
牛肉源假单胞菌的分离鉴定及菌株致腐能力差异比较
2022-03-28 12:08张若煜罗欣朱立贤刘昀阁陈雪郝剑刚黄鑫张一敏
现代食品科技 2022年3期
张若煜,罗欣,朱立贤,刘昀阁,陈雪,郝剑刚,黄鑫,张一敏*
(1.山东农业大学食品科学与工程学院,山东泰安 271018)(2.国家肉牛牦牛产业技术体系乌拉盖站,内蒙古乌拉盖管区 026321)(3.国家肉牛牦牛产业技术体系延边站,吉林延边 133000)
牛肉因其较高的营养价值,广受消费者的青睐。牛肉的腐败主要是由于微生物的增长及其胞外酶分解代谢引起的。其中假单胞菌属为有氧包装条件下的冷鲜牛肉中的优势腐败菌[1,2],该菌属多具有较强的产胞外蛋白酶与脂肪酶能力,能够分解利用肉品中的蛋白质、脂质等为自身的生命活动提供能量来源[3]。在此过程中,假单胞菌通过分泌胞外蛋白酶分解肌原纤维及肌浆蛋白[4],并通过脱氨作用分解氨基酸并产生二甲基硫等具有腐臭气味的硫化物等[5],同时分泌脂肪酶催化脂肪等分解为脂肪酸、甘油等[6],产生多种代谢产物,使牛肉变粘、变色,产生异味。
假单胞菌株对肉品的腐败能力较强,其蛋白酶与脂肪酶活性较高。顾春涛等[7]发现冷鲜牛肉中致腐能力较强的假单胞菌株具有较强的胞外蛋白酶活性,主要作用于肌原纤维;Wang等[8]研究发现在鸡肉中致腐能力较强的假单胞菌也具有较强的蛋白酶活性,使肌球蛋白、肌动蛋白、肌钙蛋白发生了不同程度的降解;葛阳杨等[9]发现大黄鱼中腐败能力较强的菌株除具有较强的蛋白酶活性外,也具有较强的脂肪酶活性。目前具有较强致腐能力的菌株其胞外酶活性较强已得到证实,但具有较强酶活性的菌株是否对肉品具有较强致腐能力尚未得到明确的定论,因此为了明晰假单胞菌的产酶能力对牛肉的腐败作用,本试验筛选了产酶能力存在差异(3株强蛋白酶活性、3株强脂肪酶活性、1株弱蛋白酶及弱脂肪酶活性)的7株假单胞菌,研究 7株菌回归污染后牛肉样品的假单胞菌数量、TVB-N值及感官品质的变化,旨在确定假单胞菌的产酶能力与牛肉腐败的关系,为防控牛肉的腐败提供理论支持。
1 材料与方法
1.1 材料与试剂
经检疫合格的新鲜牛背最长肌,山东某肉牛屠宰企业;碱性蛋白酶活性测定试剂盒、脂肪酶活性测定试剂盒,索桥生物公司;假单胞菌基础培养基、假单胞菌CFC添加剂,北京陆桥技术股份有限公司;细菌基因组DNA提取试剂盒,BioFlux公司;PCR全套试剂,Takara公司;生理生化鉴定全套试剂,青岛高科技工业园海博生物技术有限公司;所用引物由上海生工生物工程股份有限公司合成,辐照技术由蓝孚医疗科技(山东)有限公司完成。
1.2 仪器与设备
1300 SERIES A2生物安全柜,美国 Thermo Scientific公司;DHP系列智能型生化培养箱,上海一恒公司;CFX 96、PCR仪PTC-200实时PCR检测系统,美国Bio-Rad公司;SpectraMax M5微孔板检测系统,美国Molecular Devices公司;GeneQuant 100微量核酸蛋白测定仪,英国Biochrom公司;G15DWS蒸汽压力灭菌锅,厦门致微公司;5804R离心机,德国Eppendorf公司。
1.3 方法
1.3.1 牛肉源假单胞菌的分离与纯化
将宰后48 h的牛背最长肌分割成9块3 cm厚的牛排,随机分配到托盘包装中,TQBC-0775包装托盘盒[23 ℃、0%相对湿度下,透氧率为10 cm3/(m224 h),38 ℃、90%相对湿度下,水蒸气透过率为15 g/(m224 h)];Lid 1050/550密封阻隔膜[23 ℃、0%相对湿度下,透氧率为25 cm3/(m224 h),4 ℃、100%相对湿度下,水蒸气透过率:10 g/(m224 h)],参考Yang等[10]的方法对4 ℃有氧贮藏0、3、7、14和21 d的牛肉进行假单胞菌计数,并随机挑取最高稀释度可计数平板的10%的菌株划线纯化两次,纯化后的单菌落于-80 ℃保存。
1.3.2 假单胞菌产蛋白酶能力的研究
将分离纯化的假单胞菌株点样于脱脂乳平板(脱脂乳粉5%、琼脂2%)上,28 ℃培养48 h后,测量菌落直径与透明圈直径[11],通过比较H/C(透明圈直径与菌落直径之比)来初步测定菌株产蛋白酶能力的强弱。筛选H/C较大的菌株参照碱性蛋白酶活性测定试剂盒的方法对菌株进行碱性蛋白酶活性的测定[12]。
1.3.3 假单胞菌产脂肪酶能力的研究
将分离纯化的假单胞菌株点样于 Tween-80琼脂平板(蛋白胨1%、酵母膏0.5%、NaCl 0.5%、CaCl20.01%、Tween-80 0.1%、琼脂2%),28 ℃培养48 h后,测量D/C(沉淀圈与菌落直径之比)[13],通过D/C值来初步测定菌株产脂肪酶能力的强弱。筛选沉淀圈与菌落直径的比值较大的菌株参照脂肪酶活性测定试剂盒的方法对菌株进行脂肪酶活性的测定[14]。
1.3.4 假单胞菌的生理生化鉴定
通过蛋白酶活性与脂肪酶活性的实验筛选出3株强蛋白酶菌株、3株强脂肪酶菌株、1株弱蛋白酶弱脂肪酶菌株作为对照进行后续实验。
对7株菌进行糖醇类发酵实验,氧化酶反应,淀粉水解,硝酸盐产气,明胶液化,42 ℃生长实验等生理生化实验[15,16]。
1.3.5 假单胞菌的分子生物学鉴定及系统发育树的构建
细菌基因组DNA的提取:参照BioFlux公司细菌基因组DNA提取试剂盒说明书提取。
16S rDNA的聚合酶链式反应(polymerase chain reaction,PCR)扩增:PCR扩增引物采用细菌通用引物 27F(5’-AGAGTTTGATCCTGGCTCAG-3’),1492R(5’-TACGGYTACCTTGTTAYGACTT-3’)进行扩增。反应条件为:94 ℃预变性 30 s,98 ℃变性 10 s,55 ℃退火30 s,72 ℃延伸1 min。将测定的16S rDNA序列登陆NCBI进行BLAST比对,使用MEGA 7软件绘制系统发育树。
1.3.6 7株菌的回归污染
将培养至稳定期的菌液稀释至105CFU/mL,将经过辐照灭菌制备的无菌牛肉(3 cm×3 cm×3 cm)在菌液中浸渍2 s,之后托盘包装于4 ℃贮藏,在贮藏的第0、2、4、6和8 d时测定假单胞菌数量、挥发性盐基氮,并进行感官评价。
1.3.6.1 假单胞菌数
假单胞菌数参照1.3.1假单胞菌数的计数方法。
1.3.6.2 TVB-N测定
挥发性盐基氮的测定方法参照GB 5009.228-2016方法中的自动凯氏定氮仪法[17]。
1.3.6.3 感官品质
参照Zhang等[18]的方法对冷藏期间的生鲜牛肉按照1-9分值进行感官评定,评定小组由10名经过培训的小组成员组成,从样品的色泽、气味、嫩度、黏性等方面综合评定其整体可接受性,9分为最好,1分为最差,低于6分时被判定为不可接受。
1.4 数据统计分析
采用Microsoft Excel 2007和SigmaPlot 12进行数据处理和作图,假单胞菌数、TVB-N值、感官品质利用SAS软件(Version 9.0)的混合模型交互作用进行数据分析,p<0.05表示有统计学意义。
2 结果与讨论
在有氧贮藏条件下的冷鲜牛肉中,于 0、3、7、14和21 d共筛出216株假单胞菌,用于后续实验。贮藏前期(0~3 d)分离的假单胞菌数较少,共计31株,贮藏中后期(7~21 d)分离假单胞菌数较多,共计185株。
2.1 产酶较高菌株的酶活性的测定
2.1.1 产蛋白酶较高菌株的蛋白酶活性的测定

表1 产蛋白酶能力较强菌株的H/CTable 1 H/C of the strain with strong protease production ability
通过脱脂乳平板实验,从分离纯化的菌株中初步筛选出了16株H/C较大的菌株,结果如表1所示,筛选的16株菌的 H/C均大于 3.00,其中 PP3413、PP754、PP2142、PP1438和PP7410的H/C大于3.2,分别为3.38、3.30、3.29、3.26和3.23,产蛋白酶能力高于其他产蛋白酶菌株。PP3413的H/C最大。16株产蛋白酶能力强的菌株大多分离于冷鲜牛肉贮藏后期,表明在贮藏中后期,假单胞菌能够利用肉品中的蛋白质等营养物质,产蛋白酶能力强的菌株逐渐占据优势地位。Casaburi等[19]认为葡萄糖是假单胞菌优先利用的能量物质,当牛肉中游离的葡萄糖、乳酸盐等物质不足以为假单胞菌的生长提供充足的能量时,假单胞菌开始分解蛋白质利用氨基酸等物质生成含硫化物、酯、酸等异味物质,产蛋白酶能力较强的假单胞菌在贮藏后期通过蛋白水解能力深入肉中利用新的营养物质[20]。
Andreani等[21]认为假单胞菌于低温环境下分泌的蛋白酶主要为AprX碱性金属蛋白酶家族,在多数荧光假单胞菌中,AprX碱性金属蛋白酶家族被认为是参与食品腐败的唯一蛋白酶家族。利用福林酚显色法测定了以上16株菌株粗酶液碱性蛋白酶的活性,按碱性蛋白酶活性从大到小的排序方式排序,结果如图 1所示。16株菌在24 h时的碱性蛋白酶活性均在8.00 U/mL~12.00 U/mL之间,PP3413、PP754和PP1438的碱性蛋白酶活性大于10 U/mL,分别为11.3 U/mL、10.06 U/mL和10.05 U/mL,其中PP3413的碱性蛋白酶活性最高。同样,顾春涛等[7]研究发现在贮藏15 d的冷鲜牛肉汁中,有3株优势腐败假单胞菌具有较高的蛋白酶活性,均高于10.62 U/mL。因此筛选产蛋白酶能力较强,碱性蛋白酶活性较大的PP3413、PP754、PP1438用于后续实验。
2.1.2 产脂肪酶能力较高菌株的脂肪酶活性测定
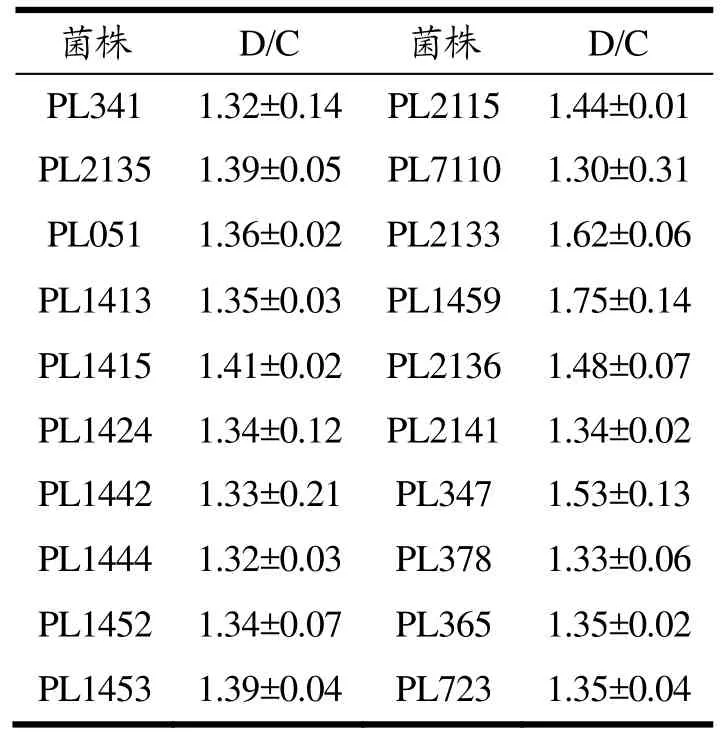
表2 产脂肪酶能力较强菌株的D/CTable 2 D/C of the strains with higher lipase production ability
通过Tween-80琼脂平板实验,从分离纯化的菌株中初步筛选出了20株产脂肪酶能力较强的菌株,结果如表2所示,20株菌的D/C值均在1.30以上,PL2115、PL2133、PL1459、PL1415、PL2136和PL347的D/C值均大于1.40,分别为1.44、1.62、1.75、1.41、1.48和1.53,其中PL1459的D/C值高于其他5株菌。20株产脂肪酶能力强的菌株也多分离于牛肉贮藏后期,表明随着肉品的腐败,菌株亦通过对脂肪的分解利用为自身生长代谢提供能量,产脂肪酶能力强的菌株比例呈上升趋势。
利用铜皂法测定各菌株粗酶液中脂肪酶的活性,按脂肪酶活性从大到小的排序方式排序,结果如图 2所示。20株菌在 24 h时的脂肪酶活性均在 0.6~1.2 U/mL之间,PL347、PL1415、PL1459和PL2133的脂肪酶活性大于1.1 U/mL,分别为1.19、1.15、1.14、1.10 U/mL,其中PL347的脂肪酶活性最高。而戴金岳等[22]测得冷鲜猪肉中的特定腐败假单胞菌(P.koreensis)的脂肪酶在3 d时活性最强,达到51.85 U/g,不同来源的假单胞菌其脂肪酶活性存在较大差异可能与假单胞菌的菌种及其利用的脂肪种类密切相关。假单胞菌的分泌的脂肪酶属于α/β型水解酶超家族[23],Ramani等[24]认为假单胞菌的脂肪酶对牛脂底物利用能力较强。因此筛选产脂肪酶能力较强,脂肪酶活性较大的PL347、PL1415、PL1459用于后续实验。
2.2 假单胞菌种水平的鉴定
通过蛋白酶与脂肪酶实验筛选出1株产蛋白酶及脂肪酶能力都较弱的菌株,命名为P011,将P011与产蛋白酶能力强的菌株PP3413、PP754、PP1438及产脂肪酶能力强的菌株 PL347、PL1415、PL1459用于后续实验。
2.2.1 生理生化鉴定结果
参考《伯杰细菌手册》和《常见细菌系统鉴定手册》,结合表3结果,初步鉴定分离菌株P011不液化明胶,42 ℃不生长,不能利用阿拉伯糖等与恶臭假单胞菌(P. putida)鉴定特征接近,分离菌株PP3413、PP754、PL1415能利用葡萄糖、果糖、蔗糖,不能利用麦芽糖为唯一碳源等与荧光假单胞菌(P.fluorescens)鉴定特征相接近,PP1438硝酸盐产气等与P. salomonii鉴定特征相近,PL347利用葡萄糖、蔗糖、阿拉伯糖醇等与草假单胞菌(P. poae)与其生理生化特征相似,PL1459不利用果糖、蔗糖等与P.libanensis生理生化特征相近。

表3 7株菌的生理生化鉴定结果Table 3 Physiological and biochemical identification results of 7 strains
2.2.2 系统发育树的构建
由图3可知,将筛选的7株菌以总DNA为模板PCR扩增后经电泳检测,均得到了1500 bp左右的条带,将PCR产物进行测序,测序结果登录NCBI网站,进行Blast序列比对,发现PP3413、PP754、PL1415与荧光假单胞菌的16S rDNA同源性达99%以上,结合生理生化试验的结果,可判定 PP3413、PP754、PL1415为荧光假单胞菌。此外,结合16S rDNA与生理生化结果将P011、PL347、PP1438、L1459分别判定为恶臭假单胞菌、草假单胞菌、P. salomonii、P.libanensis。
有学者研究表明在冷藏肉中,荧光假单胞菌占初始假单胞菌数量的 60%以上,腐败时占细菌总数的90%[25,26],恶臭假单胞菌是有氧冷藏牛肉腐败初期中常见的假单胞菌种之一[27],而草假单胞菌、P.salomonii、P. libanensis在牛肉中分离较少,草假单胞菌曾在羊奶中被检出[28],P. salomonii、P. libanensis多分离于土壤、植物、水源等。因此PL347、PP1438、PL1459的来源可能是牛饲养过程中的土壤、饲料以及水源等。
值得注意的是,肉品中常被检出的荧光假单胞菌与恶臭假单胞菌分离时期不同,恶臭假单胞菌多分离于贮藏初期[29]而荧光假单胞菌在腐败末期也常被检出[4,7]。且两种菌的蛋白酶与脂肪酶活性差异较大,本试验中,荧光假单胞菌PP754、PP3413具有较强的蛋白酶活性,荧光假单胞菌PL1415具有较强的脂肪酶活性,而恶臭假单胞菌 P011不具有蛋白酶活性及脂肪酶活性。Reichler等[30]测定不同假单胞菌种的蛋白酶及脂肪酶活性,结果表明荧光假单胞菌部分菌株具有蛋白酶活性与脂肪酶活性,而恶臭假单胞菌不具有蛋白酶活性,因此假单胞菌蛋白酶与脂肪酶的分泌可能与假单胞菌的菌种相关。恶臭假单胞菌在肉品贮藏后期检出较少,可能与其较弱的蛋白酶、脂肪酶活性,从而无法较好的利用肉品中的蛋白质、氨基酸脂肪等密切相关,荧光假单胞菌则因其较强的蛋白酶与脂肪酶活性使其成为肉品中的优势腐败菌。
2.3 假单胞菌的回归污染
2.3.1 假单胞菌计数
菌株种类和贮藏时间的交互作用显著影响牛肉中假单胞菌数(p<0.05)。随着贮藏时间的延长,各处理组中假单胞菌落数均呈现上升趋势,而对照组中未检出假单胞菌(检出限为10.00 CFU/g),表明回归接种前的牛肉样品未受到假单胞菌的污染。接种 P011、PL347的牛肉样品中假单胞菌落数从第四天时开始显著低于其他处理组(p<0.05),分别为5.60 log CFU/g、5.03 log CFU/g,在贮藏第八天时分别达到 6.59 log CFU/g和6.25 log CFU/g。而其他处理组中假单胞菌的生长较快,在贮藏第四天时就已超过6.80 log CFU/g,PP754、PP3413、PP1438、PL1415及PL1459分别为7.63、8.26、6.87、7.42、7.72 log CFU/g;贮藏至第八天时,分别达到了 9.65、9.33、8.29、8.97、9.15 log CFU/g,能较好的利用牛肉的营养物质来满足自身生长需求。其中接种PP754的样品在第八天时假单胞菌数最高,生长能力最强。值得注意的是,有学者发现在冷鲜鸡肉中具有较强蛋白酶活性的莓实假单胞菌其数量在第七天时已经达到了10.20 log CFU/g[4];在冷鲜牛肉中具有较强蛋白酶活性的假单胞菌其数量在第六天时已经超过了7.23 log CFU/g[7],本实验中发现产蛋白酶能力较强的两株菌PP754、PP3413处理组的假单胞菌数在第八天时高于其他假单胞菌处理组,因此蛋白酶活性强的假单胞菌在牛肉中的生长能力可能强于脂肪酶活性强的假单胞菌。
假单胞菌在冷鲜牛肉上的生长能力与其腐败能力存在一定的相关性,除PL347(第八天时,假单胞菌数量为6.25 log CFU/g)外,其他5株产酶能力较强的菌株在冷鲜牛肉中具有较强的生长能力(第八天时,假单胞菌数量均高于8.20 log CFU/g)。底物可能是影响假单胞菌脂肪酶分解能力的关键因素之一[31]。而产脂肪酶能力较强的PL347在牛肉中生长能力较弱,可能是由于 PL347分泌的脂肪酶对牛脂的分解能力较差。刘武[32]认为假单胞菌分泌的脂肪酶对不同底物的分解能力有较大差异,在p-NP辛酸酯与p-NP葵酸酯不同底物中的脂肪酶活性分别为100.00%与71.30%。Borch等[33]认为与细菌生长相关的营养物质利用情况会影响食品的腐败速度,这5株产酶能力较强的菌株在牛肉中具有较强的生长能力可能是由于能较好地利用牛肉中的营养物质。因此,产酶能力较强的菌株在牛肉中的生长能力不一定较强,能否较好的利用相应类别的底物与其生长繁殖密切相关。
2.3.2 TVB-N值
TVB-N是评价肉质新鲜度的重要指标之一。菌株和贮藏时间的交互作用显著影响TVB-N值(p<0.05)。所有样品的初始TVB-N值在10.64 mg/100 g左右,随着贮藏时间的延长,均呈现上升的趋势,其中接种PP754、PP3413、PL1459、PL1415的牛肉样品其TVB-N值增长速率最快,贮藏第八天时显著高于其他处理组(p<0.05,图6),分别为25.02、22.06、21.80、20.35 mg/100 g。在第六天时,接种PP754、PP3413、PL1415、PL1459的牛肉样品就已超过GB 2707-2016限量标准(15 mg/100 g),分别为 20.63、19.04、15.28、15.23 mg/100 g。其中接种PP754牛肉样品的TVB-N值从第四天时显著高于其他处理组(p<0.05),为14.99 mg/100 g;在腐败末期(第八天)达到了25.02 mg/100 g。对照组在整个贮藏期间TVB-N值增长较小(9.67~12.68 mg/100 g)。
本研究结果表明,接种产酶能力较强的菌株如PP754、PP3413(在体外蛋白酶活性较强)和PL1459、PL1415(体外脂肪酶活性较强)使牛肉中的 TVB-N值显著升高,该结果与上述假单胞菌数结果相一致。黄林等[34]也发现在冷鲜猪肉中,TVB-N值与假单胞菌数量也存在正相关性[35],当假单胞菌达到 8.30 log CFU/g时,其TVB-N值已超过25.00 mg/100 g。然而,具有较强脂肪酶活性的PL347使接种后牛肉在贮藏第八天时的TVB-N值为13.95 mg/100 g,显著低于其他产酶活性较强的假单胞菌(p<0.05)。同样,PP1438(具有强蛋白酶活性)处理组在贮藏第八天时的TVB-N也处于较低水平为15.82 mg/100 g。与该结果相类似,Wang等[8]研究发现在冷鲜鸡肉中具有较强蛋白酶活性、可降解肌钙蛋白与肌动蛋白的杀鲑气单胞菌因在肉中生长能力较低而样品中对应的 TVB-N值也显著低于其他菌株(第七天时达到30 mg/100 g),而不同的是本实验分离菌株的TVB-N值与Wang等[8]实验分离菌株的TVB-N值差异较大,可能是与菌种、肉的种类相关。进而说明,并不是所有产酶能力较强的菌株在肉品中的腐败能力均比较强,这与其菌种及在肉品中的生长能力也有一定的关系。同时本研究发现,产蛋白酶能力较强的菌株 PP754、PP3413其TVB-N值分别高于产脂肪酶能力较强的菌株PL1459、PL1415,这意味着产蛋白酶活性较强的假单胞菌其腐败能力可能优于产脂肪酶较强的菌株。
2.3.3 感官品质
接种7株腐败假单胞菌的牛肉样品的感官品质变化如图7所示。菌株和贮藏时间的交互作用显著影响感官品质评分(p<0.05),所有处理组的牛肉样品随贮藏时间的延长,感官评分均逐渐降低。贮藏第四天时,PP754、PP3413、PP1438、PL347、PL1415和PL1459处理组的感官评分就已低于6.00,分别为4.33、4.92、5.71、5.64、5.53和4.42,被判定为不可接受。在第六天时,接种PP754的牛肉样品的感官评分为2.41,显著低于其它处理组(p<0.05),其在贮藏期间腐败速率最快,在贮藏末期(第八天)肉色发绿,产生强烈的腐臭味,感官评分为1.20,被判定为完全不可接受。
肉品感官品质的下降主要是由于肉品中蛋白质、脂肪等物质的降解造成的[36,37]。在本研究中,接种产酶能力较强菌株的牛肉样品,其感官评分显著低于接种产酶能力较弱菌株 P011的牛肉样品及对照组(p<0.05),Stanborough等[38]研究发现,假单胞菌能够分泌与肉品腐败相关的蛋白酶及脂肪酶,当假单胞菌达到较高种群数量时,分泌的胞外酶活性会导致肉类品质下降并产生不良气味。Ahm等[39]也研究发现在假单胞菌占据主导地位的低温有有氧贮藏的牛肉中,含有3-甲基丁酮-1-醇、乙酸乙酯、丙酮、2-丁酮和二乙酰等代谢产物,造成了肉品的感官品质下降,同时感官品质与 TVB-N值存在一定的相关性;Benjamin等[40]研究发现,在真空包装的冷鲜牛肉中,TVB-N值主要与a*值、b*值呈负相关性(r=0.43、r=-0.34)。本研究结果与之相似,TVB-N值上升较快的假单胞菌株其气味更加难以接受,并具有较差的肉色。因此具有较强的产酶能力且能利用牛肉中的营养物质的假单胞菌株致腐能力更强,感官品质更差。同时,感官品质被认为是评价肉品品质最直接的方式[41],在第8 d时接种蛋白酶活性较强的PP754(1.20)、PP3413(3.22)牛肉的感官品质分别低于接种脂肪酶活性较强的PL1459(2.59)、PL1415(3.41)牛肉的感官品质,这表明产蛋白酶能力较强的菌株与产脂肪酶能力较强的菌株相比可能会带来更难以接受的品质劣变。
3 结论
荧光假单胞菌是冷鲜牛肉中的优势腐败菌,在筛选的7株假单胞菌中,3株荧光假单胞菌的产酶能力较强,同时其腐败能力也处于较高水平,感官评价较差。假单胞菌的产酶能力与其致腐能力并不存在必然联系,部分产酶能力较强的菌株可能因其无法以牛肉为底物进行生长代谢或在牛肉中生长能力较弱而导致在牛肉中的致腐能力较差。此外,本研究发现产蛋白酶活性较强的菌株可能比产脂肪酶能力较强菌株的腐败能力更强。因此,具有较强的酶活性且能利用牛肉中的营养物质大量生长繁殖,并产生代谢产物的假单胞菌具有更强的致腐能力。在后续研究中可进一步探究假单胞菌酶活影响牛肉品质劣变的具体机制,探究降低假单胞菌的酶活性、减弱假单胞菌对牛肉的利用能力以及控制假单胞菌数量的方法,以遏制假单胞菌对冷鲜牛肉的腐败能力,从而有效延长牛肉的货架期。
猜你喜欢
中华胰腺病杂志(2022年4期)2022-08-23
江西水产科技(2022年2期)2022-05-17
当代水产(2022年3期)2022-04-26
中国饲料(2022年5期)2022-04-26
感染、炎症、修复(2021年1期)2021-07-28
食品安全导刊·中旬刊(2020年2期)2020-06-01
肉类研究(2019年8期)2019-09-10
求知导刊(2019年15期)2019-08-30
华东师范大学学报(自然科学版)(2019年3期)2019-06-24
文苑(2018年22期)2018-11-19
