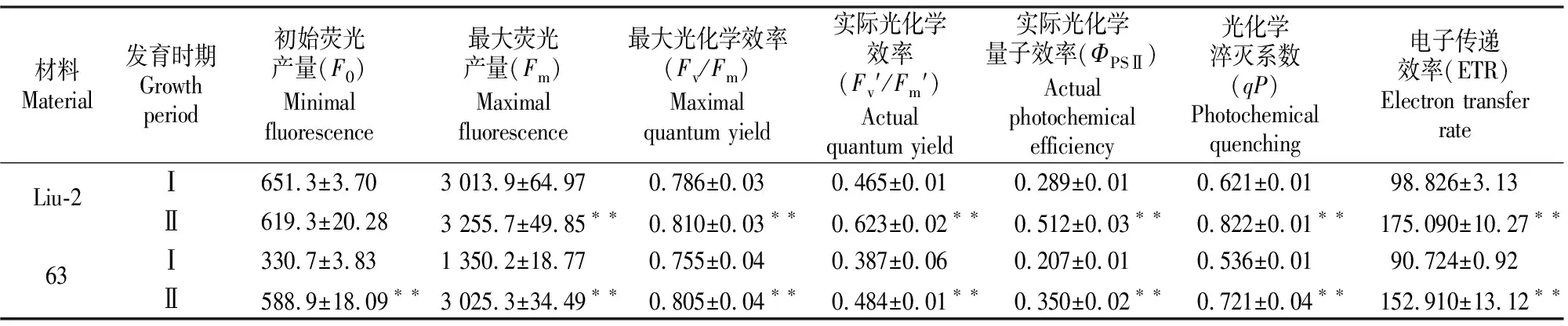
表4 Ⅰ和Ⅱ时期正常绿叶品系Liu-2及后绿突变西瓜63荧光动力学参数的变化Table 4 Changes in chlorophyll fluorescence parameters of green leaf inbred line Liu-2 and virescent mutant watermelon 63 in periods Ⅰ and Ⅱ
3 讨 论
实际观察发现,后绿突变体63子叶出土后呈淡黄色,第一片真叶也呈淡黄色,当第二片真叶展平后,子叶整体转绿。随着植株生长发育,后续萌生出的所有新叶均呈淡黄色,这种后绿的特征伴随该品系的整个生长期。目前在西瓜作物中,关于叶色突变体的报道较少。任艺慈等[10]发现一株西瓜黄化突变体YL,该突变体叶色黄化特性伴随其整个生长期。王凤辰等[14]发现一株“芽黄”西瓜突变体,该突变体嫩芽为淡黄色,第一片真叶呈淡黄色,随叶片生长发育自叶柄至全叶由黄转绿,当第二片真叶展平时,第一片真叶已经基本转绿。而与其相比,本试验的后绿突变体63的转绿速度较慢。Haileslassie[17]报道了一个西瓜后绿突变体“后绿”,该突变株叶型较后绿突变体63宽大,同时萌生出的新叶呈现淡绿色,随着叶片生长发育,叶片逐渐完全转绿。而后绿突变体63转绿后叶片中的Chl及Caro含量仍低于对照植株。综上所述,63是一株新发现的西瓜后绿突变体。
叶绿素是植物光合作用中的重要色素,叶绿素可以捕捉光能并转化为化学能[21]。Haileslassie[17]研究表明,西瓜后绿突变体“后绿”新叶的光合色素缺失,随着植株生长发育光合色素快速积累,完全转绿后叶片中的光合色素含量与对照正常绿叶品系一致。而63新叶光合色素大量缺失,转绿后其Chl、Caro逐渐提高,但仍低于对照正常绿叶品系Liu-2。该结果表明,西瓜后绿突变体63的转色机制不同于突变体“后绿”。大部分后绿突变体转绿前,叶片中的Chl a/Chl b值低于转绿后的Chl a/Chl b值,该类型后绿突变体转绿前表现为Chl、Caro大量缺失,即为“总叶绿素缺失型”[7,11,17,22]。另一种后绿突变体转绿前叶片中的Chl a/Chl b值高于转绿后的Chl a/Chl b值,该类型后绿突变体是由于Chl a向Chl b转化的过程中受阻,造成Chl b缺失,从而导致Chl缺失,即为“叶绿素b缺失型”[23]。后绿突变体63转绿前光合色素含量大量缺失,符合“总叶绿素缺失型”的描述。正常绿叶品系叶片中的Caro/Chl约为1∶3,本研究表明,正常绿叶品系Liu-2 Ⅰ时期和Ⅱ时期的Caro/Chl分别为0.297和0.289,接近为1∶3,与前人研究结果[24]一致;后绿突变体63转绿前的Caro/Chl为0.431,故叶片呈现淡黄色,转绿后Chl a、Chl b的积累速度快于Caro的积累速度,完全转绿后Caro/Chl为0.305,故叶色开始呈现绿色。
光合色素含量直接或间接影响着植物的光合能力,随着后绿突变体63转绿,叶片中的光合色素含量恢复,Pn提高,Ci降低,这也表明随着光合色素的累积,后绿突变体63光合能力可以恢复,该结果与苗晗[25]的研究结果一致。后绿突变体63的Gs及Tr的提高程度大于正常绿叶品系,可能是气孔部分关闭和光合色素缺失共同导致了植株的光合能力减弱,该规律与杨佳秀等[26]的研究结果一致。
F0为光系统Ⅱ的初始荧光产量,Fm为光系统Ⅱ中通过的最大电子数量,F0和Fm与光合色素含量相关;Fv/Fm值可以反映光系统Ⅱ中光能的最大转化效率,Fv′/Fm′则反映了光系统Ⅱ原初光能的转化效率[27],ΦPSⅡ表示光系统Ⅱ实际的光化学量子效率,qP表示光化学淬灭,ETR表示电子传递速率[28]。后绿突变体63 Ⅰ时期荧光动力学参数的测定表明,其F0、Fm均小于同时期正常绿叶品系Liu-2,说明63转绿前对光能的捕捉能力较弱,从而导致63的净光合速率低于同时期的正常绿叶品系,这与林秋云等[29]的研究结果相一致。后绿突变西瓜63及正常绿叶品系Liu-2两品系Fv/Fm、Fv′/Fm′、ΦPSⅡ、qP、ETR的变化程度比较接近,说明后绿突变西瓜63及正常绿叶品系Liu-2的光化学效率、光能实际利用率、光能转换率和电子转递效率逐渐提高,是叶片发育由幼叶发育为功能叶所致,与其光合色素含量的增加并不相关。因此笔者推测,后绿突变体63转绿后,由于叶片光合色素含量的提高,其对光的捕捉能力和荧光产量也随之提高,这是净光合速率提高的主要原因。
猜你喜欢
作物学报(2022年2期)2022-11-06
湖南农业科学(2022年1期)2022-06-06
作物学报(2022年8期)2022-05-29
蔬菜(2021年7期)2021-11-27
广西蚕业(2021年3期)2021-10-28
语文世界(小学版)(2021年5期)2021-06-04
当代水产(2021年1期)2021-03-19
山东农业科学(2018年3期)2018-03-29
山东农业科学(2018年11期)2018-01-17
安徽农学通报(2017年24期)2018-01-12
