
不同坡向川西亚高山林木竞争与叶片表型可塑性的关系研究
2022-03-30 10:58张小晶李巧玉陶建平
生态学报 2022年5期
陈 娟,张小晶,李巧玉,陶建平,*
1 三峡库区生态环境教育部重点实验室, 重庆 400715 2 重庆市三峡库区植物生态与资源重点实验室, 重庆 400715 3 西南大学生命科学学院, 重庆 400715
表型可塑性是指植物同一基因型在不同环境下产生不同表型的特性[1]。近年来,关于叶片功能性状表型可塑性变化研究得到广泛关注[2—6]。由于与外界环境接触面积最大,叶片对外界环境变化十分敏感,是联系植物与环境的重要纽带[7—9],因此也是功能性状的重要构成[10—13]。叶片性状变化可以影响植物与生长环境间的物质能量交换,揭示植物对变化环境的响应和适应[8],反映植物对资源获取和利用策略的差异[14]。因此探索植物叶片性状的表型可塑性对解释植物对不同环境的适应具有重要的生态学意义。
有研究认为,竞争对植物适应性的影响作用大小随着环境条件有利性的增加而增加[15—18]。接受这一假设相当于将性状引入生物量梯度上物种多样性分布的驼背模型[17]。该模型认为物种多样性大小与环境压力限制程度有关[17]。在环境条件利于植物生长时,具有高竞争能力的物种获得最大活力,抑制低竞争能力的物种生长,此时物种多样性较低。而随着环境压力增加,竞争物种的活力下降,而竞争能力较低的物种能够生存,但是物种多样性受到极端环境压力限制而下降。因此认为最大的物种多样性发生在环境压力梯度的中等范围。因此,竞争对性状多样性的影响作用大小很可能随着环境压力梯度的变化而变化。Erwin 等人通过模拟氮损失,发现在极端恶劣的环境中,两种苔属植物间的相互作用减弱,植物功能多样性受到限制,此时植物表现出与环境逆境相关的性状,叶片性状表型相似,可塑性较低[19]。但是,在自然森林群落中,群落生物环境和非生物环境更为复杂,成熟的林木其叶片性状变化可能更具有保守性[20—22]。而环境资源分布在自然森林群落中常常存在异质性,尤其是坡向变化使得群落环境资源差异巨大[23]。不同坡向之间虽然距离较短,但是水、热、光照的不同组合,影响不同坡向上群落资源条件[23]。周志强等人已经发现坡向变化影响了林木的种间竞争强度[24]。但是少有研究探讨自然森林群落中,在不同环境资源条件下竞争对植物叶片性状表型可塑性影响作用大小是否变化。
川西地区是青藏高原向四川盆地的过渡区域,海拔变化剧烈,为典型的高山峡谷区。在不同坡向上,环境资源条件差异较大,植被差异十分明显。因此川西地区丰富多样的植被为研究不同坡向上环境因素和竞争作用对叶片表型可塑性变化的影响提供了很好的条件。本研究以四川省阿坝藏族羌族自治州位于同一海拔梯度但坡向相反的天然次生林为研究对象,评估了不同坡向上资源条件和竞争强度,分析了不同坡向上竞争强度与叶片功能性状表型可塑性的关系的差异以及叶片表型可塑性随着个体大小增加的变化,以阐明不同坡向上叶片表型可塑性的驱动因子,以及群落内部的竞争格局以及功能性状的变异特征,为理解川西亚高山地区植物对环境的适应性策略提供理论依据。
1 研究地区与研究方法
1.1 研究区域概况
研究地点位于四川省阿坝藏族羌族自治州理县古尔沟(N31.51°,E102.90°),海拔大约2400 m。年平均气温约12℃,年平均降雨量584 mm。由于受到高山地形的影响,该地气候为冬寒夏凉的高山气候。该区域是典型的高山峡谷地貌,地形以石质山地为主,坡度多在25°以上。植被以次生的落叶阔叶林为主,主要优势种有青榨槭(Acerdavidii),五裂槭(Aceroliverianum),橿子栎(Quercusbaronii),黄栌(Cotinuscoggygria),川梣(Fraxinussargentiana),华山松(Pinusarmandii)等。林下灌木主要有川桂(Cinnamomumwilsonii),菝葜(Smilaxchina),草本层主要有苔草(Carexspp.),唐松草(Thalictrumaquilegifolium)等。当地森林主要土壤类型是山地暗棕壤,土层浅薄;成土母质为白云岩、板岩等风化物残留,土壤中石砾含量较高。
1.2 试验设计
选取研究区域内人为干扰最小,位于同一海拔梯度但坡向相反的天然次生林作为研究区域设置样地。在位于不同坡向的林内分别设置面积为2500 m2的样方3个,共6个。对于每个样方内总株数大于5的树种选定为研究目标物种,树木胸径 ≥ 2 cm 的设定为对象木,记录对象木的胸径、树高以及冠幅,并且进行挂牌编号。以树冠接触和林木的遮荫状况来确定竞争范围,竞争范围内的个体为竞争木,实测竞争范围内对象木与竞争木之间的距离。
记录样地中心海拔、坡度、坡向等环境指标,在每个样地内选取5个点,2018年8月用HH2土壤湿度计测仪测量土壤温度和土壤湿度,用Hyelec温湿计测量大气温度和大气湿度,基本信息见表1。

表1 样地基本概况
1.3 测定项目与方法
1.3.1植物叶片功能性状测定
于生长季(7月末8月初)收集阴坡和阳坡样方内对象木的冠层中部外侧成熟、完整、发育正常、无病虫害的叶片。为减少水分蒸发及叶形的变化,将刚采集的叶片立即装入编号自封袋(其号码与对象木编号对应)密封保鲜保存,进行叶性状的测量。
叶片功能性状指标包括比叶面积(SLA),叶干物质含量(LDMC),叶片全碳含量(C),叶片全氮含量(N),叶片全磷含量(P),叶片全钾含量(K),叶片全镁含量(Mg),叶片碳氮比(CNR),叶片氮磷比(NPR),叶片碳磷比(CPR)共10种叶片功能性状。测量方法按照功能性状标准测量手册测量[25],具体测定方法如下:
LDMC测定:LDMC是叶片干重(mg)与叶片饱和鲜重(g)的比值,单位mg/g。
SLA测定:计算公式如下:
(1)
LA指新鲜叶片单面面积(Leaf area),LDMC为叶片干物质含量。叶片面积采用扫描仪(EPSON V19)扫描,得到扫描叶片照片,然后采用叶面积软件得到单叶叶面积值。根据公式计算即得到SLA。
叶片元素含量测定:叶片元素含量包括叶片全碳、叶片全氮、叶片全磷、叶片全钾、叶片全镁。将80℃烘箱烘干48 h后的叶片,使用球磨仪研磨叶片,使叶片元素均一化,然后将叶片粉末过0.25 mm筛。使用元素分析仪(CHNO-Vario EL cube, Heraeus Elemental, Hanau, Germany)测定叶片全碳和叶片全氮,使用微波消解仪(SpeedWave MWS-4)消解叶片,采用电感耦合等离子体发射光谱法(ICP-OES, Thermp Fisher Icap 6300)测定研究所选叶片的其它元素。
1.3.2土壤元素测定
7月末收集阴坡和阳坡样地土壤样品。使用环刀采用五点法取样,取得土壤表层土(0—10 cm),每个样方3个重复,共30份土样,带回实验室测定土壤元素。土壤元素含量包括土壤全碳、土壤全氮、土壤全磷、土壤全钾、土壤全镁。将土壤置于80℃烘箱持续烘干48 h,用球磨仪将土壤研磨成粉末,过0.25 mm筛,使土壤元素均一化,然后用于土壤元素含量的测定。使用元素分析仪(CHNO-Vario EL cube, Heraeus Elemental, Hanau, Germany)测定土壤全碳和全氮,用微波消解仪(Speed Wave MWS-4)进行消解,然后采用电感耦合等离子体发射光谱法(ICP-OES,Thermp Fisher Icap 6300)测定。
样地土壤元素含量信息如下:
1.4 数据处理
1.4.1竞争强度计算
本文采用张跃西(1993)改进的Hegyi单木竞争模型来计算竞争指数来评估林木间竞争强度。其计算公式为:
(2)
其中,CI为竞争指数,该值越大,竞争强度越大;Di为竞争木胸径;D为对象木胸径;不同为对象木与竞争木之间的距离;N为竞争木的株数。当竞争木与对象木相同时,其林木竞争强度定义为种内竞争强度,当竞争木与对象木不同时,其竞争强度定义为种间竞争强度。种内竞争强度和种间竞争强度分开计算。研究中先计算出每棵竞争木对对象木的竞争指数,然后将N棵竞争木对该对象木的竞争指数累加,其和为该对象木的竞争强度。通过计算样方内所有对象木的平均种内竞争强度、种间竞争强度以及总竞争强度综合评价不同坡向上群落内林木竞争状况。
1.4.2群落加权性状均值计算
按照Gainier 等(2004)提出的计算样地群落内每个物种相对丰度加权后的性状均值,也就是群落加权性状均值(Community-weighted mean trait value,CWM),计算公式如下:
(3)
其中,CWM表示指定功能性状的群落加权性状均值,pi表示物种i的相对丰度(i= 1,2,3 ……S),xi是物种i的性状均值。
1.4.3性状变异性计算
利用收集的阴坡和阳坡样方内所有对象木的叶片性状值,计算阴坡与阳坡上群落的性状变异系数(Coefficient of variation,CV)值。使用自助法(Bootstrap)重复抽样1000次,获得阴坡和阳坡群落叶片性状变异系数(CV)分布以及95%置信区间,来比较阴坡和阳坡群落叶片性状变异系数是否具有差异。利用R中boot包中的boot函数完成抽样[26]。CV值计算公式如下:
其中CV表示性状数据的变异系数,sd为标准偏差,Mean为性状均值。
1.4.4数据分析
为了分析个体大小等级下的叶片性状表型可塑性,将胸径、树高、冠幅、胸径×树高×冠幅分别作为个体大小指标,将胸径、树高、冠幅、胸径×树高×冠幅分别单位Z分数标准化,按照每一等级下林木数量大致相等的原则划分3个等级,等级标准如下:

表3 林木个体大小等级
按照以上标准分别划分个体大小等级,共有胸径、树高、冠幅、胸径×树高×冠幅四个个体大小指标。比如对于胸径,将不同坡向上样方内的林木按照胸径标准划为1,2,3个等级,针对每个等级内的林木重复抽样,每次抽样完成计算叶片性状在该等级下的所有的叶片性状变异系数CV值,然后取其均值,重复抽样次数100。然后用单因素方差分析比较不同等级下的性状变异系数CV值是否具有显著性差异。
利用线性混合模型分别分析竞争强度与林木个体的胸径、树高、冠幅和胸径×树高×冠幅的关系。在模型中,竞争强度为因变量,胸径、树高、冠幅或者胸径×树高×冠幅为固定效应,物种作为随机效应。R version 3.6.1[27]用来完成本实验中所有数据分析和作图,其中线性混合模型用lme4 包来进行拟合[28]。
2 结果与分析
2.1 不同坡向林木叶片性状表型可塑性
结果显示,研究所选的10种叶片性状中有8种性状(包括比叶面积、叶碳含量、叶氮含量、叶磷含量、叶钾含量、碳磷比、碳氮比、氮磷比)显示阴坡上叶片性状表型可塑性(CV值)大于阳坡,且置信区间不重叠或重叠较少。但是林木叶片叶干物质含量、叶镁含量的表型可塑性在阴坡和阳坡上差异不大,样地林木叶片CV值的置信区间重叠(图1)。高可塑性反映了植物在干扰环境下的高生存能力,本文推测阴坡上植物更容易调节林木叶片比叶面积、碳、氮、磷、钾含量来适应群落中的变化环境,提高植物的生存能力。

图1 阴坡和阳坡上叶片性状表型可塑性
2.2 不同坡向林木竞争强度
比较阴坡和阳坡上林木平均种内竞争强度,种间竞争强度以及总竞争强度,结果显示阴坡上林木平均种内竞争强度、种间竞争强度以及总竞争强度都大于阳坡(表4),其值分别为79.41,63.29和132.19。这可能是因为阴坡上植物冠幅较大,植物生长受到邻近植物的影响较大,从而受到较大的竞争压力。

表4 阴坡和阳坡上群落林木竞争强度(±标准偏差)
2.3 不同坡向林木个体大小与叶片表型可塑性的关系
研究中选择了胸径、树高、冠幅、胸径×树高×冠幅共四种个体大小等级指标。比较阴坡和阳坡上不同个体大小等级下林木叶片性状表型可塑性的变化,结果显示阴坡上随着林木胸径、树高、冠幅、胸径×树高×冠幅的增加,林木叶片性状CV值增加,第1等级和第3等级组间具有显著性。在阳坡上随着胸径的增加,叶片性状表型可塑性显著增加,但是随着林木树高、冠幅、以及胸径×树高×冠幅的增加,叶片性状表型可塑性降低,第1等级和第2等级组之间具有显著性变化(图2)。且整体上阴坡上林木叶片性状表型可塑性CV值大于阳坡。

图2 不同坡向上个体大小等级下叶片性状表型可塑性
2.4 不同坡向林木个体大小与竞争强度的关系
采用混合线性模型分析林木个体大小和竞争强度的关系,结果显示在研究样地中阴坡上林木种间竞争强度随着林木胸径、冠幅以及胸径×树高×冠幅的增加显著性增加(表5)。阳坡上林木种间竞争强度随着林木树高、冠幅以及胸径×树高×冠幅的增加而显著增加,随着胸径的增加而显著性减少(表5)。在阴坡上中内竞争强度随着胸径、树高的增加而显著性减少。但是在阳坡上林木种内竞争强度随着树高、冠幅以及胸径×树高×冠幅的增加而显著性增加(表5),随着胸径的增加而显著性减少。阴坡上林木中内竞争强度随着冠幅的增加而减少,但是模型拟合不具有显著性,随着胸径×树高×冠幅无显著性变化(表5)。
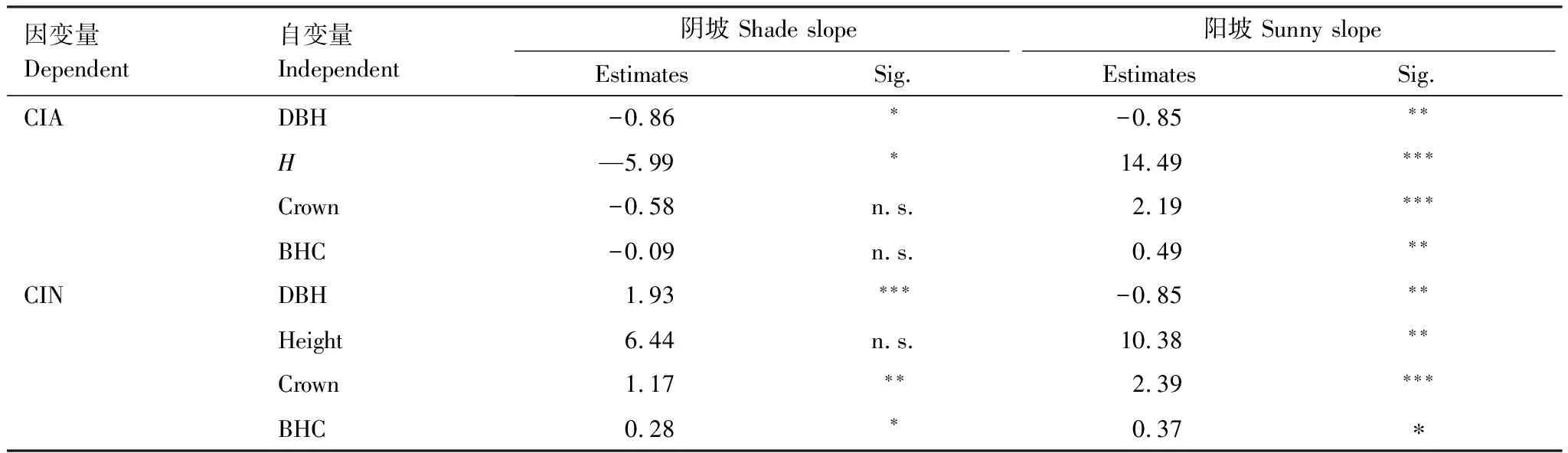
表5 不同坡向上林木大小与竞争强度的关系
3 讨论
叶片性状可塑性变化体现了植物对生长环境干扰的适应和响应,环境因素和竞争主要通过影响植物可利用资源条件影响植物表型可塑性[29—30]。研究中阴坡和阳坡样地是位于同一海拔梯度的两片天然次生林,坡向的改变导致环境因素包括太阳辐射强度、土壤理化性质、气温等都随之改变。在样地群落环境因子调查以及土壤元素测定结果显示,阴坡上其大气湿度和土壤湿度都高于阳坡,土壤养分条件优于阳坡,整体上阴坡群落环境资源条件优于阳坡(表1,表2)。研究结果显示,在川西亚高山天然次生林群落中,样地中阴坡上群落叶片比叶面积、碳含量、氮含量、磷含量、钾含量、碳磷比、碳氮比、氮磷比其表型可塑性大于阳坡(图1)。叶片比叶面积、碳、氮、磷、钾含量反映了林木其资源获取和利用能力[31—32],碳氮比、碳磷比、氮磷比反映了林木其碳氮同化能力和养分吸收利用速率[31]。本研究推测,在资源环境条件更好的阴坡上,植物更容易通过调节与养分利用与吸收相关的叶片表型来适应群落中的变化环境,提高植物体在多变环境下的生存能力。同时,这种高表型可塑性反映了植物在干扰环境下的高生存能力,该植物体往往具有高生态幅[33]。

表2 不同坡向的土壤元素参数值(平均值±标准偏差)
进一步比较阴坡和阳坡上平均林木竞争强度发现阴坡其种内竞争强度、种间竞争强度都高于阳坡(表4)。与阳坡相比,在群落环境资源条件更优的阴坡上,林木间的高竞争强度很可能是叶性状高可塑性的主要原因。在阴坡上,当林木间竞争强度逐渐增加时,林木能吸收和利用资源渐少,林木主要通过增加光照以及养分吸收和利用速率提高其生长和适应能力来响应竞争环境变化,而且植物生长受到资源环境压力较少,林木很容易调节植物叶片性状,此时群落内叶性状可塑性较高。
胸径、树高、冠幅、胸径×树高×冠幅都反映了林木个体大小。研究发现样地中阴坡和阳坡上林木个体大小与表型可塑性关系呈现相反的模式,结果显示阴坡上随着个体大小的增加,表型可塑性增加,阳坡上随着个体大小的增加,表型可塑性降低(图2)。这可能也是不同坡向上环境条件和竞争不同程度影响林木叶片表型可塑性的证据之一。研究样地中的阴坡水分和养分资源条件优于阳坡,林木生长环境主要受到竞争作用影响。阴坡上随着林木个体的增加,表型可塑性增加,说明林木在整个生长过程中都能够适应竞争强度变化。当林木间竞争强度较大时,林木可吸收和利用资源有限,随着个体的增加,植物受到竞争干扰增加,波动较大,叶性状表型可塑性增加。反之,研究样地中的阳坡上林木生长主要受到环境资源条件限制,植物在限制环境条件表现出性状相似性,表型可塑性较小。随着个体大小增加,需要获取和利用资源增加,进一步增加资源限制,植物生存压力加剧,因此植物对竞争环境变化适应性降低,叶性状表型可塑性进一步降低。
有趣的是,分析种内竞争和种间竞争对林木个体生长的关系发现,阴坡上种内竞争强度随着林木胸径、树高增加而显著性减少,但在阳坡上随着树高、冠幅以及胸径×树高×冠幅的增加而显著性增加(表5)。本研究推测,不同坡向之间的资源环境差异主要影响了林木间种内竞争作用与生长的关系。因此林木的可吸收和利用资源变化可能主要由林木间种内竞争强度增加引起。因为种内竞争主要由相同物种的资源利用普遍重叠而造成。研究样地中阴坡环境资源条件优于阳坡,随着林木不断生长,胸径、树高的增加,资源利用重叠增加,植物间种内竞争愈激烈。在激烈的竞争环境条件条件下,种内竞争引起的自疏效应增加,种群密度逐渐降低,种内竞争也随之降低。但是由于阳坡其生长条件较为恶劣,林木生长主要受到环境作用限制,种内竞争引起的自疏效应不明显,因此种内竞争强度反而随着个体大小增加而增加。
综上,研究发现样地中阴坡和阳坡上木本植物叶片性状表型可塑性表达具有差异,不同坡向上的环境资源条件影响了林木间竞争作用对叶片表型可塑性的影响作用大小。在阴坡上,其群落资源环境条件优于阳坡,林木生长受到环境资源限制较小,在林木生长过程中,竞争引起的资源重叠加剧,尤其是种内竞争强度的变化,此时植物通过调整叶片性状增加其适应能力,阴坡上林木叶片可塑性较高。但是干旱而且养分资源相对贫瘠的阳坡,林木生长受到环境资源条件限制,在林木生长过程中,竞争进一步引起资源重叠加剧。但是受到生长限制的林木很难通过调整叶片性状来增加其适应能力,叶片表现出与环境逆境适应相关的性状,此时叶片表型可塑性较低,最后阳坡上叶性状表型可塑性低于阴坡。因此,在阴坡上叶片表型可塑性主要受到竞争强度的影响,而在阳坡上主要受环境资源的限制。因此本研究认为群落环境的资源条件会限制竞争对叶片性状的影响作用大小。当林木生长受到环境资源限制时,竞争对叶片性状变化的影响作用会降低,此时对叶片性状变化的影响上环境因素则起到主导作用。需要指出的是,由于受到川西地区高山地形险峻的影响,研究样地数目的限制导致本文无法继续探讨坡向的转变如何具体影响了林木资源环境条件,以及进一步探究环境因素如何具体影响了竞争作用对叶片表型可塑性影响作用的大小。因此,未来研究可能需要增加研究样地,建立合适坡向梯度变化,或者建立长期样地观测,才能进一步阐明环境因素是如何影响竞争作用对叶片表型可塑性影响作用大小。
猜你喜欢
昆明医科大学学报(2022年1期)2022-02-28
种子科技(2021年17期)2021-10-29
思维与智慧·下半月(2021年7期)2021-08-13
昆明医科大学学报(2021年8期)2021-08-13
昆明医科大学学报(2021年6期)2021-07-31
中成药(2018年11期)2018-11-24
现代农村科技(2018年11期)2018-11-15
课程教育研究(2017年11期)2017-04-17
