
雷州半岛以东近岸海域浮游动物群落结构与环境因子关系
2022-03-31 09:08:58黄彬彬吴风霞郑淑娴田丰歌
广东海洋大学学报 2022年2期
黄彬彬,吴风霞,郑淑娴,田丰歌,何 静
雷州半岛以东近岸海域浮游动物群落结构与环境因子关系
黄彬彬1,2,吴风霞3,郑淑娴4,田丰歌1,2,何 静1,2
(1.国家海洋局南海环境监测中心 // 2.自然资源部海洋环境探测技术与应用重点实验室 // 3.中国水产科学院南海水产研究所//4.国家海洋局南海规划与环境研究院,广东 广州 510300)
【目的】研究雷州半岛以东近岸至浅海过渡区海域浮游动物群落结构与环境因子的关系,探索浮游动物群落动态变化规律。【方法】根据2017-2018年在雷州半岛以东近岸海域四个季节的调查资料,对该海域浮游动物种类组成、丰度、生物量、优势种和群落结构进行研究。【结果与结论】四个季节共鉴定浮游动物208种,浮游幼体23类,隶属于18个类群,以桡足类最多,水螅水母类次之;浮游动物丰度平均值以春季最高(972.11 m-3),夏季(646.86 m-3)和秋季(177.27 m-3)次之,冬季最低(98.92 m-3);春季生物量(485.45 mg·m-3)明显高于其他季节,夏季生物量(72.81 mg·m-3)最低;不同类群和优势种丰度存在季节变化,春夏季枝角类丰度最高,秋冬季以桡足类为主,鸟喙尖头溞()为春夏季第一优势种,肥胖软箭虫()和精致真刺水蚤()分别为秋、冬季第一优势种。聚类分析将浮游动物群落在40%相似性水平上分为5个组群,即春季组群1和组群2、夏季组群、秋季组群和冬季组群。相关性分析表明,水深、水温、盐度和叶绿素a是影响海区浮游动物群落结构的主要因素。
浮游动物;群落结构;聚类分析;雷州半岛
雷州半岛位于我国大陆最南端,半岛东面为南海西北部的广阔海域,南部毗邻琼州海峡,与海南岛隔海相望,其岸线曲折,近岸生态系统多样,海洋生物资源丰富[1]。自20世纪60年代开始,对雷州半岛沿岸海域浮游动物群落已有不少报道,如鉴江口和湛江港(湾)海域桡足类种类组成[2-5],对雷州半岛沿岸及灯楼角珊瑚礁海区浮游动物种类组成、生态类群和数量分布[6-7],以及对硇洲岛海藻场浮游动物群落结构和季节变化等[8]。以往对雷州半岛海域浮游动物的研究主要集中在河口、海湾和岛屿附近海域,而对近岸至浅海过渡区的报道较少。雷州半岛以东近海属南海西北部大陆架海域,该海域流系复杂,主要受到粤西沿岸流、雷州半岛东部近海气旋式环流(冷涡)和南海暖流的影响,多种流系的相互作用使该海域具有相对独特的环境特征和较高的生物多样性[9-11]。本研究通过采样调查对该海区浮游动物种类组成、丰度时空分布和群落结构进行研究,并结合环境数据对主要影响因子进行分析,可为南海海洋生物多样性研究和海洋生态环境评价提供科学依据,为丰富该海域海洋生态系统结构和功能研究提供基础资料。
1 材料与方法
1.1 站位布设和采样
如图1所示,2017-2018年间于春季(5月)、夏季(8月)、秋季(10月)和冬季(2018年1月)在雷州半岛以东海域布设13个站位(冬季在8号和10号站未采样),对海区浮游动物进行调查。采样及样品处理均按照文献[12]提供的方法进行。使用浅水Ⅰ型浮游生物网和大型浮游生物网(水深> 30 m时)由海底至海面垂直拖曳采集浮游动物。采集到的样品加入体积分数约5%的中性甲醛溶液进行固定,带回实验室在体视显微镜下进行种类鉴定和个体计数。对水温、盐度、pH、溶解氧和叶绿素a等环境因子进行同步采样测定。水温使用CTD现场测定,盐度、pH和溶解氧分别使用盐度计法、电位计法和碘量法测定,叶绿素a浓度使用分光光度法测定。
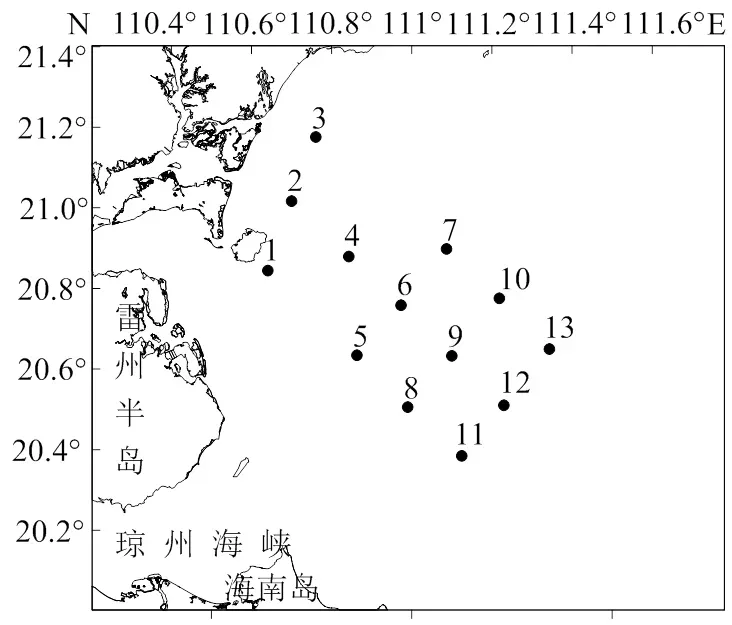
图1 雷州半岛以东海域采样站位
1.2 数据处理
1.2.1 多样性指数和优势度
优势度:(n/) ×f,
均匀度:/ log2,
其中,P为第种的个数与本站总个数的比值,n和f分别为第种浮游动物的个体数与出现频率,为种类数。根据≥0.02划分优势种[13]。
1.2.2 统计分析 群落相似性聚类分析和群落结构分析使用多元统计软件PRIMER V5.2(Plymouth Routines In Multivariate Ecological Research)进行。选取海区出现频率在20%以上的浮游动物种类用于群落结构分析,以减少机会种对结果的干扰,丰度通过平方根转换以降低数据的极化程度[14]。采用Bray-Curtis相似性系数进行等级聚类[15]。采用One-way ANOSIM程序检验群落划分结果的差异显著性。结果分析中不同值表明差异显著(< 0.05)或差异极显著(< 0.01)。
使用SPSS23.0软件,利用皮尔森相关系数(Pearson Correlation Coefficient)对浮游动物丰度、种类数、多样性与水温、盐度等环境因子的相关性进行分析。
2 结果
2.1 丰度和生物量
调查海域浮游动物丰度季节差异较大,春夏季明显高于秋冬季。春季丰度平均值为972.11 m-3,为各季节最高,夏季次之,平均值为646.86 m-3,秋季丰度出现大幅下降,平均值为177.27 m-3,冬季丰度最低,平均为98.92 m-3。
春季和夏季浮游动物丰度分布规律基本一致,高值区主要分布在沿岸的1 ~ 4号站,由此向外,随着水深增加,丰度呈下降趋势(图2);秋季丰度分布趋势不明显,呈斑块状分布,仍以1号站最高;冬季硇洲岛附近的2号和1号站丰度高于其他站位。
图2 浮游动物丰度平面分布
Fig.2 Horizontal distribution of zooplankton abundance
调查海区春季浮游动物生物量显著高于其它季节,平均值为485.45 mg·m-3,其次为冬季和秋季,分别为162.85 mg·m-3和133.07 mg·m-3,夏季生物量最低,平均值为72.81 mg·m-3。夏季和冬季生物量分布趋势与丰度基本一致,从沿岸向外,生物量总体呈下降趋势(图3)。春季生物量分布与丰度并不一致,8号、9号站由于海樽数量较多,形成生物量高值中心,其次才是沿岸区域。相关性分析表明,浮游动物生物量与丰度呈极显著正相关(= 0.392,< 0.01)。
2.2 群落结构分析
2.2.1 种类组成 经统计,海区4个季节共鉴定浮游动物208种,隶属于8个门18个类群(表1)。其中以桡足类最多,共计出现87种,占总种数的41.8%,其次为水螅水母类34种,占总种数的16.3%,管水母类出现18种,占总种数的8.7%。此外记录到各类生物的浮游幼体23类。夏季种类最多,共出现134种,其次为秋季和春季,分别出现96种和93种,冬季最少共记录71种。

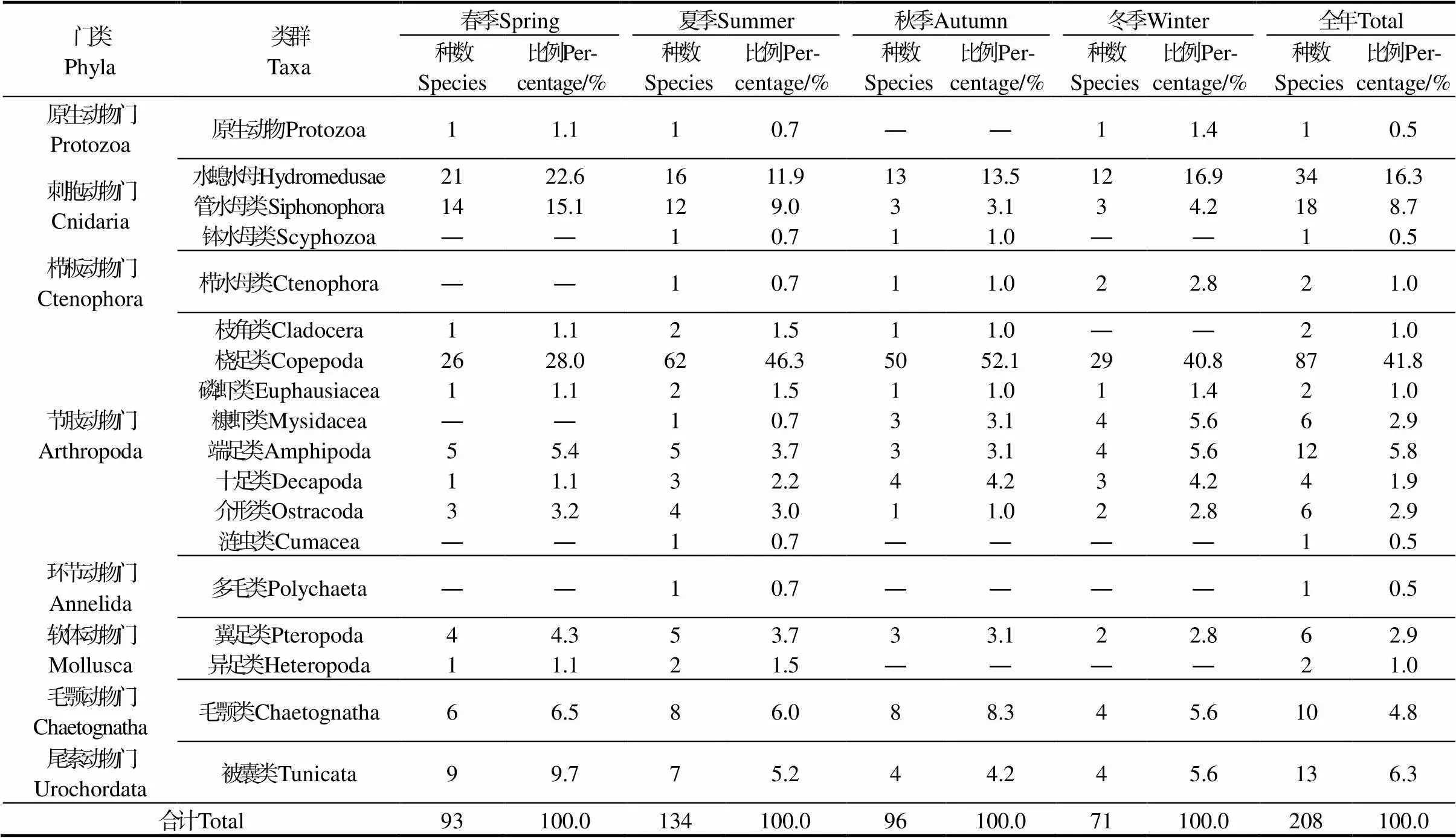
表1 浮游动物种类组成
注:“―”表示该类群未出现在本航次。Note: “—” indicates this taxa was absent from the survey.
浮游动物种类平面分布不均匀,各个季节总体表现为由沿岸向近海,种类数随水深增加而增加。春季种类数在29 ~ 47之间,除离岸最远的12、13号站种类数超过40外,其余站位相对均匀;夏季种类数介于23 ~ 73,5号站最低,12号站最高;秋季种类数在26 ~ 56之间,4号站最低,12号站最高;冬季种类数在15 ~ 44之间,3号站最低,13号站最高。
2.2.2 主要类群和优势种 浮游动物各类群丰度差异大,各季节优势类群不尽相同。枝角类丰度春季最高,为572.47 m-3,夏季次之,春夏季丰度百分比分别为58.9%、39.6%,均为当季丰度最高的类群,冬季则未出现;被囊类丰度春季较高,达240.42 m-3,其余季节都较低;桡足类丰度夏季最高,为138.61 m-3,丰度比例为21.4%,仅次于枝角类,秋冬季丰度虽不及夏季,但在群落中占绝对优势,丰度比例都超过40%,春季桡足类丰度最低。浮游幼体也是海区重要类群之一,夏季丰度最高,为95.91 m-3,仅次于枝角类和桡足类,其次为春季,冬季丰度最低,仅为11.21 m-3。
如表2所示,各季节浮游动物优势种组成有所不同,全年共统计优势种21种。春季鸟喙尖头溞()和双尾纽鳃樽()占绝对优势,两者丰度合计占总丰度76.6%,前者主要分布在近岸水深较浅的1 ~ 7号站,后者则主要分布水深较大的8 ~ 13号站。夏季以鸟喙尖头溞和肥胖三角溞()两种枝角类优势度最高,两者丰度合计占总丰度39.6%,分布仍以浅水区为主,与春季相比,鸟喙尖头溞分布范围扩大,除13号站外均有出现。秋季肥胖软箭虫()优势度最高,所有站位均有分布。精致真刺水蚤()、微刺哲水蚤()和宽额假磷虾()在冬季占绝对优势,三者丰度合计占总丰度42.8%,主要分布在沿岸浅水区。
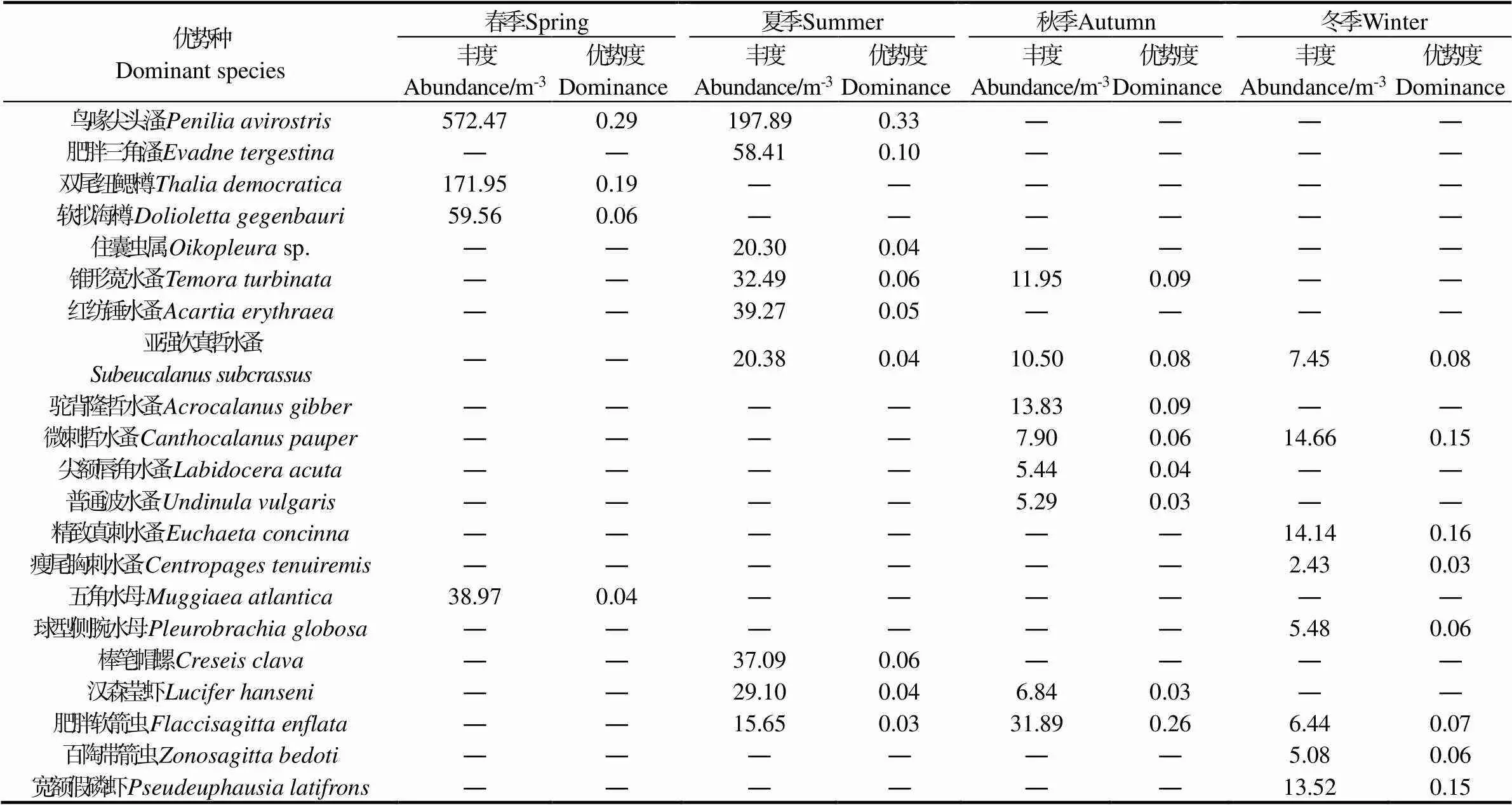
表2 浮游动物优势种的优势度和平均丰度
注: “―”表示该种优势度< 0.02,未判定为优势种。Note: “—” indicates specie dominance was lower than 0.02.
2.2.3 多样性指数和均匀度 浮游动物多样性指数变化范围为1.35 ~ 4.90,全年平均值为3.15。春季由于优势种优势度偏高,多样性指数普遍较低,介于1.35 ~ 2.86之间,平均为2.01,为全年最低;夏季多样性指数介于1.82 ~ 4.90之间,平均值3.72,为全年最高;秋季多样性指数在2.56 ~ 4.18之间,平均为3.32;冬季平均值3.57,变化范围3.35 ~ 3.83。群落均匀度变化范围为0.26 ~ 0.92,全年平均值为0.63。各季节平均值以冬季最高(0.76),其次为夏季(0.69)和秋季(0.69),春季最低(0.36)。
2.2.4 群落聚类分析 选择海区出现率在20%以上的种类,根据种类丰度对站位进行等级聚类,结果如图4所示。按照40%的相似性,可以将海区4个季节浮游动物群落划分为5个组群,第1组和第2组为春季群落,第3组为夏季群落,第4组为秋季群落,第5组为冬季群落,其中夏季10号、12号和13号站被划入秋季群落。聚类分析的结果表明,浮游动物群落具有较为显著的季节性变化特征。
One-way ANOSIM检验的结果(表3)表明,以浮游动物丰度划分的5个组群中,任意2个组群之间的差异性显著(Global Test:= 0.902,= 0.001)。其中,组群1、组群5之间以及组群2、组群5之间差异最大(= 1.000,= 0.001),而组群3、组群4之间差异最小(= 0.706,= 0.001)。
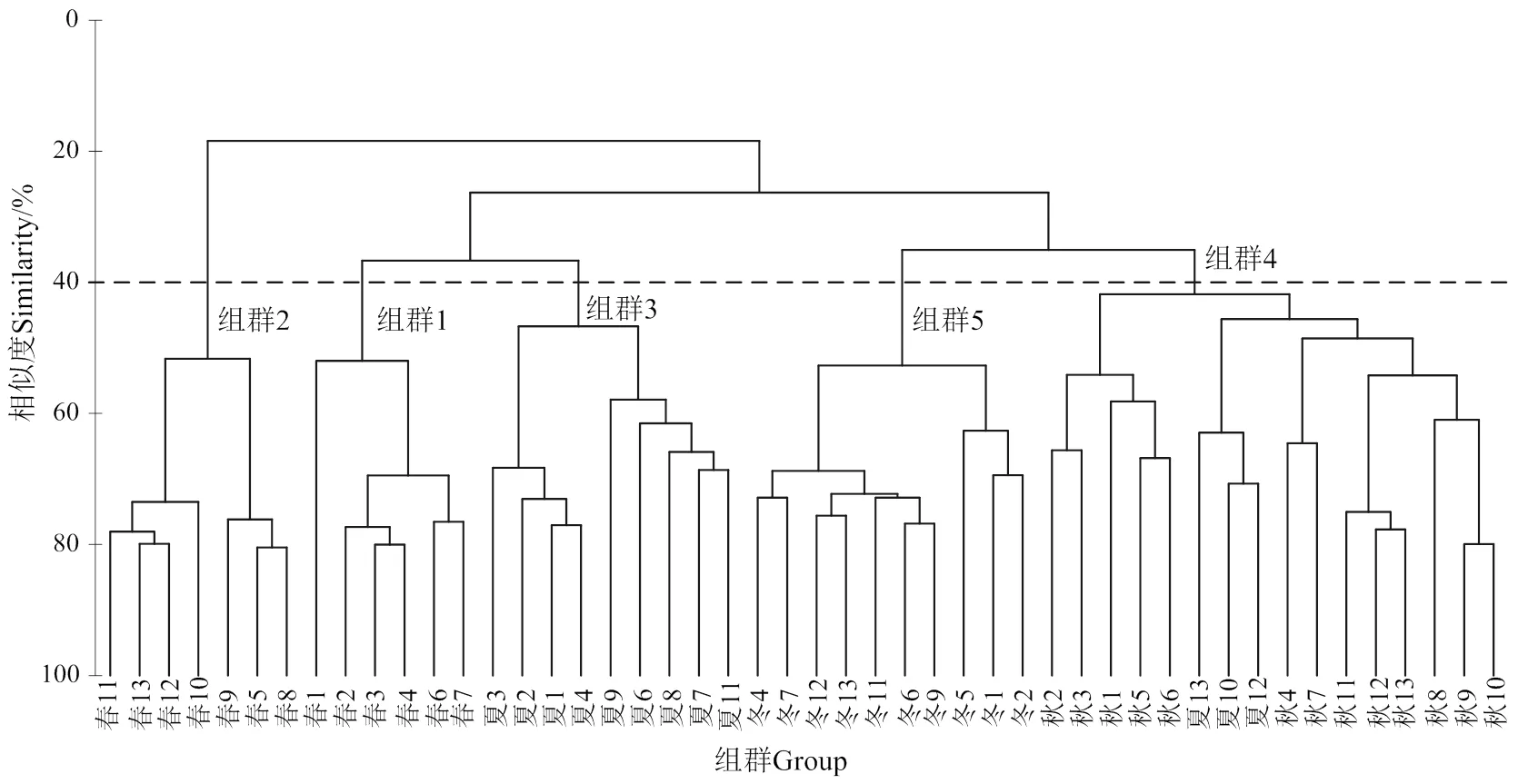
图4 调查站位浮游动物群落等级聚类

表3 One-way ANOSIM结对检验结果
2.3 浮游动物群落与环境因子相关性
将浮游动物丰度、生物量、种类数、多样性与水温、盐度等环境因子进行相关性分析,结果如表4所示。浮游动物丰度与叶绿素a呈极显著正相关(< 0.01),与溶解氧呈显著正相关(< 0.05),与水深则呈极显著负相关(< 0.01);种类数则与水深、水温和盐度均呈极显著正相关(< 0.01),与叶绿素a呈极显著负相关(< 0.01),与溶解氧呈显著负相关(< 0.05);生物量与丰度呈极显著正相关(< 0.01),与其它环境因子相关性不显著。
表4 浮游动物群落与环境因子相关性分析

Table 4 Correlation analysis between zooplankton community and the environmental factors
注: **表示相关性极显著(< 0.01),*表示相关性显著(< 0.05)。
Note: ** means highly significantly correlated at the 0.01 level, and * means significantly correlated at the 0.05 level.
3 讨论
3.1 浮游动物与环境因子的关系
浮游动物群落结构受多个环境因子的共同影响。水深不同决定了对浮游动物空间容量的差异。调查海域属近岸向浅海的过渡地带,水深变化较大,浮游动物种类数与水深相关性极显著(< 0.01),由近岸向外海显著增加。近岸浅水区水深不足20 m,种类较少,冬季可低至20种以下,而外围海域水深达50 m,夏季种类数超过60种,形成了种类多样化的特点,符合种类数随水深增加而增加的一般规律[16]。由于近岸海域受陆地工农业生产和人类生活污水排放影响,营养物质丰富,饵料来源充足,有利于浮游动物的大量繁殖,所以丰度较外海更高。
水温、盐度也是影响浮游动物群落结构的重要因素。相关性分析显示浮游动物种类数与水温、盐度均呈极显著正相关(< 0.01)。水温可以影响浮游动物的新陈代谢强度,进而影响浮游动物的生长、发育和繁殖,温度过高或过低均不利于浮游动物的增殖[17-18]。如中华哲水蚤的适温范围虽然较宽,在1 ~ 27 ℃之间均可生存,但其繁殖盛期最适温度在10 ~ 18 ℃,上限大约为23 ℃[19-21]。调查海域水温季节性变化明显,表层水温平均值夏季最高(29.4 °C),秋季(28.3 °C)和春季(26.2 °C)略低,冬季最低(17.2 °C)。浮游幼体种类和丰度可以反映海区浮游动物群落的发展水平。春季水温回升,浮游动物进入繁殖旺盛期,浮游幼体种类为各季节之最,夏季水温最高,幼体丰度也达到最高峰,而冬季较低水温已不适宜多数浮游动物的繁殖,幼体种类和丰度均为各季节最低。浮游幼体丰度与水温呈现正相关,说明水温是影响浮游动物群落发展的重要环境因子。盐度对于浮游动物维持正常的生理活动具有重要意义,并通过影响浮游动物的发育繁殖使其生态特征发生改变[22-23]。调查海区表层盐度平均值以冬季最高(33.2),夏季最低(30.4),受沿岸流影响,近岸在春夏季呈现一定的低盐特征。春夏季浅水区浮游动物以低盐类群为主,种类少,但丰度高,深水区主要受外海水影响,高盐环境决定了群落以外海类群为主,种类数高,但丰度低。
水温、盐度为浮游动物的生长发育提供了环境基础,食物供应则是其存活的主导因子[8]。浮游动物丰度与叶绿素a之间极显著的正相关性表明,充足食物来源对浮游动物种群的维持至关重要。春季随着水温升高,浮游植物大量增加为摄食者提供了充足食物来源,在水温合适、饵料充足的条件下,浮游动物数量迅速增加。秋季浮游植物数量出现大幅下降,浮游动物数量相应减少。冬季叶绿素a含量虽高于秋季,但浮游动物丰度并未随之增加,而是继续下降,造成这种不一致的原因是海区水温较低,不利于浮游动物的繁殖。通过对环境因子与浮游动物群落结构的关系分析,可以看出,浮游动物群落结构变动与水温、盐度、浮游植物等多个因素密切相关,是多个生物和非生物因素共同作用的结果。
3.2 浮游动物群落结构与水团的关系
海流与水团是影响浮游动物水平分布格局的重要因素,对群落结构和优势种的演替具有重要影响[24]。雷州半岛东部近岸终年存在向西流动的粤西沿岸流,冬季较弱,夏季最强,雷州半岛以东海域在夏季还存在气旋式环流[25-26]。
不同水团对浮游动物群落结构的影响体现在群落聚类的结果中。春末是南海夏季季风开始的时期,加上沿岸径流输入,粤西沿岸流逐渐加强[10],近岸低盐水团在海域浅水区的影响增强。从群落聚类结果来看,组群1包含了1-4、6-7号站,该组群是以鸟喙尖头溞为主要优势种的近岸低盐类群。鸟喙尖头溞作为一种典型的近岸低盐种,对沿岸水团具有一定指示作用。在春季该种主要分布在水深30 m以浅的海域,具有极高丰度和优势度,体现了沿岸流在近岸的影响。组群2则代表了近岸以外海域,春季沿岸流在该区域的影响较弱,形成了以双尾纽鳃樽等亚热带外海种为代表的外海类群。
夏季粤西沿岸流强度最大,影响范围最广,与外海水混合后形成高温低盐的近岸混合水团在海区占主导地位[27]。聚类结果显示,夏季多数站位都划入了组群3,反映了沿岸流的影响范围较春季更大,群落中鸟喙尖头溞和肥胖三角溞几乎遍布整个海域。在沿岸流与外海水共同作用下,近岸低盐类群和近岸暖水类群成为海区主要优势类群。
秋冬季随着沿岸流强度减弱,外海水团的影响增强,浮游动物群落结构发生改变,以鸟喙尖头溞为代表的低盐类群退出群落,近岸暖水类群分布更加广泛,外海高盐种类增加,精致真刺水蚤、宽额假磷虾等外海种在冬季分布范围可延伸至近岸浅水区。聚类的结果将秋冬季站位分别划分为两个组群,并将夏季外围站位划入秋季组群,表明外海水团的影响在秋冬季占主导地位,并且这种影响在一定程度上降低了群落结构的空间差异性。
3.3 浮游动物优势种的生态适应
浮游动物群落结构随海流季节性消长而呈现周期性变化,种类组成和个体丰度处于不断波动之中。优势种的时空分布和更替很大程度上决定了浮游动物群落结构的动态变化。
海区浮游动物群落优势种具有显著的季节变化特征。枝角类在春夏季大量出现,在浮游动物群落中占绝对优势地位。鸟喙尖头溞在春夏季均为第一优势种,且优势度较高。根据徐兆礼等[28]研究,该种具有广温广盐的适应性,最适温度高于25 ℃,最适盐度低于20,所以应属于亚热带近海种,其高丰度分布区可以作为沿岸水团的标志,可以指示沿岸流影响的边界。本研究中,鸟喙尖头溞在春季和夏季的分布存在差异。春季在浅水区形成高密度分布区,而在离岸较远的海域受外海高盐水的影响,数量急剧减少,海区出现率仅为46%,呈现明显的近岸分布,夏季其分布范围较春季扩大,外围海域仍有一定的数量,秋冬季随着水温的下降和外海水影响的增强,逐渐从群落中消失。笔者认为,鸟喙尖头溞的空间分布和季节变动与沿岸流的季节性变化密切相关,可以作为表征粤西沿岸流影响边界的指示种。
双尾纽鳃樽是春季另一个主要优势种,丰度仅次于鸟喙尖头溞。海樽类多数物种可以作为水团或海流的指示种[29]。对东海海樽类的研究[30]表明,该种为亚热带外海种,在冬季就具有一定的数量,春季发展为最主要的优势种,其高丰度水域,能够较为准确地反映黑潮入侵陆架水团和沿岸水交汇的位置。本调查中该种高丰度同样出现在春季,从其分布来看,密集分布区主要在水深大于30 m,盐度大于32的站位,呈现显著的高盐特征。对南海中沙西沙海域的研究[31]认为,海樽类平面分布和及群落格局与周边海域存在的气旋式环流、上升流及暖流等的变化有较好的相关性。本研究中,双尾纽鳃樽的分布区域与雷州半岛以东的气旋式环流存在重合,两者之间具有怎样的关联尚需进一步研究。但结合其分布和沿岸流的特征来看,笔者认为该种高丰度聚集区对外海水和沿岸流的交汇区域亦具有一定指示作用。这也就是夏季沿岸流影响范围扩大时该种数量大大降低的原因。
肥胖软箭虫为暖水广布种,在本海区四个季节均有出现,优势度以秋季最高,为当季第一优势种,其次为夏季、冬季,春季优势度最低。徐兆礼等[32]认为在东海该种的分布与外海暖水团的关系密切,从春季到秋初,丰度随水温的升高而增加。对北部湾北部海域的调查发现,该种在四季均为主要优势种,夏秋两季丰度较高,冬春季随水温的降低数量有所减少[33]。本研究中,该种季节变化趋势与北部湾一致,春季数量较少,夏季随着水温的升高,丰度显著增加,至秋初仍维持在高丰度,冬季随着水温下降丰度也降至最低,显示与水温呈现出正相关。空间分布上,该种与前述的鸟喙尖头溞和双尾纽鳃樽有所不同,四个季节总体出现率高达94%,且各个季节数量分布相对均匀,无明显高丰度聚集区,与沿岸流和外海水团的季节性变化没有显著关系,显示其对环境有广泛适应性。
[1] 龚玉艳, 张才学, 孙省利, 等.2010年夏季雷州半岛海岸带浮游植物群落结构特征及其与主要环境因子的关系[J].生态学报, 2012, 32(19): 5972-5985.
[2] 沈嘉瑞, 李茯香.广东鉴江口与湛江港的桡足类[J].动物学报, 1963, 9(4): 571-592.
[3] 沈嘉瑞, 李茯香.广东湛江口的桡足类[J].动物分类学报, 1966(3): 213-223.
[4] 张才学, 龚玉艳, 王学锋, 等.湛江港湾浮游桡足类群落结构的季节变化和影响因素[J].生态学报, 2011, 31(23): 101-111.
[5] 龚玉艳, 张才学, 陈作志, 等.湛江湾浮游动物群落结构特征及其周年变化[J].海洋科学, 2015, 39(12): 46-55.
[6] 尹健强, 黄晖, 黄良民, 等.雷州半岛灯楼角珊瑚礁海区夏季的浮游动物[J].海洋与湖沼, 2008, 39(2): 131-138.
[7] 张才学, 龚玉艳, 孙省利, 等.2010年夏季雷州半岛沿岸海域浮游动物群落结构特征[J].海洋学报, 2014, 36(4): 91-99.
[8] 刘耀谦, 孙省利, 张才学.2014—2015年硇洲岛大型海藻场浮游动物的群落结构和季节变化[J].海洋学报, 2018, 40(12): 94-111.
[9] 尹健强, 黄良民, 李开枝, 等.南海西北部陆架区沿岸流和上升流对中华哲水蚤分布的影响[J].海洋学报, 2013, 35(2): 143-153.
[10] 杨士瑛, 鲍献文, 陈长胜, 等.夏季粤西沿岸流特征及其产生机制[J].海洋学报, 2003, 25(6): 1-8.
[11] 谢玲玲, 曹瑞雪, 尚庆通.粤西近岸环流研究进展[J].广东海洋大学学报, 2012, 32(4): 94-98.
[12] 国家质量监督检验检疫总局, 中国国家标准化管理委员会.海洋调查规范第6部分: 海洋生物调查: GB/T 12763.6—2007[S].北京: 中国标准出版社, 2008.
[13] 徐兆礼, 陈亚瞿.东黄海秋季浮游动物优势种聚集强度与鲐鲹渔场的关系[J].生态学杂志, 1989, 8(4): 13-15.
[14] ZHANG Q T, WARWICK R M, MCNEILL C L, et al.An unusually large phytoplankton spring bloom drives rapid changes in benthic diversity and ecosystem function[J].Progress in Oceanography, 2015, 137: 533-545.
[15] CLARKE K R, WARWICK R M.Similarity-based testing for community pattern: the two-way layout with no replication[J].Marine Biology, 1994, 118(1): 167-176.
[16] 郑重, 李少菁, 连光山.海洋桡足类生物学[M].厦门: 厦门大学出版社, 1992: 1-309.
[17] DEVREKER D, SOUISSI S, SEURONT L.Effects of chlorophyll concentration and temperature variation on the reproduction and survival of(Copepoda, Calanoida) in the Eastern English Channel[J].Journal of Experimental Marine Biology and Ecology, 2005, 318(2): 145-162.
[18] LENZ P H, HOWER A E, HARTLINE D K.Temperature compensation in the escape response of a marine copepod,(Crustacea)[J].The Biological Bulletin, 2005, 209(1): 75-85.
[19] WANG R.The Yellow Sea cold bottom water: an oversummering site for(Copepoda, Crustacea)[J].Journal of Plankton Research, 2003, 25(2): 169-183.
[20] HUANG C, UYE S, ONBÉ T.Geographic distribution, seasonal life cycle, biomass and production of a planktonic copepod Calarms sinicus in the Inland Sea of Japan and its neighboring Pacific Ocean[J].Journal of Plankton Research, 1993, 15(11): 1229-1246.
[21] LI C, SUN S, WANG R, et al.Feeding and respiration rates of a planktonic copepod () oversummering in Yellow Sea cold bottom waters[J].Marine Biology, 2004, 145(1): 149-157.
[22] MOUNY P, DAUVIN J C.Environmental control of mesozooplankton community structure in the Seine Estuary (English Channel)[J].Oceanologica Acta, 2002, 25(1): 13-22.
[23] FRONEMAN P W.Response of the plankton to three different hydrological phases of the temporarily open/closed kasouga estuary, South Africa[J].Estuarine, Coastal and Shelf Science, 2002, 55(4): 535-546.
[24] 郑白雯, 曹文清, 林元烧, 等.北部湾北部生态系统结构与功能研究Ⅰ.浮游动物种类组成及其时空变化[J].海洋学报, 2013, 35(6): 154-161.
[25] 伍伯瑜.珠江口以西陆架海域环流研究Ⅱ.[J].台湾海峡, 1990, 9(1): 14-21.
[26] 孙湘平, 修树孟, 苏玉芬.“东、南海陆架暖流”的初步探讨[J].海洋通报, 1996, 15(2): 1-10.
[27] 程国胜, 孙佳东, 俎婷婷, 等.2011年夏季南海北部海区水团分析[J].热带海洋学报, 2014, 33(3): 10-16.
[28] 徐兆礼, 高倩, 陈华, 等.东海浮游枝角类和涟虫类生态适应性[J].生态学杂志, 2007, 26(11): 1782-1787.
[29] 张金标, 连光山, 王云龙, 等.台湾海峡东部海域海樽类被囊动物的分布[J].台湾海峡, 2003, 22(3): 279-285.
[30] 徐兆礼, 孙军, 林茂.东海海樽类生态类群统计分析[J].生态学报, 2008, 28(11): 5698-5705.
[31] 李亚芳, 杜飞雁, 王亮根, 等.南海中沙西沙海域海樽类群落结构特征研究[J].南方水产科学, 2016, 12(4): 64-70.
[32] 徐兆礼, 陈亚瞿.东海毛颚类优势种及与环境的关系[J].中国水产科学, 2005, 12(1): 76-82.
[33] 郑白雯, 曹文清, 林元烧, 等.北部湾北部生态系统结构与功能研究Ⅱ.浮游动物数量分布及优势种[J].海洋学报, 2014, 36(4): 82-90.
Relationships Between Community Structure of Zooplankton And Environmental Factors in the East Leizhou Peninsula Coastal Area
HUANG Bin-bin1,2, WU Feng-xia3, ZHENG Shu-xian4, TIAN Feng-ge1,2, HE Jing1,2
(1.,// 2.,// 3.,// 4.,,510300,)
【Objective】This survey aimed to study the relationships between community structure of zooplankton and environmental factors in the coastal area of east Leizhou Peninsula .【Method】Seasonal sampling was performed from May 2017 to January 2018.The species composition, dominance, abundance, biomass,community structure of zooplankton were analyzed.【Result and Conclusion】A total of 208 zooplankton species with 23 categories of pelagic larvae were identified during the investigation, which belonging to 18 taxa.The copepods were the most abundant group, followed by hydromedusae.The average of zooplankton abundance was higher in spring (972.11 m-3) and summer (646.86 m-3) than that in autumn (177.27 m-3) and winter (98.92 m-3).The biomass of zooplankton was highest in spring (485.45 mg·m-3), and lowest in summer (72.81 mg·m-3).The abundance of different groups and dominant species changed seasonally.Cladocera had the highest abundance in spring and summer, while copepod was dominant in autumn and winter.was the most dominant species in spring and summer.They were replaced byandin autumn and winter respectively.By hierarchical cluster analysis, the zooplankton community could be divided into five assembles at a similarity of 40%, including spring groups 1 and 2, summer group, autumn group and winter group.Correlation analysis indicated that the water depth, temperature, salinity and chlorophyll a were the main factors influencing the structure of the zooplankton community.
Zooplankton; community structure; cluster analysis; Leizhou Peninsula
Q958.8
A
1673-9159(2022)02-0053-09
10.3969/j.issn.1673-9159.2022.02.007
2021-10-12
广东省渔业生态环境重点实验室开放基金项目(FEEL-2020-3);国家自然科学基金项目(41606191)
黄彬彬(1984-),男,工程师,研究方向为海洋生物生态。E-mail: bbhua2011@163.com
何静,工程师,研究方向为海洋浮游生物。E-mail: jinghe_2013@163.com
黄彬彬,吴风霞,郑淑娴,等.雷州半岛以东近岸海域浮游动物群落结构与环境因子关系[J].广东海洋大学学报,2022,42(2):53-61.
(责任编辑:刘岭)
猜你喜欢
桉树科技(2022年2期)2022-07-11 09:32:40
海洋信息技术与应用(2021年3期)2022-01-17 02:37:26
现代苏州(2021年3期)2021-09-10 07:12:13
江苏教育研究(2020年28期)2020-11-23 07:30:10
广州文博(2020年0期)2020-06-09 05:14:34
建筑热能通风空调(2018年5期)2018-07-09 03:16:44
湖北文理学院学报(2017年2期)2017-04-16 05:09:12
环球人文地理·评论版(2016年9期)2017-03-15 16:55:59
网络安全和信息化(2017年3期)2017-03-10 07:45:51
海洋渔业(2016年6期)2016-04-16 03:00:23
