
三个亚热带森林优势种凋落物非结构性碳水化合物含量的季节动态
2022-03-30 10:58:26吴福忠吴秋霞康自佳杨玉盛倪祥银
生态学报 2022年5期
胡 仪,吴福忠,2,3,吴秋霞,康自佳,岳 楷,2,3,杨玉盛,2,3,倪祥银,2,3,*
1 福建师范大学地理科学学院, 福州 350007 2 湿润亚热带生态—地理过程教育部重点实验室, 福州 350007 3 福建三明森林生态系统与全球变化野外科学观测研究站, 三明 365002
碳水化合物是植物光合作用产物,主要以结构性碳水化合物和非结构性碳水化合物(NSC)两种形式存在[1]。NSC主要有可溶性糖和淀粉,是植物代谢的重要物质基础和能量的主要储存形式,参与植物的物质代谢和信号传导[2]。NSC的含量通常可以反应植物整体的碳供应状况[3],体现树木阶段性碳获取和碳支出的平衡关系[4]。植物叶和枝等器官在完成代谢活动后枯萎掉落,以凋落物的形式归还土壤[5—6]。凋落物作为森林养分循环的重要载体,通过分解过程将植物中部分有机物和养分归还土壤[7—8],是森林土壤有机质形成的重要来源[9],也是森林生态系统有机碳和养分的重要储存库[10]。同时,凋落物初始养分含量决定了分解过程中养分释放速率和归还量,而且凋落物分解速率和养分释放量在很大程度上影响着森林土壤有效养分的供应状况、植物养分吸收策略,进而影响整个生态系统养分循环过程[11]。过去几十年来,诸多学者对森林凋落物归还动态、养分含量、生态化学计量及其对凋落物分解的影响进行了大量的研究,在元素(C、N、P、K、Ca、Mg等大量元素和Fe、Mn、Zn、Cu等微量元素)归还、结构性碳水化合物(木质素、纤维素等)降解过程等方面形成了较为系统的认识[11—16]。然而,凋落物中可溶性糖、淀粉等NSC作为易分解组分,可以在凋落物分解早期阶段快速释放,并被土壤微生物快速利用参与土壤生物地球化学循环过程。因此,充分认识新鲜凋落物(分解初始阶段)中NSC含量动态规律,是深刻揭示森林生态系统碳和养分循环的关键之一。
叶片是植物光合作用的主要器官,是树木重要的碳源[17],指示着树木的生理状况和碳供应水平。枝连接树木的碳供应器官和碳需求器官,源源不断地维持着营养物质从叶向树干和根部的运输[18]。同时,叶和枝是凋落物节律性归还的主要形式。研究发现,在叶、枝、花、果等不同类型的凋落物中,凋落叶占年凋落物总量的半数以上(所占比例为51.4%—65.1%);枝次之,约占凋落物总量的4.7%—26.3%[19]。这些凋落叶和枝所携带的易分解碳和养分是土壤微生物的主要碳和养分来源,并参与微生物代谢和土壤有机质形成[20]。因此,研究凋落物节律性归还过程中新鲜凋落叶和枝中NSC含量动态规律对深入认识森林土壤生物地球化学循环过程至关重要。
我国的亚热带地区分布着北半球同纬度地区面积最大的湿润季风性亚热带常绿阔叶林。然而,近几十年来,大面积地带性常绿阔叶林逐渐转化为人工林(主要是杉木、马尾松等),对山地生态系统的结构和功能产生了深刻影响[21]。亚热带水热丰沛,淋溶强烈,凋落物中NSC等易分解组分可快速释放进入土壤并参与生物地球化学循环过程。然而,现在的关键科学问题是:亚热带大面积的地带性常绿阔叶林转换为杉木和马尾松人工林后,凋落物NSC含量及其季节动态是否发生了显著改变并不清晰。我们提出以下假设:1)叶作为光合产物的主要合成场所,凋落叶中NSC含量高于凋落枝;2)杉木和马尾松凋落叶和枝中NSC含量低于地带性常绿阔叶树种米槠。因此,本研究以亚热带常绿阔叶林优势树种米槠(Castanopsiscarlesii)和典型造林树种杉木(Cunninghamialanceolata)、马尾松(Pinusmassoniana)为研究对象,逐月收集这三个树种新鲜凋落叶和凋落枝,并分析其NSC(可溶性糖和淀粉)含量,以期为亚热带森林可持续经营提供一定的理论依据。
1 材料与方法
1.1 研究样地概况
研究样地位于福建三明森林生态系统与全球变化野外科学观测研究站(26°11′N,117°28′E)。该区域以低山为主,平均海拔约300 m,坡度约25—45°。气候类型为亚热带季风气候,年均气温19.4 ℃,年均降水量1700 mm,降雨主要集中在3—8月[22]。土壤为花岗岩发育的红壤,平均容重为1.04 g/cm3。地带性植被为亚热带常绿阔叶林,优势树种为米槠、木荷(Schimasuperba)、格氏栲(Castanopisskawakamii)等。但从1958年开始,大面积天然林被砍伐以营造人工林,主要是杉木和马尾松等。目前,该地区人工林面积约占森林覆盖面积的47.5%[23]。本研究选取1976年经强度择伐后自然演替形成的米槠次生林和同年由米槠天然林经皆伐、炼山后种植形成的杉木、马尾松人工林为研究对象,3个样地相距约800 m,样地基本信息见表1。

表1 3种林分样地信息
1.2 样品采集
在米槠次生林、杉木人工林、马尾松人工林上、中、趾坡位置分别建立1个20 m × 20 m的样地作为重复,各重复样地坡向、坡度、冠层相似。每个样地随机布设5个0.7 m × 0.7 m的尼龙网质凋落物收集框(孔径为1 mm,距离地面0.5 m)。于2013年7月至2014年6月期间,每月(雨季每半月收集,防止淋溶流失)收集新鲜凋落物,并筛选出目标树种(米槠、杉木、马尾松)的新鲜凋落叶和凋落枝,分别置于80 ℃烘箱烘干至恒重,粉碎,过80目筛待测。
1.3 样品分析
NSC(可溶性糖和淀粉)含量采用苯酚浓硫酸法进行测定[24]:
可溶性糖的提取:称取60 mg样品,加入10 mL 80%乙醇,静置萃取24小时后以4000 r/min离心10 min,将上清液转入100 mL容量瓶,再在剩余残留物中加入5 mL 80%乙醇,离心5 min,获取上清液,反复3次,并将所有上清液转入容量瓶定容,装入离心管,于4 ℃保存待测。
淀粉的提取:将上述提取后的残余物于65 ℃烘干,加入10 mL蒸馏水,混匀后放置在沸水浴中糊化15 min,冷却至60 ℃,加入1 mL 0.5% α-淀粉酶,于60 ℃水浴锅保温1 h,使淀粉充分分解,之后将其加热至沸腾使酶失活。再将其2000 r/min离心5 min,过滤定容至100 mL,装入离心管,于4 ℃保存待测。
可溶性糖和淀粉含量的测定:取1 mL的待测液放入试管,加入1 mL溶于80%乙醇的28%苯酚,然后立即加入5 mL浓硫酸,用涡旋仪振荡15 s,静置15 min后于490 nm比色,测定吸光值,并按蔗糖标准曲线转换为可溶性糖和淀粉的百分比含量(%),即100 g凋落物中所含有的可溶性糖或淀粉含量,NSC含量为可溶性糖和淀粉含量之和[25]。
水溶性氮、水溶性磷、总氮含量的测定:
精确称取0.5000 g干样,放入150 mL锥形瓶中,加入50 mL蒸馏水,常温下震荡30 min,过0.45 μm滤膜,即为待测液。水溶性氮(water-soluble nitrogen, WSN)含量采用凯氏定氮法测定,水溶性磷(water-soluble phosphorus, WSP)含量采用钼锑抗比色法测定[26]。总氮(total nitrogen, TN)采用元素分析仪测定。
1.4 数据分析
采用重复测量方差分析(ANOVA)检验不同树种和凋落物类型在时间序列上对NSC、可溶性糖、淀粉含量的影响。采用Tukey多重比较检验NSC、可溶性糖、淀粉含量在同一时期、不同树种间的差异显著性。采用重复测量方差分析分别对凋落叶和枝检验不同树种在时间序列上对NSC、可溶性糖、淀粉含量的影响。采用Pearson相关分析检验凋落物中NSC含量与水溶性氮(WSN)、水溶性磷(WSP)、总氮(TN)、碳氮比(C/N)的相关性。
2 结果
2.1 3个树种凋落物NSC含量季节变化
3个树种凋落叶中NSC含量为2.18%—5.56%,且都具有明显的时间动态(P=0.027,图1)。米槠凋落叶中NSC含量平均值为4.35%,在6—9月持续上升(3.37%—5.56%),9月至次年1月间波动幅度较大(3.03%—5.57%),其他月份较为稳定(4.08%—4.69%)。杉木凋落叶NSC含量平均值为3.24%,在10月至次年4月保持较高水平(3.44%—4.32%),且在1月达到全年最高值,5—9月含量较低(2.18%—2.56%)。马尾松凋落叶中NSC含量平均值为4.01%,在7—8月达全年高值(分别为4.89%和4.68%),其它月份在3.85%左右。然而,总体上来看,凋落叶中NSC含量在3个树种间存在显著差异,其中,米槠和马尾松凋落叶中NSC含量均显著高于杉木(表2)。

图1 米槠、杉木和马尾松新鲜凋落叶和枝中NSC含量的时间动态
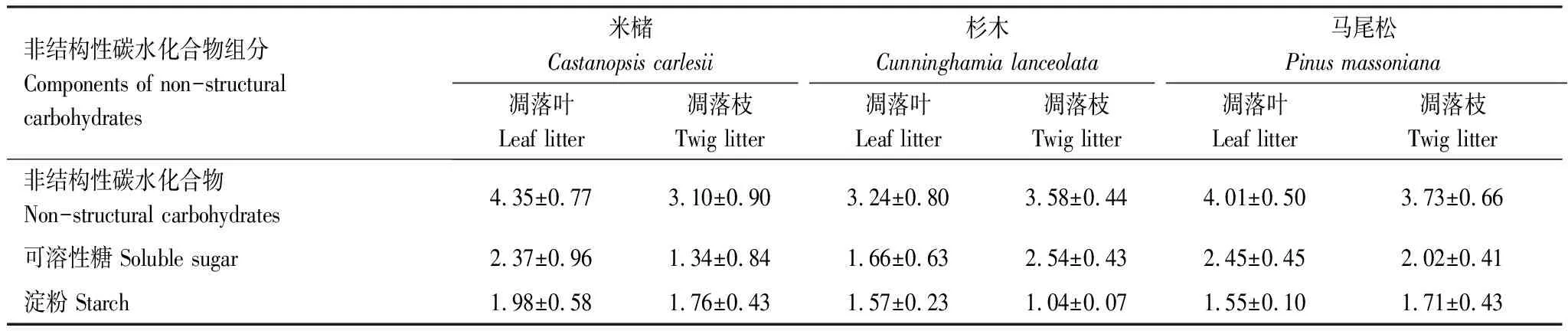
表2 3个树种凋落物中NSC、可溶性糖和淀粉含量的平均值/%
3个树种凋落枝中NSC含量为1.87%—5.27%,总体均显著低于其凋落叶(P=0.004,表3)。其中,米槠凋落枝中NSC含量平均值为3.10%,具有明显的时间动态,在9月至次年1月间波动幅度较大(1.87%—4.22%),在2—6月和7—9月较为稳定,分别保持在3.69%—3.95%和1.87%—2.19%。杉木和马尾松凋落枝中NSC含量时间动态不显著,杉木全年维持在其平均值(3.58%)左右,而马尾松在1月达全年最大值(5.27%),其它月份在2.75%—4.44%间波动,全年平均值为3.73%,显著高于米槠凋落枝中NSC含量。

表3 凋落物类型和树种对NSC、可溶性糖和淀粉含量的重复测量方差分析
2.2 3个树种凋落物可溶性糖含量季节变化
3个树种凋落叶中可溶性糖含量为0.97%—3.85%(图2),且具有明显的时间动态。其变化规律与NSC含量变化规律大体一致,且米槠和马尾松凋落叶中可溶性糖含量也显著高于杉木(米槠、杉木、马尾松凋落叶中可溶性糖含量平均值分别为2.37%、1.66%、2.45%)。
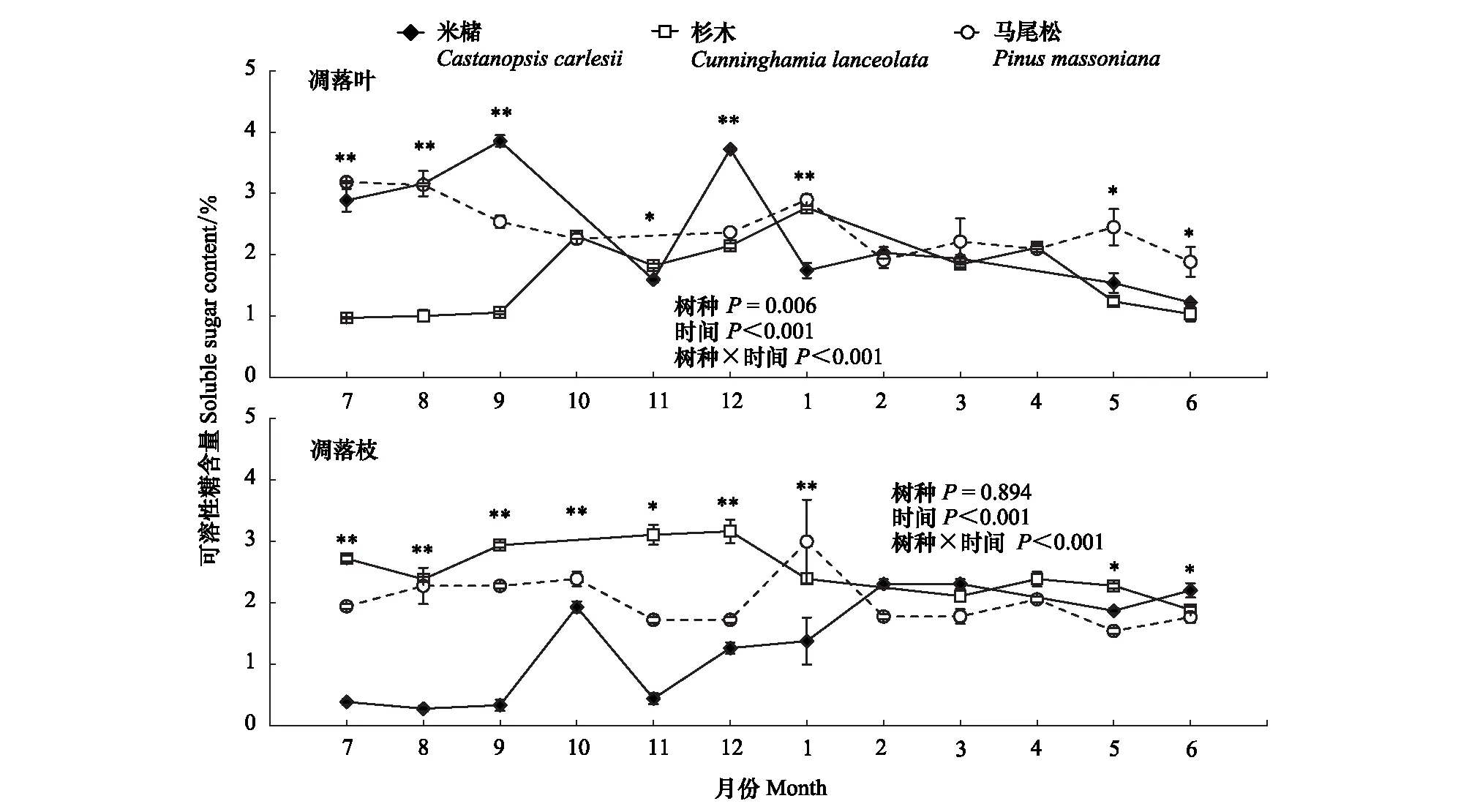
图2 米槠、杉木和马尾松新鲜凋落叶和枝中可溶性糖含量的时间动态
3个树种凋落枝中可溶性糖含量为0.28%—3.16%,与凋落叶中可溶性糖含量无显著差异(P=0.082),且米槠和杉木凋落枝中可溶性糖含量具有明显的时间动态。米槠凋落枝中可溶性糖含量平均值为1.34%,在11月至次年2月持续上升(0.44%—2.31%),2—6月和7—9月分别处在全年最高值(1.87%—2.31%)和最低值(0.28%—0.38%)。杉木凋落枝中可溶性糖含量平均值为2.54%,9—12月处于全年较高值(2.94%—3.16%),1—8月处于全年较低值(1.89%—2.72%),但总体波动范围较小。马尾松凋落枝中可溶性糖含量平均值为2.02%,除在1月较高外,其余月份均保持在1.97%左右,无明显时间动态。同时,凋落枝中可溶性糖含量在3个树种间差异显著,总体表现为杉木>马尾松>米槠。
2.3 3个树种凋落物淀粉含量季节变化
3个树种凋落叶中淀粉含量为1.21%—2.75%(图3),且米槠和杉木凋落叶中淀粉含量存在显著的时间动态。米槠凋落叶中淀粉含量平均值为1.98%,在7—12月间相对稳定,保持在1.38%—1.71%,其余月份含量较高且波动较大。杉木凋落叶中淀粉含量平均值为1.57%,在7—11月持续上升,11月至次年6月小范围缓慢波动(1.32%—1.87%)。马尾松凋落叶中淀粉含量平均值为1.55%,全年波动幅度小,维持在1.57%左右。且树种间凋落叶中淀粉含量差异显著,米槠显著高于杉木和马尾松。

图3 米槠、杉木和马尾松新鲜凋落叶和枝中淀粉含量的时间动态
3个树种凋落枝中淀粉含量为0.94%—2.96%,显著低于凋落叶中淀粉含量(P=0.038),且3个树种凋落枝中淀粉含量均无明显时间动态。米槠凋落枝中淀粉含量平均值为1.76%,在12月达全年最大值(2.96%),其余月份均在1.38%—1.93%小幅度波动。杉木凋落枝中淀粉含量较为稳定,全年保持在小范围内波动(0.94%—1.17%),平均值为1.04%。马尾松凋落枝中淀粉含量平均值为1.71%,在8—11月持续上升(1.33%—2.55%),2—4月总体呈下降趋势(2.27%—1.21%)。凋落枝中淀粉含量在不同树种间差异显著,米槠和马尾松均高于杉木。
2.4 凋落物中NSC与其它组分的相关关系
线性相关分析结果表明,凋落叶中NSC含量与C/N呈显著负相关关系(r=-0.335,P<0.01)。凋落枝中NSC含量与WSN(r=-0.142,P=0.185)、WSP(r=-0.047,P=0.658)、TN(r=-0.087,P=0.392)、C/N(r=-0.164,P=0.106)均无显著相关关系(图4)。

图4 凋落叶和枝中NSC含量与其它组分的相关关系
3 讨论
本研究通过比较亚热带常绿阔叶林优势树种米槠和主要造林树种杉木和马尾松新鲜凋落叶和枝中NSC含量发现,森林转换后,树木生理特性和凋落物节律特征显著影响新鲜凋落叶和枝中NSC含量。其中,米槠和马尾松凋落叶中NSC含量显著高于凋落枝,与本文中假设1相符,而杉木凋落叶中NSC含量显著低于凋落枝,可能是源于其自身生物学特性。树种间凋落物NSC含量差异显著,但与假设2不完全相同,其中米槠和马尾松凋落叶中NSC含量显著高于杉木,而马尾松凋落枝中NSC含量显著高于米槠。
凋落物中NSC及其组分含量具有明显的时间动态。树木中NSC的积累是其为应对生长和环境变化而形成的动态过程,因此NSC及其组分含量的变化受到树木节律和季节变化的共同影响[3,27]。由于米槠、杉木和马尾松的叶性状、物候等种间差异,3个树种凋落物NSC含量呈现出复杂的动态变化。总体上,米槠、杉木和马尾松凋落叶和凋落枝中NSC含量在11—12月呈上升趋势,可能是由于期间雨量较少,养分不易淋失。而2—6月正值雨季,树木凋落叶和枝中NSC含量下降,一部分原因是可溶性糖随着强烈的雨水淋溶而流失,另一个原因可能是春季萌芽生长,老叶凋落(亚热带很多树种凋落物高峰期发生在春季),并在凋落前将部分养分转移到嫩叶中,致使衰老叶中含量相对较低[28]。5月以后,树木处于径向快速生长阶段,叶片中大量光合产物被输送到枝、树干等器官,以供木质部的快速生长,从而使该阶段凋落叶和枝中NSC含量减少[29]。同时,可溶性糖是由易溶于水的单糖和寡聚糖组成,可为植物直接供能,而淀粉是葡萄糖分子聚合而成的多聚体,常温下不能溶解,是植物的重要储能物质[30—31]。在热量充足条件下,树木不需要储存较多的淀粉来抵御低温,而是倾向于以可溶性糖(可直接利用)的形式来存储碳源。因此,米槠、杉木和马尾松中NSC以可溶性糖为主,淀粉含量较少,且受降水影响,可溶性糖和淀粉的溶解性质差异导致了可溶性糖含量的年际波动范围较大,而淀粉含量全年波动幅度较小,与张异平等人[29]的研究相符。
不同类型的凋落物NSC及其组分含量差异显著。据刘万德等人[32]研究表明,器官间的NSC含量差异主要源于各器官的功能差异。植物叶片作为植物光合作用的主要器官,产生大量的糖分供植物进行生理代谢活动,但其中大部分被输送到植物的其它部位以供植物的生长和新陈代谢,只有少量过剩的可溶性糖转化为淀粉,临时储存在叶片中,因此叶中淀粉含量较低。同时叶片自身的较高代谢速率要求保证正常的细胞膨胀压,这就使得叶片中NSC尤其是可溶性糖含量相对较高。枝是叶片向树根输送养分的通道,在树体内起到传输碳水化合物的作用[18],通常情况下NSC含量波动范围较小[33]。本研究结果表明,凋落叶中可溶性糖含量高于凋落枝,凋落叶中淀粉含量与凋落枝中淀粉含量相差较少,正好与此研究结果相符。
不同树种间的NSC及其组分含量存在显著差异。总体上,杉木凋落叶中NSC及其组分含量以及马尾松凋落叶中淀粉含量均显著低于米槠凋落叶,杉木凋落枝中可溶性糖含量显著高于米槠但淀粉含量显著低于米槠凋落枝,马尾松凋落枝中NSC和可溶性糖含量显著高于米槠凋落枝。这种差异的根本原因在于不同树种的生物学特性差异[30]。在亚热带季风气候背景下,各树种根据其自身特性采取不同的生长策略来适应水热等环境条件以提高生存能力,而NSC尤其是可溶性糖,在调节植物代谢活动(特别是环境胁迫条件下)发挥着重要功能。因此,不同树种间凋落叶和枝中NSC及其组分含量具有显著差异。同时,米槠次生林转化为杉木和马尾松人工林后,凋落物节律发生了很大改变。据统计,米槠凋落物量在4月达到峰值,其他月份凋落量均较少,而杉木和马尾松凋落物量全年变化不大,但存在多个高峰期。樊后保等人[34]研究表明,凋落物养分归还有明显的时间动态,且森林转化后,同一时期的NSC及其组分含量和凋落量都发生改变,因此归还到土壤表面的NSC等易分解组分含量也随之发生巨大改变。这些养分进入土壤后可快速释放并参与土壤微生物周转和土壤有机质形成[20],并深刻影响森林土壤生物地球化学循环过程。因此,在造林过程中,可适当选择凋落物中NSC等易分解组分含量较高的树种,促进森林生态系统中以凋落物为载体的碳和养分循环,促进亚热带人工林可持续经营。
4 结论
凋落叶和枝中NSC含量均有明显时间动态,这与树木物候节律以及环境因素密切相关。不同类型的凋落物NSC含量存在显著差异,米槠、杉木和马尾松凋落叶中NSC含量分别为3.03%—3.56%、2.18%—4.37%、3.38%—4.89%,凋落枝中NSC含量分别为1.87%—4.22%、2.88%—4.28%、2.75%—5.27%。米槠和马尾松凋落叶中NSC含量高于凋落枝,而杉木凋落枝中NSC含量高于凋落叶。不同树种间凋落物中NSC含量差异显著,米槠和马尾松凋落叶中NSC含量显著高于杉木,而马尾松凋落枝中NSC含量显著高于米槠。可见,亚热带地带性常绿阔叶林转化为杉木、马尾松人工林后,凋落物中NSC含量的动态规律也随之发生了显著改变,进而对凋落物中养分归还和森林土壤生物地球化学过程产生影响。因此,深入探讨不同树种凋落物NSC含量动态规律及其与养分的耦合关系,结合NSC等易分解组分及其参与的森林土壤生物地球化学过程,可为亚热带造林过程中树种的选择提供一定的理论依据,以期更好地实现亚热带人工林可持续经营。
致谢:感谢福建师范大学三明站工作人员在野外采样工作中给予的帮助,感谢郭鸿蓉、朱玲、吴若冰、魏文涛在室内分析工作中提供的帮助。
猜你喜欢
农民致富之友(2020年8期)2020-05-11 05:54:38
绿色科技(2019年5期)2019-11-29 13:17:37
现代园艺(2017年23期)2018-01-18 06:58:24
现代园艺(2017年23期)2018-01-18 06:58:19
中成药(2016年4期)2016-05-17 06:07:52
现代食品(2016年14期)2016-04-28 08:10:38
广西林业科学(2016年2期)2016-03-20 05:53:20
广西林业科学(2016年1期)2016-03-20 05:33:00
广西林业科学(2016年1期)2016-03-20 05:32:59
广西林业科学(2016年1期)2016-03-20 05:32:58
