
Antibiotics, gut microbiota, and irritable bowel syndrome: What are the relations?
2022-03-31 08:04ZarinaMamievaElenaPoluektovaValerySvistushkinVasilySobolevOlegShifrinFranciscoGuarnerVladimirIvashkin
World Journal of Gastroenterology 2022年12期
Zarina Mamieva, Elena Poluektova, Valery Svistushkin, Vasily Sobolev, Oleg Shifrin, Francisco Guarner,Vladimir Ivashkin
Abstract Irritable bowel syndrome (IBS) is a functional gastrointestinal disorder in which recurrent abdominal pain is associated with defecation or a change in bowel habits (constipation, diarrhea, or both), and it is often accompanied by symptoms of abdominal bloating and distension. IBS is an important health care issue because it negatively affects the quality of life of patients and places a considerable financial burden on health care systems. Despite extensive research, the etiology and underlying pathophysiology of IBS remain incompletely understood.Proposed mechanisms involved in its pathogenesis include increased intestinal permeability, changes in the immune system, visceral hypersensitivity, impaired gut motility, and emotional disorders. Recently, accumulating evidence has highlighted the important role of the gut microbiota in the development of IBS.Microbial dysbiosis within the gut is thought to contribute to all aspects of its multifactorial pathogenesis. The last few decades have also seen an increasing interest in the impact of antibiotics on the gut microbiota. Moreover, antibiotics have been suggested to play a role in the development of IBS. Extensive research has established that antibacterial therapy induces remarkable shifts in the bacterial community composition that are quite similar to those observed in IBS.This suggestion is further supported by data from cohort and case-control studies,indicating that antibiotic treatment is associated with an increased risk of IBS. This paper summarizes the main findings on this issue and contributes to a deeper understanding of the link between antibiotic use and the development of IBS.
Key Words: Gut microbiota; Irritable bowel syndrome; Antibiotics; Intestinal barrier; Gut motility; Gut sensitivity
INTRODUCTION
Recent advances in culture-independent techniques have greatly expanded our understanding of the human gut microbiota and its functionalities. It is becoming increasingly recognized that gut bacteria play a pivotal role in host homeostasis and are involved in the progression and development of numerous human diseases.
The gut microbiota is established early in life, remains relatively stable thereafter, and is subject to shaping by environmental and host factors (e.g.,age, diet, lifestyle, and medications)[1 ,2 ]. With regard to the environment, antibiotics have been reported to play a particularly important role in the modulation of the gut microbial community. However, most studies in this area were undertaken 30 to 40 years ago and relied on culture-based techniques. Global antibiotic use has grown 66 % since 2000 and continues to grow at a high rate[3 ,4 ]. This fact, along with rapid technological advancements for cultureindependent analysis, has reinforced the need to take a fresh look at antibiotic-induced changes in the human gut microbiota and clinical consequences of antibiotic intervention. Several studies have reported that antibiotic treatment is associated with an increased risk of irritable bowel syndrome (IBS)[5 -8 ].
IBS is a common gastrointestinal disorder affecting 10 %-15 % of the population in Europe and North America[9 ]. This condition negatively affects the quality of life of patients and imposes a significant socioeconomic burden[10 ]. Over the past few decades, the gut microbiota has emerged as a potential factor that contributes to the pathophysiology of IBS[11 ,12 ]. Microbial dysbiosis within the gut has been implicated in intestinal barrier dysfunction, visceral hypersensitivity, impaired gastrointestinal motility,and altered immune response[13 -17 ]. Moreover, various studies have consistently shown the efficacy of microbiota-directed therapies, including prebiotics, probiotics, nonabsorbable antibiotics, dietary changes, and fecal microbial transplantation, in alleviating IBS symptoms[18 ].
In this paper, we provide a brief overview of the human gut microbiota and its impact on host homeostasis. We highlight what is currently known regarding the role of gut bacteria in the pathophysiology of IBS. Furthermore, we provide an overview of the most up-to-date literature about the impact of antibiotics on gut microbiota composition and discuss a possible link between antibiotic use and the development of IBS. Finally, we identify knowledge gaps and uncertainties that must be filled to orient future research in this area.
GUT MICROBIOTA AND ITS ROLE IN HOST HOMEOSTASIS
The human gut microbiota is a community of microorganisms that inhabit the gastrointestinal tract and is composed of approximately 1014 bacterial cells[19 ,20 ]. In healthy adults, more than 90 % of gut bacteria belong to four dominant phyla, namely,Firmicutes, Bacteroidetes, Actinobacteria,andProteobacteria,whereas other phyla are far less abundant[21 ,22 ].
Currently, the gut microbiota is considered an indispensable “organ” within the body with distinct metabolic and immune functions (Table 1 ). Most of its effects are mediated through metabolites.
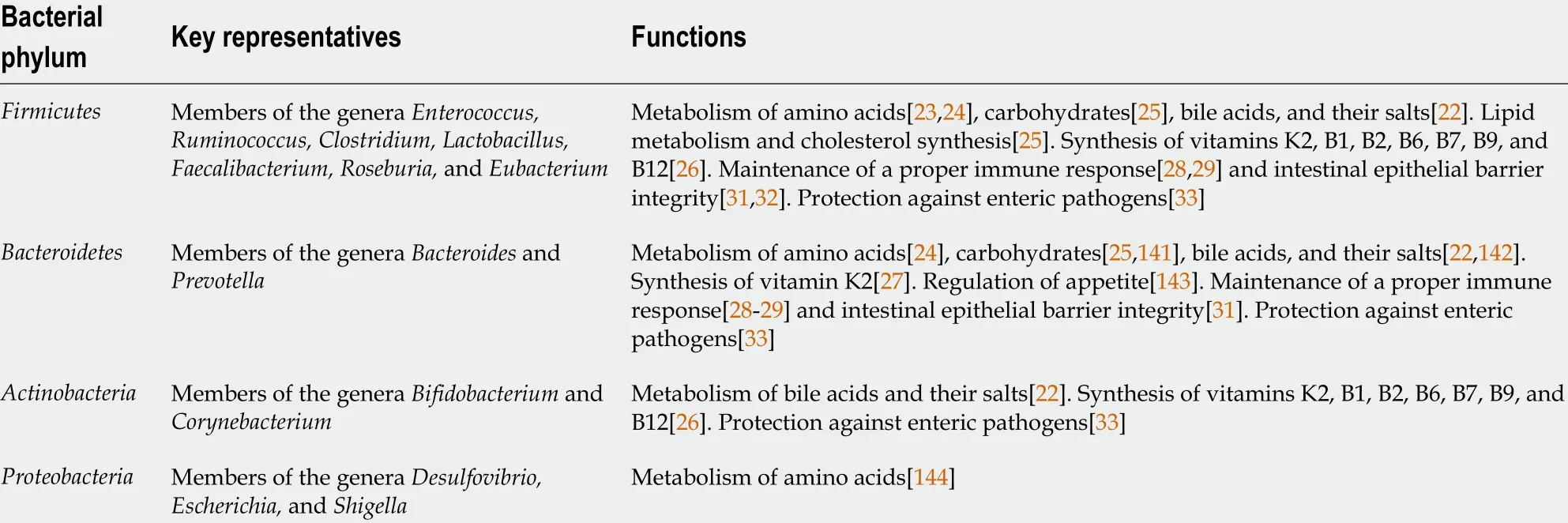
Table 1 Gut microbiota functions
Thus, some of the most important roles of the gut microbiota include metabolism of dietary compounds[23 -25 ], synthesis of vitamins[26 ,27 ], regulation of the immune response[28 -30 ], maintenance of intestinal epithelial barrier integrity[25 ,31 ,32 ], and protection against enteric pathogens[33 ].
MODERN CONCEPT OF IBS: EVOLVING ROLE OF GUT MICROBIOME
Despite extensive research, the etiology and underlying pathophysiology of IBS remain incompletely understood. Proposed mechanisms involved in its pathogenesis include visceral hypersensitivity,impaired gut motility[13 ,34 ], increased intestinal permeability[34 -36 ], emotional disorders[11 ,37 ], and changes in the immune system[34 ,37 ,38 ].
Over the past decade, there has been an increasing amount of literature on the role of the gut microbiota in the pathogenesis of IBS. The concept of the “microbiota-gut-brain” axis has been proposed[14 -17 ], supporting the crucial role of microbial dysbiosis in the development of IBS symptoms. It is thought that, in genetically predisposed individuals, environmental factors alter the composition of the gut microbiota, leading to disruption of intestinal epithelial barrier integrity[13 ]. Once the intestinal barrier is breached, bacteria interact with the immune system of the host, provoke a series of immune reactions, and lead to low-grade mucosal inflammation in the gut wall. Collectively, these changes result in sensitivity and motility abnormalities, emotional disorders, and the development of IBS symptoms (abdominal pain, bloating, and alterations in bowel habits)[35 ]. Interestingly, the gut microbiota not only initiates such a pathological cascade in IBS but also contributes to all aspects of its multifactorial pathogenesis through the release of metabolites[11 ,12 ]. These provisions will be discussed below.
Microbiota and motility/sensitivity abnormalities
The enteroendocrine system modulates gut motor and sensory functions through the secretion of neuropeptides and neurotransmitters[39 ].
Bacterial metabolites are able to stimulate the production of several neuropeptides, including neuropeptide Y, peptide YY, glucagon-like peptide-1 (GLP-1 )[40 ], cholecystokinin, and substance P(Figure 1 )[15 ,41 ].
For instance, short-chain fatty acids (SCFAs), secondary bile acids, and indole, which are produced by members of the generaClostridium, Bacteroides,andRuminococcus[23 ,25 ], stimulate intestinal L-cells to secrete GLP-1 [42 ]. GLP-1 reduces postprandial motility in the upper gastrointestinal tract (antrum,duodenum, and jejunum) and increases colonic transit[43 ,44 ]. A study conducted by Li et al[45 ] reported decreased serum GLP-1 levels and reduced mucosal expression of GLP-1 receptors in patients with constipation-predominant IBS (IBS-C). The authors suggested that lower GLP-1 levels lead to the loss of its prokinetic effects in the colon, resulting in constipation and abdominal pain. In a rat model of bowel dysfunction, administration of the GLP-1 receptor agonist exendin-4 alleviated stress-induced defecation and visceral pain sensitivity[46 ,47 ]. Clinical interventions in patients with IBS demonstrated that the synthetic GLP-1 analog ROSE-010 reduced abdominal pain and increased colonic transit[45 ,48 ].The underlying molecular mechanisms responsible for the amelioration of symptoms remain unknown.The authors suggest that modulation of enteric neuronal function and tight junction expression, as well as the activation of serotonergic pathways in the colon, may play a role.
Secondary bile acids and SCFAs, which are mainly produced byEubacterium, Bacteroides, andClostridium(clusters IV, XI, XIII, and XIVa)[22 ], promote serotonin synthesis from colonic enterochromaffin cells[49 ]. Serotonin is an important neurotransmitter that, among its other functions, regulates gastrointestinal motility[50 ]. Serum serotonin levels were found to be increased in those with diarrheapredominant IBS (IBS-D) and reduced in those with IBS-C[34 ].
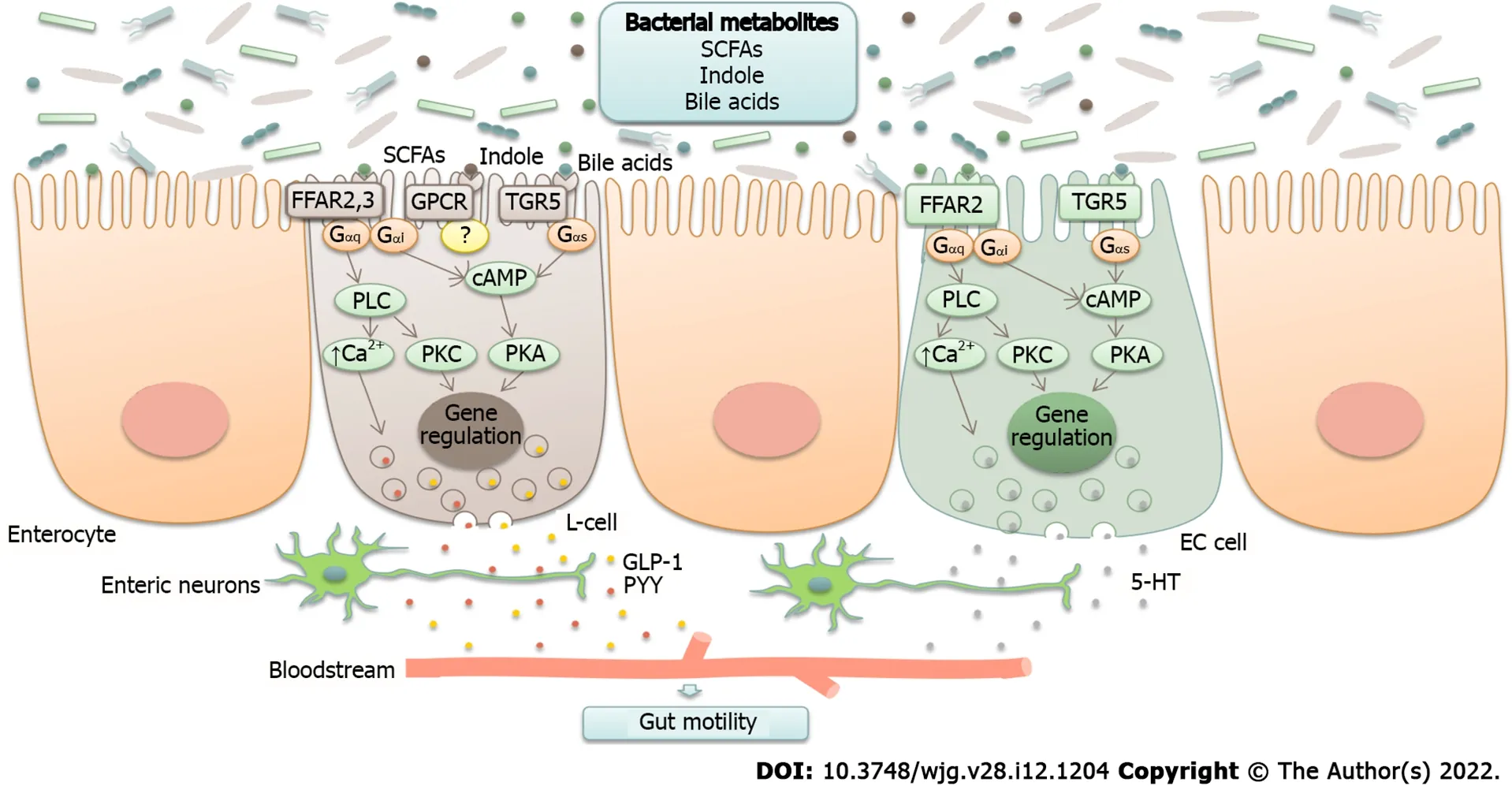
Figure 1 Neurotransmitter modulation by gut microbiota (schematic illustration). Bacterial metabolites, such as short-chain fatty acids (SCFAs),secondary bile acids, and indole, are able to stimulate the production of neurotransmitters, including glucagon-like peptide-1 (GLP-1 ), peptide YY (PYY), and serotonin (5 -HT). They act through G-protein coupled receptors (GPCRs) FFAR2 , FFAR3 , and TGR5 that are coupled to different types of G proteins (Gαs, Gαq, and Gαi) and activate different pathways known to regulate gene expression and promote exocytosis by raising intracellular Ca2 + levels. SCFAs are recognized by FFAR2 and FFAR3 . Enteroendocrine L-cells express both of these proteins, whereas enterochromaffin (EC) cells have been reported to express FFAR2 . Bile acids are recognized by TGR5 receptors expressed in L-cells and EC cells. The sensing of indole remains elusive, although it is thought to act through GPCR. Gαs stimulates adenylate cyclase and elevates cyclic adenosine monophosphate (cAMP), which activates protein kinase A (PKA). Gαi inhibits the cAMP pathway. Gαq stimulates phospholipase C (PLC), resulting in the generation of diacylglycerol (DAG) and inositol triphosphate (IP3 ), which activate protein kinase C (PKC) and induce intracellular Ca2 + release[23 ,138 -140 ]. SCFA: Short-chain fatty acids; GLP-1 : Glucagon-like peptide-1 ; PYY: Peptide YY.
The serotonin system represents a potential therapeutic target in IBS. The effects of serotonin are mediated through 5 -HT receptors located on the surface of distinct cell types. Fourteen different serotonin receptor subtypes have been identified and classified into seven groups (5 -HT1 -7 ), with 5 -HT3 and 5 -HT4 being the most investigated receptors in the intestine. Both receptor subtypes are expressed on neurons within the myenteric and submucosal plexuses of the enteric nervous system, intrinsic and extrinsic sensory neurons, interstitial cells of Cajal, enterocytes, and enterochromaffin cells[51 ]. 5 -HT3 receptors are involved in the contraction of intestinal smooth muscle and in gut-brain communication through vagal afferent fibers[52 ]. Activation of 5 -HT4 receptors induces neuronal release of acetylcholine and accelerates the peristaltic reflex[53 ]. 5 -HT3 receptor antagonists have been shown to improve abdominal pain and global IBS symptoms in patients with nonconstipated and IBS-D[54 ,55 ]. 5 -HT4 agonists have been shown to relieve overall and individual symptoms (abdominal pain/discomfort,stool frequency, stool consistency, and straining during defecation) in patients with IBS-C[56 -58 ].However, cardiovascular side effects were seen with these drugs, and they were either withdrawn from the market (cisapride) or approved for a limited population (tegaserod). Therefore, new safe and welltolerated 5 -HT4 agonists are under development[59 ,60 ].
A number of animal studies have shown the prominent role of the gut microbiota in visceral hypersensitivity[41 ]. For example, colonization of germ-free rats with the gut microbiota from patients with IBS reduced the pain threshold to colorectal distension[42 ]. Furthermore, the beneficial effects of probiotic strains (e.g., Lactobacillus reuteri,Lactobacillus plantarum, Lactobacillus helveticus,andBifidobacterium longum) in alleviating visceral sensitivity have been documented[61 -63 ].
Thus, the microbiota influences the main pathogenetic factors of IBS (i.e.,motility and sensitivity)both directly and through microbial metabolites.
Microbiota as a regulator of stress and emotional responses
The physiological response to stress is mediated through the hypothalamic-pituitary-adrenal (HPA) axis[64 ]. Activation of this axis results in the release of corticotropin releasing hormone (CRH) from the paraventricular nucleus of the hypothalamus. CRH acts on the anterior pituitary and induces the production of adrenocorticotropic hormone (ACTH), which in turn stimulates the adrenal cortex to secrete cortisol.
Different types of stressors are known to contribute to the development, maintenance, and exacerbation of IBS symptoms[11 ]. The results of multiple studies suggest that there is HPA axis dysregulation in IBS. For instance, patients with IBS were found to have excess levels of ACTH in the plasma and cortisol in the serum in response to CRH infusion[65 ].
Growing evidence indicates that the gut microbiota is involved in the regulation of HPA axis activity.It has been shown that colonization with beneficial microorganisms in early life is of great importance for the normal development of this axis[66 ]. Moreover, alterations in the gut microbiota may influence the release of ghrelin and galanin, which are endocrine peptides contributing to the stress response through modulation of CRH, ACTH, and glucocorticoid secretion[40 ,67 ].
Dysfunction of the HPA axis, along with alterations in neurotransmitter metabolism, appear to be crucial factors in the development of psychiatric disorders, such as anxiety and depression[68 ,69 ]. A recent meta-analysis of 27 studies have reported elevated levels of anxiety and depression in patients with IBS as compared to those in healthy controls[70 ]. Comorbid emotional disorders lead to persistence of symptoms, drive patients to seek medical care, and contribute to poor outcomes[11 ].
A growing body of literature supports the association between microbial dysbiosis and the development of anxiety and depression. For instance, certain species within theLactobacillaceaeandBifidobacteriaceaefamilies are known to produce gamma-aminobutyric acid (GABA). GABA is the main inhibitory neurotransmitter of the central nervous system, playing an important role in the pathogenesis of mood disorders[49 ,71 ]. Interestingly, a specific type of GABA receptor (GABA-b) is localized on submucosal and myenteric neurons of the enteric nervous system[72 ] and is thought to be involved in the modulation of gut motility and sensitivity[37 ]. Furthermore, members of the generaBacillusandEscherichiahave been found to produce other neurotransmitters affecting mood and behavior, such as dopamine, serotonin, and norepinephrine[15 ,73 ]. In recent studies, germ-free mice have been widely used as a tool for assessing the role of intestinal microbes in brain function and behavior. Studies on germ-free and specific pathogen-free mice indicate that intestinal microbes can cause imbalances of the HPA axis, resulting in an anxiety-like behavioral phenotype[74 ]. Fecal microbiota transplantation studies have indicated the rodent-to-rodent and human-to-rodent transfer of anxiety-like behaviors[75 ,76 ]. Moreover, animal studies have shown that transplantation of the microbiota from depressed patients to rodents is able to induce depression-like behavior. The authors linked microbiota-induced depression in mice to alterations in the cAMP-response element binding protein (CREB) signaling pathway in the olfactory bulb[77 ] and alterations in carbohydrate and amino acid metabolism[78 ].
However, despite the data obtained, further research is needed to investigate the difference in emotional disorder levels in patients with postinfectious and other forms of IBS.
Microbiota and host immunity
Recently, considerable literature has grown around the theme of immune system activation in IBS. For instance, an increased number of mast cells located in close proximity to enteric nerve fibers have been found in colonic biopsies from patients with IBS and have been associated with the severity of symptoms[11 ,38 ,79 ]. Mast cells are thought to be key players in intestinal mucosal inflammation[79 ].Their degranulation causes the release of inflammatory mediators (histamine, serotonin, and proteases),resulting in lymphocyte activation and cytokine imbalance[80 ]. Patients with IBS were found to have higher levels of proinflammatory interleukin (IL)-6 , IL-8 , IL-1 β, and tumor necrosis factor-α (TNF-α) and lower levels of anti-inflammatory IL-10 in both serum and the intestinal mucosa[81 ,82 ]. These changes result in altered pain thresholds and visceral hypersensitivity[38 ,83 ]. In addition, mast cell degranulation has been shown to reduce the expression of tight junction proteins, probably through tryptase release[13 ]. Apart from mast cells, increased numbers of eosinophils and intraepithelial lymphocytes have been observed in colonic biopsies from patients with IBS[11 ,79 ].
Gut bacteria play an important role in the modulation of the immune response. For example, butyrate produced by members of the phylumFirmicutes[25 ] induces the differentiation of regulatory T cells[29 ,84 ], thereby preventing an excessive immune response and autoimmunity[22 ,85 ]. Furthermore,Lactobacillispp. metabolize dietary tryptophan into indole-3 -aldehyde, which acts as an aryl hydrocarbon receptor (AHR) ligand[85 ]. AHR is a ligand-activated transcription factor that is expressed by immune cells and regulates the number of intraepithelial lymphocytes and IL-22 production[86 ].Probiotic strains, such asLactobacillus rhamnosus,Lactobacillus casei,andBifidobacterium breve,were shown to induce IL-4 and IL-10 production, whereasL. reuteriandL. plantarumwere found to downregulate the expression of TNF-α[87 ,88 ].
The importance of the interaction between the gut microbiota and host immune system in IBS is highlighted by a number of studies in patients with postinfectious IBS, indicating activation of the gastrointestinal immune system after acute gastroenteritis[89 ,90 ]. Moreover, animal studies have shown that stress-induced changes in the gut microbiota are associated with altered immune response and increased susceptibility to enteric pathogens[91 ,92 ].
Microbiota and intestinal barrier integrity
Intestinal epithelial barrier integrity is of great importance for gut homeostasis, as it prevents the translocation of luminal antigens to the mucosa, thus averting the development of low-grade mucosal inflammation in the gut wall (Figure 2 ).
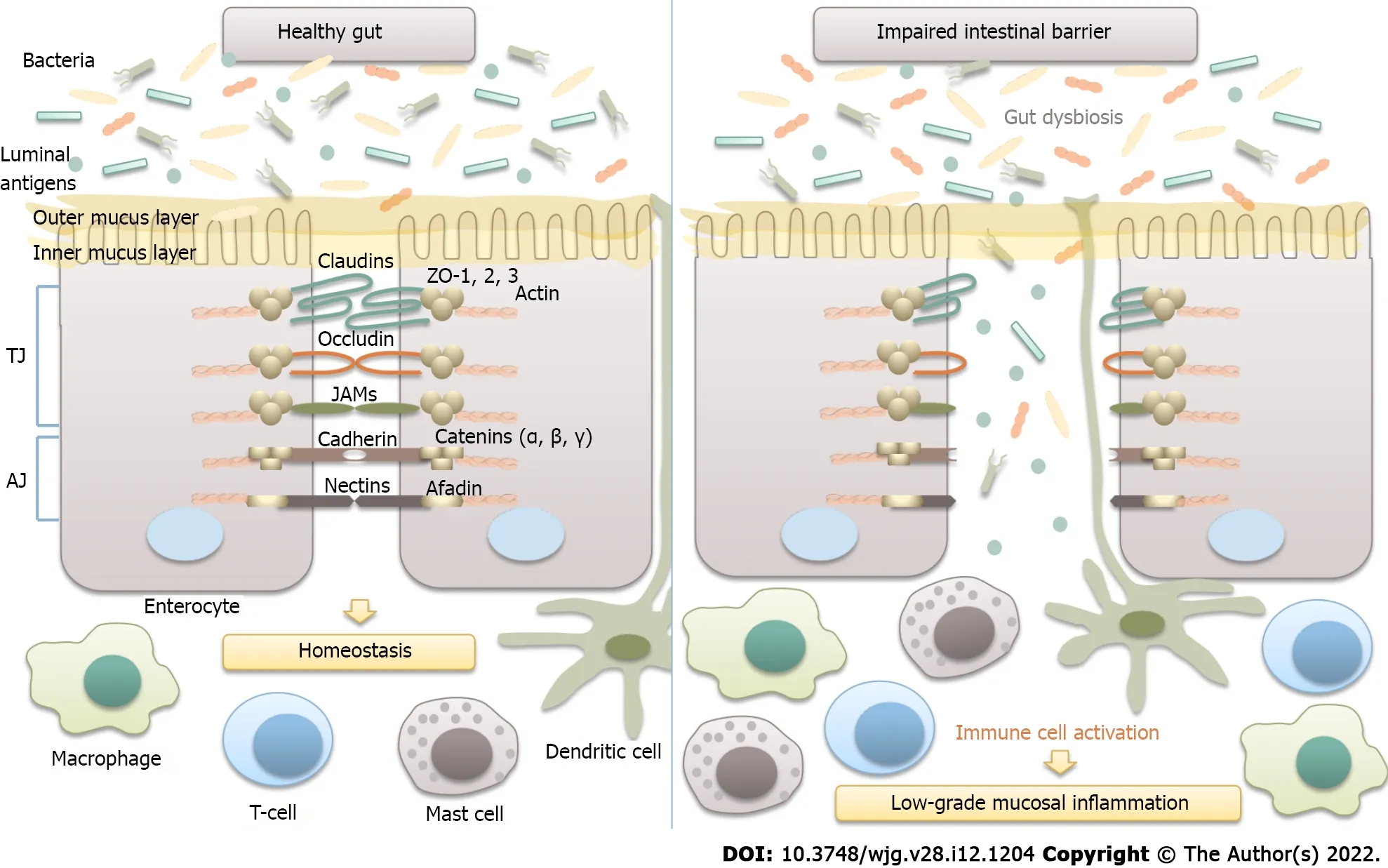
Figure 2 Microbiota and intestinal barrier integrity. The intestinal barrier plays an essential role in maintaining host homeostasis. It is mainly composed of the mucus layer, the epithelial layer, and the underlying lamina propria. Intestinal epithelial cells are tightly attached to each other by junctional complexes. Tight junctions (TJs) are composed of several proteins, including occludin, claudins, zonula occludens (ZOs), and junctional adhesion molecules (JAMs), which interact with each other, as well as with the cytoskeleton. The adherence junction is composed of the nectin-afadin system and the E-cadherin-catenin system. Intestinal epithelial barrier integrity prevents the translocation of bacteria and luminal antigens to the mucosa, thus averting their interaction with the host immune system and the development of low-grade mucosal inflammation in the gut wall. TJ: Tight junctions; AJ: Adherence junction; JAM: Junctional adhesion molecules.
An increased density of epithelial gaps has been shown by electron microscopy in gut biopsies of patients with IBS[93 ]. Furthermore, histological examination of colonic biopsies revealed decreased expression of tight junction proteins, such as occludin; claudins 1 , 3 , and 5 ; and zonula occludens-1 [13 ,36 ,82 ,93 ]. Increased serum levels of anti-flagellin antibodies in patients with IBS further support the substantial role of intestinal permeability in the pathogenesis of IBS[94 ].
The gut microbiota is an important determinant of intestinal epithelial barrier integrity. In particular,certain gut bacteria, such asBacteroides thetaiotaomicron, Faecalibacterium prausnitzii,andRuminococcusspp., were shown to affect the mucus layer thickness and composition[1 ,22 ,31 ]. Moreover, SCFAs, which are produced predominantly by members of the generaEubacterium, Clostridium, Ruminococcus,andFaecalibacterium,have been demonstrated to augment the expression of claudins 3 and 4 and occludin.Polyamines (putrescine, spermidine, and spermine), which are produced by certain species within the
Clostridium, Enterococcus, Streptococcus,andLactobacillusgenera,have been shown to stimulate the production of E-cadherin and zonula occludens-1 [95 ]. There is also evidence that probiotic strains ofBifidobacteriumandLactobacilluspromote intestinal barrier function and prevent bacterial translocation[32 ,96 ].
Most likely, the preservation of the optimal composition of the microbiota (e.g.,a sufficient number of SCFA producers) may serve as a factor preventing the development of IBS.
GUT MICROBIAL COMPOSITION IN PATIENTS WITH IBS
A considerable amount of literature has been published on the compositional changes of the gut microbiota in patients with IBS. Although data from these studies are inconsistent and even conflicting,some common features can be found (Table 2 ). The discrepancy in findings is possibly due to differences in the population studied (e.g.,age, lifestyle, initial microbiota composition, prior antibiotic and/or probiotic use, and diagnostic criteria for IBS) and methodological issues, such as study design and methods for microbiota assessment and data analysis.
The majority of authors report decreased microbial diversity in patients with IBS[97 -101 ].Furthermore, a substantial number of studies have shown a lower abundance of butyrate-producingbacteria from the genusFaecalibacterium, mainlyF. prausnitzii,[97 ,98 ,102 ,103 ] as well as an increase in the abundance of theEnterobacteriaceaefamily, including pathogens such asEscherichia coliandEnterobacterspp.[98 ,104 -106 ]. Moreover, patients with IBS were found to have a reduced prevalence ofBifidobacterium,providing a range of beneficial properties to the host[98 ,103 ,104 ,106 ,107 ]. Significant differences inLactobacillusnumbers were also observed between patients with IBS and healthy controls, but the findings of different studies were not consistent. Some authors reported an increased amount ofLactobacillus[98 ,99 ,102 ,108 ], while others documented a decrease in the abundance of this commensal[103 ,104 ,106 ,107 ,109 ].
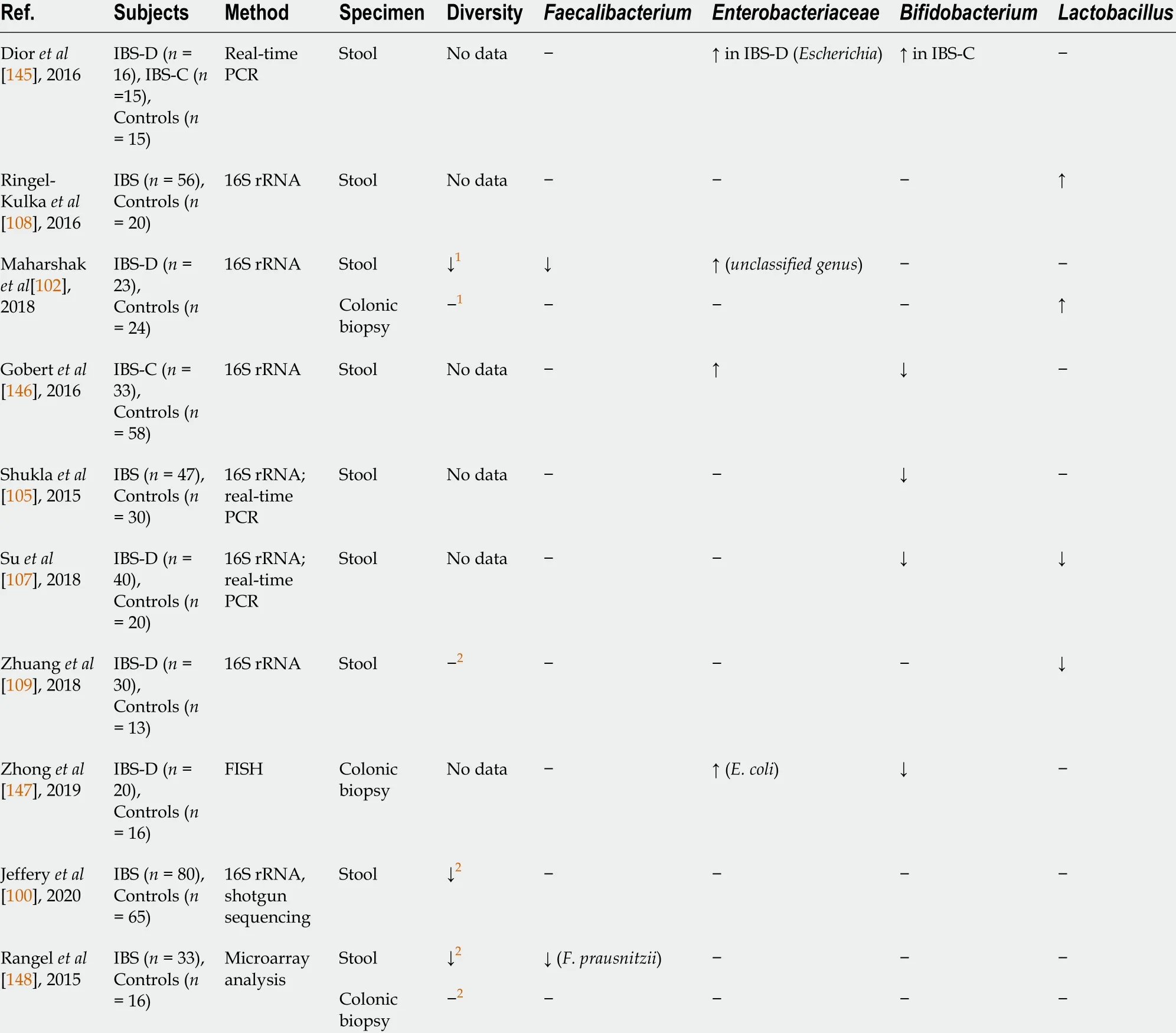
Table 2 Compositional changes in gut microbiota in patients with irritable bowel syndrome (common threads)
Overall, there seems to be some evidence to indicate that patients with IBS have decreased numbers of bacteria contributing to the maintenance of host homeostasis and proper immune response, as well as increased numbers of microbes with proinflammatory properties.
ANTIBIOTICS, GUT MICROBIOTA, AND IBS
Effects of antibiotics on gut microbiota composition
The discovery of antibiotics in the early 20thcentury was a great milestone in the history of medicine, as it changed the natural course of most infectious diseases and saved countless lives[110 ,111 ]. However, a growing number of studies have shown that inappropriate use of antibiotics promotes the development of antibiotic resistance[112 ,113 ]. Furthermore, accumulating evidence indicates that antibiotic exposure in early life increases the risk of obesity and autoimmune and allergic diseases[114 -117 ].
During the past four decades, there has been an increasing interest in the impact of antibiotics on the composition of the gut microbiota. A substantial number of studies in this area were conducted in the 1980 s and 1990 s and relied on culture-based techniques. However, researchers indicate that up to 80 % of gut bacteria are nonculturable[118 ]. Therefore, the focus has shifted to culture-independent approaches mainly based on 16 S rRNA gene sequence analysis.
Extensive research has established that antibiotic treatment induces a dramatic loss of diversity and remarkable shifts in community composition (Table 3 ), with the time of recovery varying substantially[119 -123 ].
The inconsistency in the results of various studies can be attributed to substantial heterogeneity in sample characteristics (age, ethnicity, diet,etc.) and study methodology. Furthermore, antibiotic characteristics, such as their class, pharmacokinetics (absorption and excretion), range of action, and dosing regimen, have been shown to shape the response of the gut microbiota to antibiotic perturbation[124 ].For instance, vancomycin is poorly absorbed when administered orally, resulting in high fecal concentrations. Therefore, it significantly alters the composition of the gut microbiota by increasing pathogenicProteobacteria, such asKlebsiella,Escherichia, andShigella, and decreasing members of theBacteroidetesphylum[122 ]. Lipophilic antibiotics (e.g.,lincosamides and macrolides) are eliminated mainly by biliary excretion and therefore cause profound changes in the intestinal microbiota[125 ]. For example,treatment with clindamycin resulted in a reduction in microbial diversity and a decrease inRoseburia,Lachospira, Coprococcus, Dorea,andRuminococcus. Changes in microbial composition were observed throughout 12 mo after clindamycin exposure[121 ]. In a recent study conducted by Haak et al[123 ], it was shown that treatment with broad-spectrum antibiotics (ciprofloxacin, vancomycin, and metronidazole) promotes the growth ofStreptococcusandLactobacillus.Furthermore, the authors found reduced numbers of anaerobes producing SCFAs, such asBacteroides,Subdoligranulum, andFaecalibacterium. Interestingly, a return toward baseline was observed between 8 and 31 mo, but the composition of the microbiota often remained changed from its initial state.
There is some evidence that antibiotics can indirectly affect the composition of the gut microbiota.This is due to interdependence among different microbial taxa, as they have a variety of shared metabolic pathways[124 ,126 ]. Thus, the loss or reduction of certain taxa affects the growth of other members of the community. As an example, vancomycin treatment reduces the number of Gramnegative commensals, although this drug selectively targets Gram-positive bacteria[127 ].
In a recent systematic review, Zimmermanet al[128 ] summarized data from 129 studies on the effect of antibiotics on the composition of the gut microbiota. The authors concluded that the majority of antibiotics (amoxicillin, amoxicillin/clavulanate, cephalosporins, lipopolyglycopeptides, macrolides,ketolides, clindamycin, tigecycline, quinolones, and fosfomycin) increase the abundance ofEnterobacteriaceae,mainlyCitrobacterspp.,Enterobacterspp.,andKlebsiellaspp. These bacteria contain molecules that directly enhance the inflammatory response of the host and may play a significant role in the alteration of bile acid metabolism[129 ]. Moreover, expansion of bacteria belonging to theEnterobacteriaceaefamily was associated with inflammatory bowel diseases, both in animal models and in humans[130 ,131 ]. Zimmerman et al[128 ] reported that amoxicillin, piperacillin, ticarcillin, cephalosporins (except fifth generation cephalosporins), carbapenems, and lipoglycopeptides facilitate the overgrowth ofEnterococcus,while treatment with macrolides and doxycycline results in decreased numbers of these bacteria. It has conclusively been shown that piperacillin, ticarcillin, carbapenems, macrolides,clindamycin, and quinolones markedly reduce the abundance of anaerobic bacteria. Finally, the authors documented that the most long-lasting changes in the community structure are caused by ciprofloxacin(1 year), clindamycin (2 years), and clarithromycin plus metronidazole (4 years).
Another negative effect of antibiotic treatment is the loss of colonization resistance. Depletion of beneficial gut commensals, such asLachnospiraceae, Ruminococcaceae,andClostridium scindens, as well as changes in their metabolic activity promote overgrowth ofClostridium difficile, Enterococcus, and other pathogens[33 ,124 ].
Antibiotics as a risk factor for IBS
Data from large cohort and case-control studies indicate that antibiotics are a risk factor for functional gastrointestinal disorders and IBS in particular. A retrospective study on more than 26000 patients showed that exposure to macrolides and tetracyclines may be associated with the development of IBS[5 ]. Similarly, a prospective case-control study found that antibiotic treatment of nongastrointestinal infections was associated with the development of IBS [odds ratio (OR) = 2 .30 ; 95 % confidence interval(CI): 1 .22 -4 .33 ; P = 0 .01 ] and other functional gastrointestinal disorders (OR = 1 .90 ; 95 %CI: 1 .21 -2 .98 ;P=0 .005 )[6 ]. A longitudinal study by Кrogsgaard et al[7 ] also identified that the use of antibiotics was a predictor for IBS (OR = 1 .8 ; 95 %CI: 1 .0 -3 .2 ). Additionally, a recent meta-analysis showed that the use of antibiotics for infectious enteritis was associated with an increased risk of IBS (OR = 1 .69 ; 95 %CI: 1 .20 -2 .37 )[8 ].
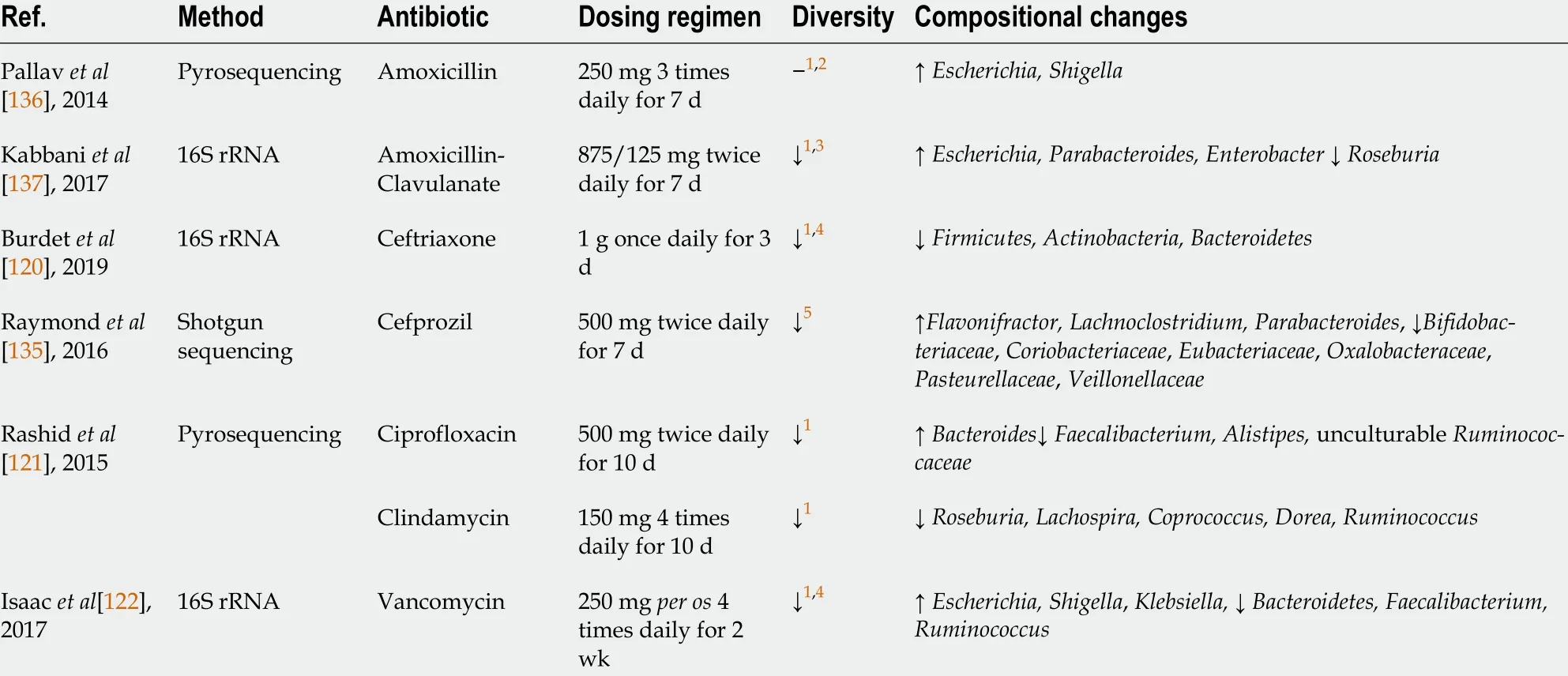
Table 3 Effects of antibiotics on gut microbiota composition (based on culture-independent approaches)
However, nonabsorbable antibiotics can be used to treat IBS. In a double-blind, randomized, placebocontrolled study, treatment with neomycin resulted in a 35 % improvement in composite scores of IBS symptoms, compared with only 11 % for placebo (P < 0 .05 )[132 ]. Nonetheless, the use of this antibiotic is limited by the risk forC.difficileinfection and systemic adverse events. A recent meta-analysis of four studies and 1803 patients showed that rifaximin was more effective than placebo in the overall improvement of IBS symptoms (OR = 1 .19 ; 95 %CI: 1 .08 -1 .32 and OR = 1 .36 ; 95 %CI: 1 .18 -1 .58 ,respectively,P< 0 .05 for both). There was no difference in adverse events between rifaximin and placebo[133 ]. Due to its safety, rifaximin was approved by the Food and Drug Administration for the treatment of IBS-D.
Similarities in gut microbiota between patients with IBS and those after antibiotic exposure
Analysis of data on changes in the gut microbiota in patients with IBS and those after antibiotic exposure uncovers some common features and trends. For instance, decreased microbial diversity[97 -99 ,121 ,128 ] and a reduction in the abundance of Faecalibacterium, particularly F. prausnitzii[97 ,98 ,102 ,121 ,122 ],have been observed in both cases.F. prausnitziiis one of the most abundant bacterial species in the gut, exhibiting anti-inflammatory effects through inhibition of IL-8 production, promotion of IL-10 secretion, and upregulation of regulatory T cells[134 ]. Moreover, patients with IBS were shown to have reduced numbers ofBifidobacterium[98 ,103 ,104 ,106 ,107 ]. Likewise, several studies have reported a decreased abundance of these commensals after antibiotic exposure[121 ,128 ,135 ]. Most members of the genusBifidobacteriumare known to exert beneficial effects on host health, including competitive exclusion of enteric pathogens, metabolism of dietary compounds, and regulation of the immune response[22 ,26 ,33 ]. Furthermore, both IBS and antibiotic exposure are characterized by overgrowth ofEnterobacteriaceae[98 ,104 ,106 ,136 ,137 ].TheEnterobacteriaceaefamily includes pathogenic bacteria (e.g.,Escherichia, Shigella, Klebsiella,andEnterobacter)with proinflammatory properties that may contribute to low-grade inflammation in the gut wall[98 ].
CONCLUSION
There is clear and consistent evidence from a variety of studies that patients with IBS have altered composition of the gut microbiota and that these alterations are related to the generation of gastrointestinal symptoms. However, studies comparing fecal microbiota in patients with IBS and healthy controls produced variable findings. To date, there is still no consensus on distinct microbiome signatures in IBS. Although some common threads reviewed here were found, prospective large-scale studies need to be carried out to shed light on this issue. Independent analysis of the gut microbiota and its metabolites will help to develop novel microbiota-based treatment strategies that target the underlying pathophysiology of IBS rather than focusing on symptom alleviation.
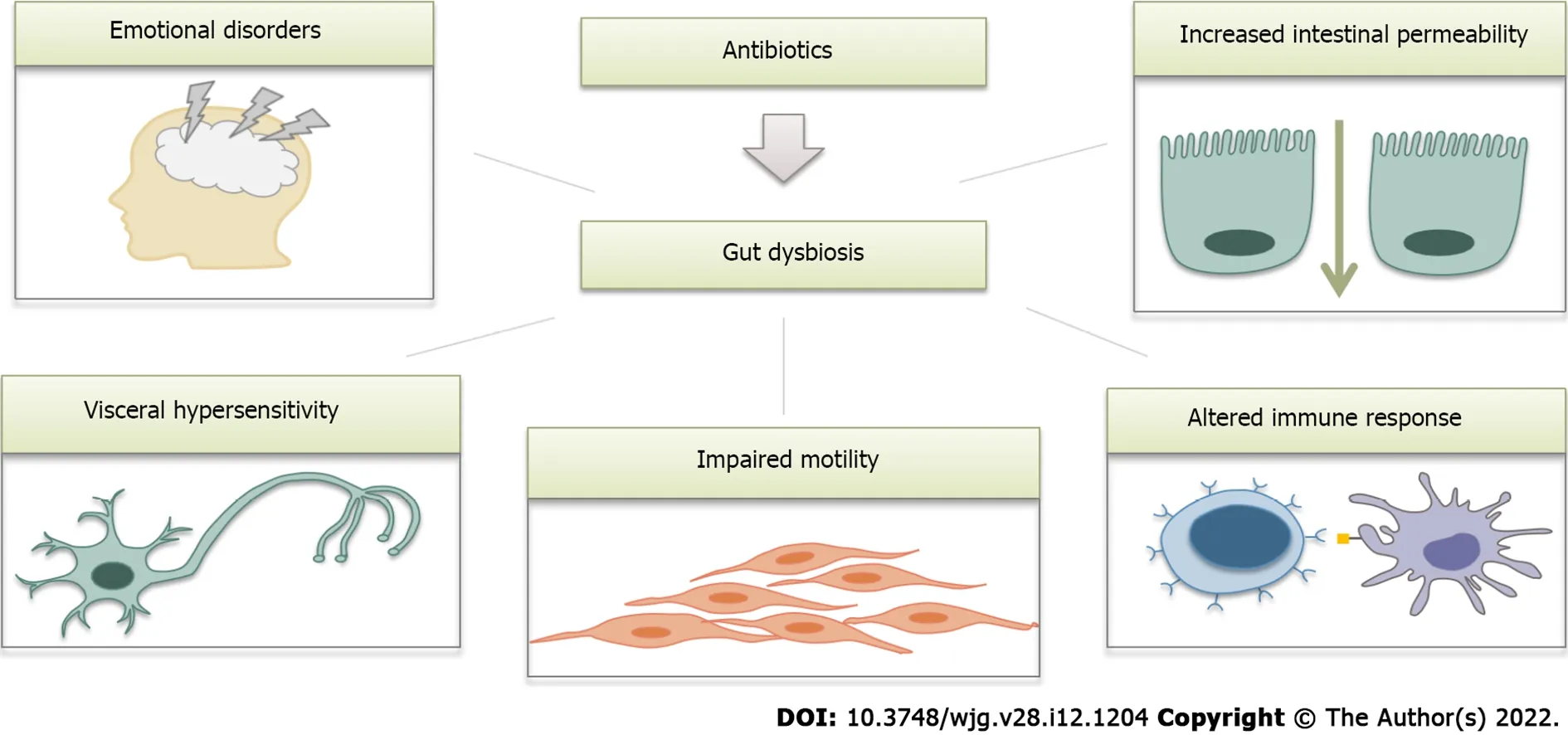
Figure 3 Possible link between antibiotic use and the development of irritable bowel syndrome (schematic illustration). Antibiotics cause profound changes in the gut microbiota and therefore contribute to all mechanisms involved in the pathogenesis of irritable bowel syndrome.
A number of recent studies have addressed the effects of antibiotics on gut microbiota composition,and these effects were found to be quite similar to those observed in IBS. We suggest that the Rome V criteria could provide a new definition of postantibiotic IBS. As major disruptors of the gut microbiota,antibiotics seem to contribute to all aspects of IBS pathogenesis (Figure 3 ). However, further research in this area is definitely warranted.
FOOTNOTES
Author contributions:Mamieva Z and Poluektova E took the lead in writing the manuscript; Svistushkin V and Sobolev V contributed to interpreting the relevant literature, provided critical feedback, and helped shape the manuscript; Shifrin O, Guarner F, and Ivashkin V substantially contributed to the conception and design of the article and revised the paper; all authors read and approved the final manuscript.
Conflict-of-interest statement:The authors declare no conflict of interests for this article.
Open-Access:This article is an open-access article that was selected by an in-house editor and fully peer-reviewed by external reviewers. It is distributed in accordance with the Creative Commons Attribution NonCommercial (CC BYNC 4 .0 ) license, which permits others to distribute, remix, adapt, build upon this work non-commercially, and license their derivative works on different terms, provided the original work is properly cited and the use is noncommercial. See: https://creativecommons.org/Licenses/by-nc/4 .0 /
Country/Territory of origin:Russia
ORCID number:Zarina Mamieva 0000 -0002 -5673 -7920 ; Elena Poluektova 0000 -0002 -9038 -3732 ; Valery Svistushkin 0000 -0001 -7414 -1293 ; Vasily Sobolev 0000 -0002 -7372 -3299 ; Oleg Shifrin 0000 -0001 -8148 -2862 ; Francisco Guarner 0000 -0002 -4051 -0836 ; Vladimir Ivashkin 0000 -0002 -6815 -6015 .
S-Editor:Fan JR
L-Editor:Wang TQ
P-Editor:Fan JR
 World Journal of Gastroenterology2022年12期
World Journal of Gastroenterology2022年12期
- World Journal of Gastroenterology的其它文章
- Emerging role of colorectal mucus in gastroenterology diagnostics
- Similarities, differences, and possible interactions between hepatitis E and hepatitis C viruses: Relevance for research and clinical practice
- Microbiologic risk factors of recurrent choledocholithiasis postendoscopic sphincterotomy
- Epidemiological, clinical, and histological presentation of celiac disease in Northwest China
- Near-infrared fluorescence imaging guided surgery in colorectal surgery
- Spinal anesthesia alleviates dextran sodium sulfate-induced colitis by modulating the gut microbiota