
镉污染条件下水稻对假单胞菌TCd-1微生物修复的生理响应
2022-03-30 10:57:58文欢欢郑新宇肖清铁汪敦飞律其鑫韩博伦王玉洁樊荣荣郭尤睿林瑞余
生态学报 2022年5期
文欢欢,郑新宇,2,肖清铁,2,汪敦飞,律其鑫,韩博伦,王玉洁,樊荣荣,游 武,郭尤睿,钱 鑫,林瑞余,2,*
1 福建省农业生态过程与安全监控重点实验室,福建农林大学生命科学学院, 福州 350002 2 作物生态与分子生理学福建省高校重点实验室,福建农林大学, 福州 350002
镉(Cd)是一种人体非必需的元素,具有潜在的“致癌、致畸、致突变”效应,极易通过食物链进入人体并富集,从而危害人体健康[1]。据《全国土壤污染状况调查公报》显示,我国耕地土壤重金属污染点位超标率高达15.5%,其中镉的点位超标率约为7%,居重金属首位[2]。此外,水稻是我国主要粮食作物之一,但有研究表明,部分地区市售大米近10%存在镉超标,已引起人们的高度关注[3]。稻米镉超标与我国农田土壤镉污染比较突出密切相关,利用物理、化学、生物及其复合技术修复镉污染土壤,降低稻米镉含量,将为建立安全的水稻生产系统奠定基础。

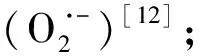
1 材料与方法
1.1 试验材料
供试菌株为镉耐性的假单胞菌(Pseudomonassp.)TCd-1(专利号:CN103952333A)[18],是课题组前期从水稻品种“PI312777”的根际土壤筛选得到,保藏于武汉大学中国典型培养物保藏中心(保存号:CCTCC M2013649)。菌悬液培养条件:牛肉膏0.5%,酵母粉1.0%,氯化镁0.5%,pH 6.3,温度33℃,接菌量1.25%,转速160 rpm,培养24 h后用于接种。接种前取菌悬液100 mL分次在4℃、12000 rpm下离心15 min,去上清液,菌株沉淀物用无菌水稀释至100 mL,稀释后的菌液浓度测定为(5.33±0.24)×1010个/mL。
镉高耐性水稻“特优671”和镉低耐性水稻“百香139”是课题组前期从238份近年我国主栽的水稻品种中筛选得到。供试水稻的种子用2.0%的次氯酸钠溶液浸泡消毒30 min后,置于30℃恒温培养箱培养至种子露白(期间定期补水),然后均匀播于沙盘中,待水稻长至3叶1心期时,选取长势一致水稻用于盆栽试验,每盆移栽水稻秧苗1株。
盆栽土壤取自福建农林大学水稻实验田,土壤镉含量为0.18 mg/kg,土壤经自然风干、研磨,过筛2 mm后,分装于(直径18 cm,高15 cm)的深色塑料盆钵中,每盆2.5 kg,施入氮肥(以纯N计)150 mg/kg、磷肥(以P2O5计)100 mg/kg、钾肥(以K2O计)100 mg/kg,并在盆栽前进行10 mg/kg镉添加处理,土壤混匀,保持水深2—3 cm,每日搅动1次,陈化4周后用于水稻栽培。
1.2 盆栽管理与采样
盆栽试验于2019年3月至8月在福建农林大学生命科学学院玻璃房进行,设置4种处理:(1)CK:不加镉、不接种假单胞菌TCd-1(2)Cd10:添加10 mg/kg镉;(3)CKB:未加镉,接种假单胞菌TCd-1;(4)Cd10B:添加10 mg/kg镉,接种假单胞菌TCd-1。各处理5次重复。在移栽水稻返青后,进行接种,接种量20 mL/kg(即2%)。盆栽期间定期浇水,保持桶里水量高于土面2—3 cm,以保证水稻正常生长。在分蘖期、抽穗期、成熟期分别测定水稻的株高、净光合速率;在抽穗期测定水稻根系活力、叶绿素含量、叶片抗氧化酶活性与抗氧化物质含量、MDA与超氧阴离子含量;在成熟期测定水稻各部位的镉含量。
1.3 测定方法
1.3.1水稻株高、生物量及净光合速率的测定
在水稻分蘖期、抽穗期、成熟期,采用精度0.1 cm的直尺测定水稻株高;在成熟期将水稻全株取出后,用去离子水洗净根部,再用吸水纸吸干,将样品分为根、茎和叶三部分,置于烘箱105℃杀青0.5 h后,转至72℃烘至恒重,测定其干重,生物量为根、茎、叶干重之和,3次重复。净光合速率(Net photosynthetic rate,Pn)采用LI—6400便携式光合测定仪(LI—COR公司,美国)测定,5次重复。
1.3.2根系活力和叶绿素含量测定
根系活力采用四氮唑(TTC)染色法测定[19],结果以TTC还原量(mg h-1g-1FW)表示,测定3次重复。叶绿素含量采用分光光度法测定[20]。
1.3.3抗氧化酶活性测定


1.3.4类黄酮和总酚含量测定
类黄酮含量测定采用甲醇提取和分光光度法[26]。准确称取新鲜去中脉水稻叶片0.5 g,加5 mL含1% HCl的甲醇溶液(甲醇∶水∶盐酸=79∶20∶1),反应30 min后,4℃提取24 h,取0.5 mL提取液,稀释、定容至25 mL,在325 nm处比色测定吸光度(OD)值;总酚含量测定以没食子酸标准曲线,再由280 nm处的OD值计算求得。总酚、类黄酮含量均以鲜重表示。
1.3.5水稻镉含量的测定
镉含量测定利用原子火焰分光光度法进行[27]。在成熟期,将水稻整株取出后,先用自来水冲洗根部土壤,再用去离子水洗净,用吸水纸吸干后,将样品分为根、茎、叶、糙米4个部位,装入信封,置于烘干箱105℃杀青1 h后,72℃烘干48 h,样品用玛瑙研钵研碎,过100目筛,待测。
1.4 数据处理与统计分析
采用Excel 2010和SPSS 22.0软件对数据进行统计分析。采用单因素(one-way ANOVA)和Duncan法进行方差分析和多重比较(P=0.05)。图表中数据为平均值±标准差。
2 结果与分析
2.1 假单胞菌TCd- 1对镉污染水稻各器官镉含量的影响
结果表明,不同处理“特优671”各部位的镉含量高低为:根>茎>叶>糙米,不加镉处理“百香139”表现为:叶>茎>根>糙米,加镉处理的为:茎>根>叶>糙米;水稻不同部位的镉含量在不同处理间均表现为:Cd10>Cd10B>CKB≈CK(表1)。在镉处理下,“特优671”根、茎、叶、糙米的镉含量分别为35.72、9.69、4.34、0.84 mg/kg,接种菌株后依次下降了30.5%、39.1%、40.7%、29.2%(P<0.05);“百香139”的分别为13.18、31.26、6.57、1.33 mg/kg;接种菌株后,依次下降了52.2%、51.7%、18.4%、38.8%(P<0.05)。可见,假单胞菌TCd-1处理后能够显著降低Cd10处理水稻中各器官的镉含量(P<0.05)。

表1 不同处理水稻的镉含量
2.2 假单胞菌TCd-1对镉污染水稻株高和生物量的影响
如图1所示,在分蘖期,CKB处理“特优671”、“百香139”的株高分别为58.70、48.70 cm依次比CK处理提高了7.1%、20.3%;Cd10B处理的株高分别为41.33、35.66 cm,依次比Cd10处理提高了10.6%、17.1%。在抽穗期,CKB处理“特优671”、“百香139”的株高分别为94.19、88.33 cm,依次比CK处理提高了8.4%、15.0%,Cd10B处理的株高分别为88.59、81.89 cm,依次比Cd10处理高10.2%、4.8%。在成熟期,CKB处理“特优671”、“百香139”的株高分别为101.77、98.40 cm,依次比CK处理提高了11.9%、7.5%,Cd10B处理的株高分别为92.18、95.23 cm,依次比Cd10处理高6.7%、12.3%。加菌处理后“特优671”的生物量无显著性差异;CKB、Cd10B处理“百香139”的生物量分别显著增加了29.72%,29.90%(图1)。可见,镉污染抑制了不同镉耐性水稻株高的生长,降低其生物量,接种菌株可显著缓解镉污染的抑制作用。
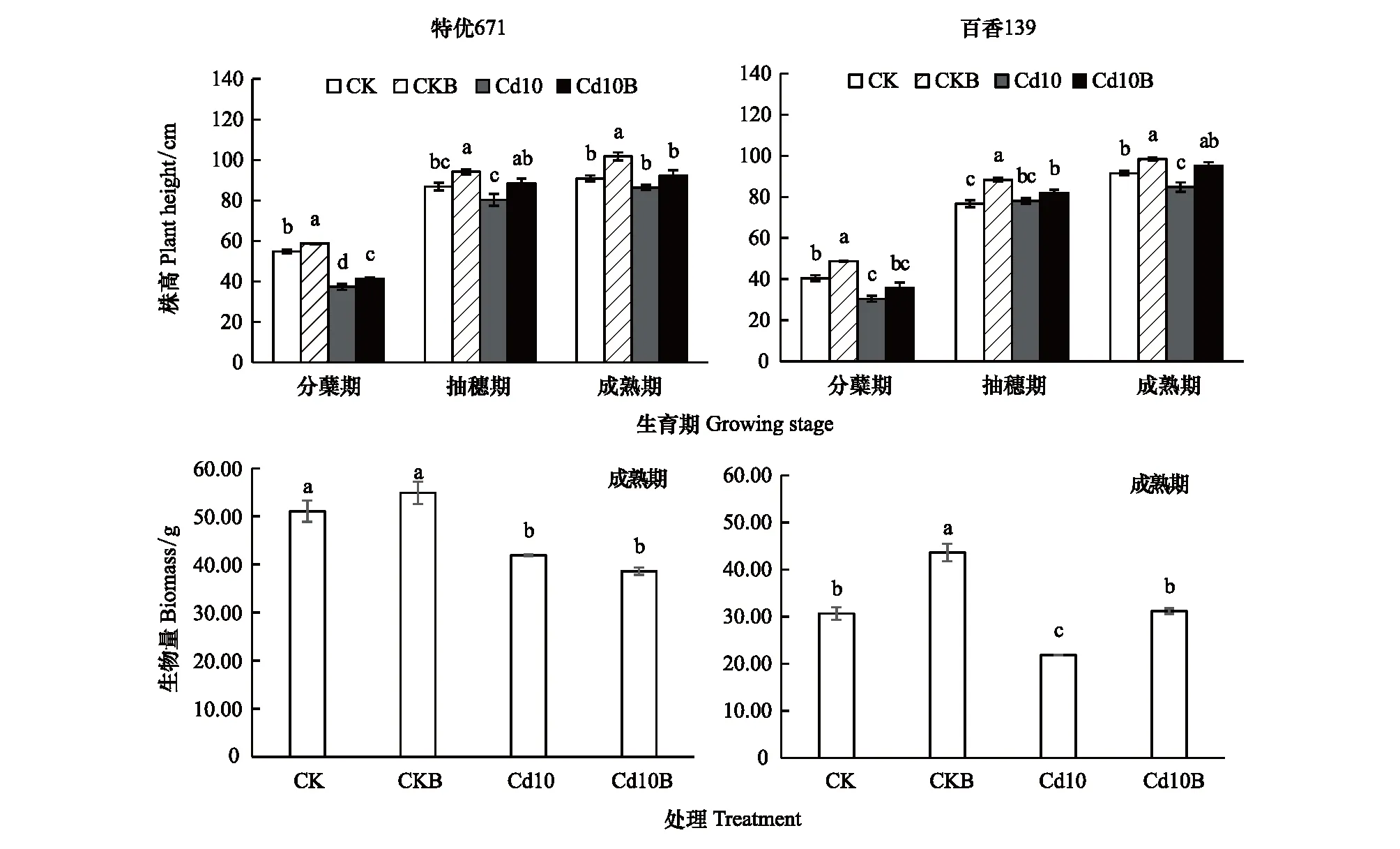
图1 不同处理水稻“特优671”、“百香139”的株高和生物量
2.3 假单胞菌TCd-1对镉污染水稻根系活力和叶片净光合速率(Pn)的影响
CKB处理“特优671”和“百香139”的根系活力分别为0.874、1.196 μg g-1h-1,依次比CK处理提高了9.3%、8.8%;Cd10B处理的根系活力分别为0.779、1.015 μg g-1h-1,依次比Cd10处理提高了3.5%、20.3%(图2)。CKB处理“特优671”和“百香139”的Pn分别为23.1、23.7 μmol m-2s-1,依次比CK处理提高了7.9%、33.1%;Cd10B处理的Pn分别为22.8、17.0 μg g-1h-1,依次比Cd10处理提高了53.0%、63.5%(图2)。可见,镉污染降低了水稻的根系活力和Pn,接种菌株显著提高镉污染水稻的根系活力和光合作用能力。

图2 不同处理水稻的根系活力和净光合速率
2.4 假单胞菌TCd-1对镉污染水稻叶片叶绿素含量的影响
如图3所示,CKB处理“特优671”的叶绿素a(Chlorophyll a,Chla)、叶绿素b(Chlorophyll b,Chlb)含量及叶绿素a/叶绿素b(Chlorophyll a/Chlorophyll a,Chla/Chlb)分别为2.26 mg/g、3.57 mg/g、0.64,依次比CK处理高48.2%、37.0%和8.2%;Cd10B处理“特优671”的Chla、Chlb含量及Chla/Chlb分别为2.23 mg/g、3.34 mg/g、0.67,依次比CK处理高262.1%、224.8%和11.6%。CKB处理“百香139”的Chla、Chlb含量及Chla/Chlb分别为1.53 mg/g、2.79 mg/g、0.55,依次比CK处理高出21.2%、28.6%和-6.2%;Cd10B处理“百香139”的Chla、Chlb含量及Chla/Chlb分别为1.22 mg/g、2.14 mg/g、0.57,依次比CK处理高34.3%、31.8%和2.3%。可见,接种菌株能够显著提高水稻的Chla、Chlb含量并调节Chla/Chlb,缓解镉污染对叶片叶绿素的伤害。

图3 不同处理水稻抽穗期叶绿素a含量、叶绿素b含量、叶绿素a/叶绿素b
2.5 假单胞菌TCd-1对镉污染水稻叶片抗氧化酶系统的影响
“特优671”和“百香139”叶片的MDA含量均以Cd10处理最高,分别为1.58、1.95 μmol/g,CdB10处理的分别为0.97、1.39 μmol/g,依次比Cd10处理降低了38.5%、28.9%,CKB处理与CK处理间无显著差异(图4)。Cd10B处理“特优671”叶片的SOD、POD、CAT活性分别比Cd10处理高出7.3%、82.0%、58.8%,CKB处理的POD、CAT活性比CK处理提高了53.8%、36.8%;Cd10B处理“百香139”叶片的SOD、POD、CAT活性分别比Cd10处理高出138.5%、106.2%、172.7%,CKB处理SOD、POD、CAT活性依次比CK处理高出17.8%、12.5%、29.4%(图4)。可见,Cd10处理显著降低了水稻叶片的抗氧化能力,增强了膜质的过氧化作用,接种菌株表现出显著的缓解镉胁迫效应。
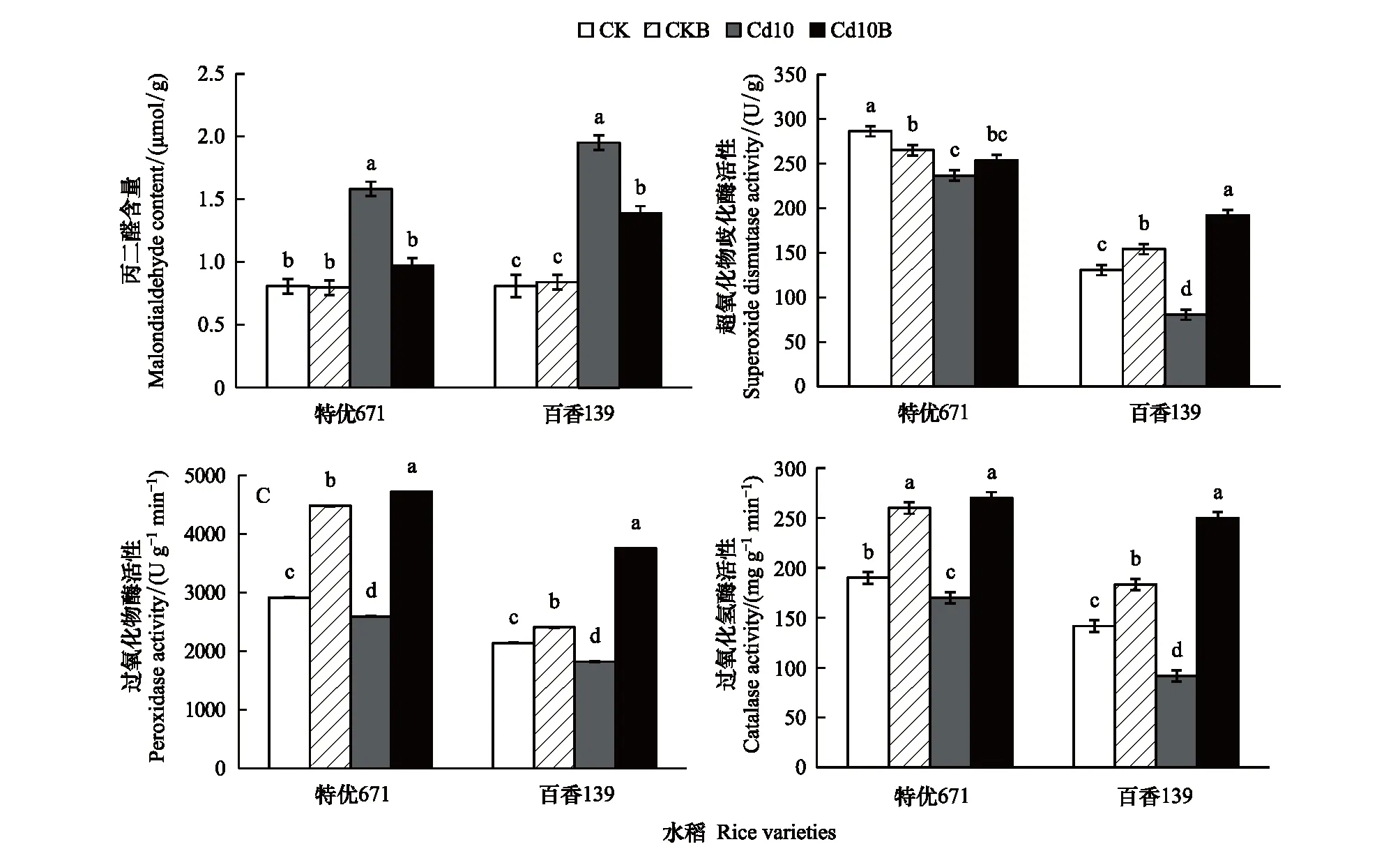
图4 不同处理水稻叶片MDA含量、SOD活性、POD活性、CAT活性
2.6 假单胞菌TCd-1对抽穗期水稻叶片类黄酮、总酚和含量的影响


图5 不同处理水稻抽穗期类黄酮含量、总酚含量含量
3 讨论
3.1 假单胞菌TCd- 1降低了成熟期水稻的镉含量
已有研究表明,一些耐重金属的细菌可通过细胞壁吸附、胞外沉淀和螯合、胞内累积和转化、以及抗氧化系统等作用,缓解重金属污染对植物的毒害作用[5]。刘悦畅等[28]在农田土壤中添加沼泽红假单胞菌(Rhodopseudanonaspalustris)和枯草芽孢杆菌(Bacillussubtilis)菌液,发现添加的微生物能够促进小白菜的生长,其主要作用途径为添加的菌液降低了土壤中镉的生物有效性,小白菜对镉的吸收减少,减轻了镉毒害;李晴晴等[29]发现施用解淀粉芽孢杆菌(Bacillusamyloliquefaciens)FH-1菌剂可显著促进大田种植水稻的生长;陆仲烟等[30]发现伯克氏菌(Burkholderiasp.D54)能显著提高10 mg/L镉处理水稻根的鲜重。赵树民等[31]发现巨大芽孢杆菌(Bacillusmegaterum)LY02对镉和铜污染土壤修复具有作用,能够显著促进黑麦草的生长。本研究发现假单胞菌TCd-1可显著减少不同镉耐性水稻对镉的吸收,这与其分离自水稻根际土壤,能够成功在水稻根际,并且具有极强的镉耐性及镉富集能力有关。同时它是一种具有较强的根际集群能力的植物根际促生菌[17],还可能代谢产生铁载体、分泌植物抗病物质及生长调节物质等,从而调解了水稻的生长[14,32—33]。
3.2 假单胞菌TCd-1通过提高水稻根系活力和光合作用促进镉胁迫水稻的生长
研究表明,镉污染可破坏叶绿素等光合色素的合成,改变Chla/Chlb,造成PSI、PSⅡ和捕光色素—蛋白质复合体的解体或抑制其光合作用蛋白的形成,从而影响光合作用能量传递链上的电子传递等过程[13]。Cd2+进入植物细胞后还能取代叶绿体中存在的Fe2+、Zn2+、Mg2+等二价阳离子,并与叶绿体中相应的蛋白结合,如含—SH的蛋白,导致叶绿体结构破坏,抑制叶绿素前体复合物合成,促进叶绿素降解,降低植物叶绿素含量和光合作用[14]。同时,已有研究表明假单胞菌属的菌株可分泌铁载体与重金属Cd2+、Pb2+和Zn2+等结合形成稳定的复合物,降低土壤中重金属的浓度,缓减重金属对植物根系的毒害作用,促进植株生长[15]。李文华等[34]研究发现,卡伍尔链霉菌TJ430(StrptomycescavourensisTJ430)和荧光假单胞菌P32(PseudomonasfluorescensP32)能显著提高分蘖期水稻的叶绿素含量,降低水稻的镉含量,有利于水稻生长。本研究表明,接种菌株显著提高了在镉污染水稻“特优671”和“百香139”的根系活力、Pn、Chla以及Chlb含量,有效地缓解了镉污染对不同镉耐性水稻的光合抑制作用。这与菌株能够改变根际土壤镉的生物有效性,抑制水稻对镉的吸收、降低根际土壤的镉毒性,减少镉自根系向地上部迁移,进而减轻镉对叶绿体的破坏,提高叶绿素含量,增强了光合作用有关。也可能与菌株通过增强次生代谢物(如植物生长素、解毒物质等)直接或间接促进根系生长、增强根系活力,促进了叶绿素合成,诱导水稻镉抗性增强有关。
3.3 假单胞菌TCd-1通过提高水稻抗氧化系统能力及抗氧物质含量,增强了水稻镉耐性
猜你喜欢
四川劳动保障(2021年3期)2021-06-09 07:08:56
阅读(科学探秘)(2020年8期)2020-11-06 06:22:48
中国果业信息(2019年1期)2019-01-05 17:41:42
现代园艺(2017年13期)2018-01-19 02:28:05
中成药(2017年9期)2017-12-19 13:34:21
生物学教学(2017年9期)2017-08-20 13:22:32
中华老年口腔医学杂志(2016年2期)2017-01-15 14:24:48
西南农业学报(2016年5期)2016-05-17 05:42:36
中国农业文摘-农业工程(2016年5期)2016-04-12 05:38:09
现代检验医学杂志(2015年1期)2015-02-06 01:59:05
