
山东半岛不同种群鳗草种子形态学特征及萌发特性研究❋
2022-03-26 02:10谢坤秀牛淑娜迟会凯张沛东
中国海洋大学学报(自然科学版) 2022年4期
谢坤秀,牛淑娜,迟会凯,张沛东
(海水养殖教育部重点实验室(中国海洋大学),山东 青岛 266003)
海草床作为三大典型近海海洋生态系统之一,具有极高的生态服务价值,不仅能为众多的海洋动物提供食物来源与产卵、育幼、栖息、庇佑场所,还能起到减缓水流、净化水体、稳定沉积物、固碳等作用[1-7]。然而,海草床属脆弱性生态系统,受人类活动及自然因素影响,当前世界范围内的海草床不断退化,消失速率超过每年7%[8]。由于海草种子具有体积小、便于运输、能保持遗传多样性等优势,利用种子种植法进行海草床规模化修复逐渐成为近年来研究的热点。
种子种植法的成功主要取决于种子萌发与实生苗建成的效果,而种子的萌发受到不同种群的显著影响。van Lent等[9]发现,荷兰西南部和法国西部不同种群的鳗草(Zosteramarina)种子萌发率相差约15%。Koch等[10]发现,美国南部佛罗里达海域的川蔓草(Ruppiamaritima)种子萌发率显著低于东南部的北卡罗来纳州种群。影响种子萌发及实生苗活力的因素通常分为外部因素和内部因素,其中外部因素主要是种子萌发的环境条件,内部因素则主要包括种子规格和营养贮藏物质的含量[11-13]。目前,对于陆生植物种群间种子形态及营养物质含量对种子萌发及幼苗生长的研究已有许多报道[14-16],但关于海草的相关研究还较少。
鳗草是中国温带海域海草的优势种类,主要分布于山东、河北、辽宁沿海[17]。调查显示,山东半岛日照海域和乳山海域的鳗草海草床已经消失,而莱州湾芙蓉岛附近海域的鳗草海草床面积则从1 300 hm2退化至不足5 hm2[17]。当前,山东半岛主要以威海近海和青岛近海分布有面积较大的鳗草海草床。然而不同种群的鳗草种子规格、萌发特性和实生苗生长状况等存在的差异尚不明确。因此,探明不同种群鳗草种子的形态学特征和萌发特性,是实现优质种源筛选以及成功开展海草种子种植的关键。
本研究以鳗草为对象,分别采集山东威海天鹅湖海域、俚岛湾海域和青岛汇泉湾海域的鳗草种子,比较分析不同种群间鳗草种子的形态学及内部营养物质含量的异同;通过室内萌发实验,探讨不同种群间鳗草种子萌发率、实生苗存活率和生长的差异,从而查明不同种群鳗草种子的萌发策略,为优化鳗草种子种植技术提供理论参考。
1 材料与方法
1.1 种子采集及保存
2019年7月,分别于山东省威海市天鹅湖海域、俚岛湾海域和山东省青岛市汇泉湾海域采集鳗草种子(见图1)。首先选取鳗草分布较多的区域,通过人工方式收集生殖枝,收集过程中未破坏鳗草植株的营养枝及其地下组织,且未在同一区域进行大量采集。摘取具有种子的佛焰苞放入33 cm× 33 cm(长×宽)、网目大小为 1.1 mm的尼龙网兜中,将其置于自然海域并固定。待尼龙网兜中的佛焰苞充分降解后,将其取出倒入塑料桶中反复搅拌,去除漂浮的有机碎屑及小型贝类、虫类外壳等杂质,筛选出成熟的鳗草种子。筛选后的鳗草种子收集至广口瓶带回实验室,置于温度4 ℃、盐度30的海水中储存,期间每2天换水1次,每天定期搅拌以减少水中的真菌数量。
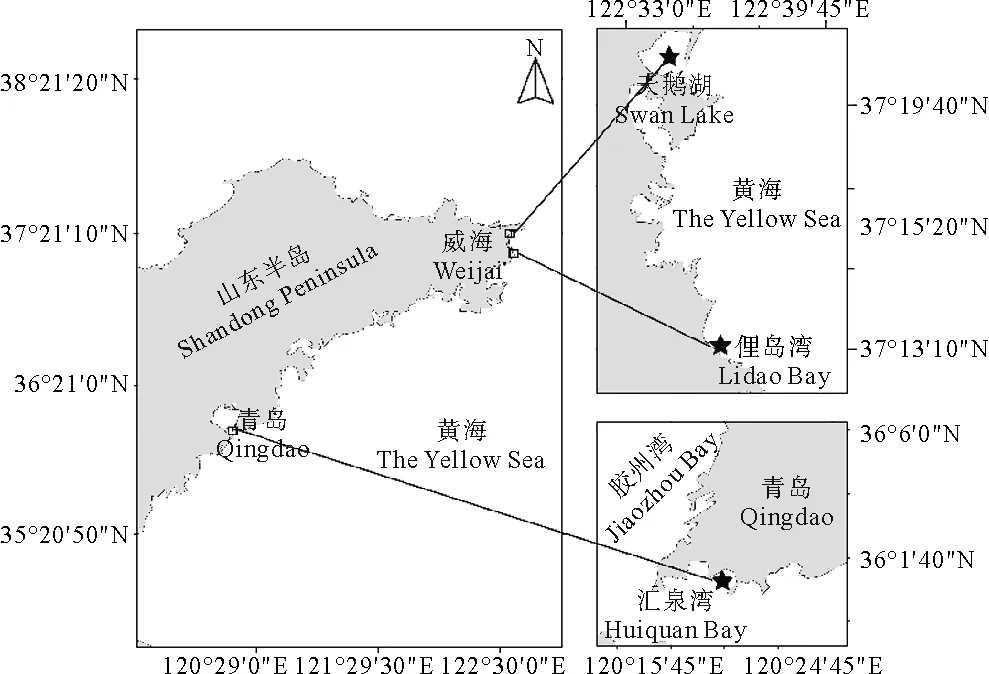
(星号代表采集地点。Asterisk indicates collection site.)
1.2 实验过程与样品分析
1.2.1 不同种群鳗草种子的形态学比较 从不同种群的鳗草种子中随机选取成熟饱满种子各100粒,吸干表面水分后分别测定种子的质量、长径、短径、纵肋数和最大横截面积等形态学指标。根据种子的长径和短径,计算种形系数(长径/短径)[18]。
1.2.2 不同种群鳗草种子的内部贮存物质测定 从不同种群的鳗草种子中随机选取成熟饱满种子各1 000粒,随机分为10份,分别对每份种子测定水分、总蛋白、总脂肪和总糖等内部营养贮存物质的含量,其中水分使用烘干法[19],总蛋白含量使用凯氏定氮法[20],总脂肪含量使用酸水解法[21],总糖含量使用3,5-二硝基水杨酸比色法[22]。
1.2.3 不同种群鳗草种子萌发的比较 于2019年11月,从不同种群的鳗草种子中随机选取成熟饱满的种子各300粒,先使用75%乙醇溶液浸泡0.5 min,无菌海水清洗3遍,再使用0.1%氯化汞溶液浸泡15 min,无菌海水清洗3遍。消毒后将各种群的种子平均分为3份,每份100粒,分别放入100 mL烧杯,烧杯中加入80 mL等量的盐度10的海水,置于光照培养箱(GXZ-0288,宁波江南仪器厂)中进行种子的快速萌发。萌发过程无光照,温度15 ℃。萌发实验共进行30 d,期间每天观察种子萌发情况并统计萌发数,将已萌发的种子挑出进行下一步的实生苗培养实验,每2天换水1次。实验结束后,计算各种群种子的累积萌发率、发芽指数(GI)和平均萌发历期(MTG)。
萌发率=萌发的种子数/种子总数×100%;
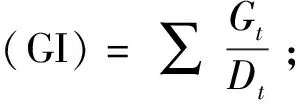
式中:Dt为发芽日数;Gt为与Dt对应天数的萌发粒数;n为相应天数萌发的种子数;d为计算种子萌发数量时的萌发天数;N为萌发种子的总数。
1.2.4 不同种群鳗草实生苗存活、生长及光合生理的比较 经种子快速萌发,天鹅湖、俚岛湾和汇泉湾种群分别获得124、113和81株实生苗。将不同种群获得的实生苗以盐度每天升高5的速率进行盐度驯化,至盐度30停止。各种群驯化后的实生苗以20株为1组种植于底部铺设5 cm厚细沙的100 mL烧杯中,株数取整后各种群分别使用4~6个烧杯(4~6个重复),并用防水纸标识组别,然后将烧杯放入40 cm × 40 cm × 20 cm(长×宽×高)的玻璃水槽中(实际水体25 L),置于上述的光照培养箱中培养30 d。期间保持水温17 ℃、盐度30、光照强度45 μmol·m-2·s-1、光照周期12 L∶12 D、pH=8.3±0.2,使用充氧泵持续充氧保持DO≥6.0 mg·L-1,每2天使用虹吸法换水1次以保持水体清澈,烧杯在玻璃水槽中的位置每天随机调换以避免光照条件差异的影响。经30 d培养结束后统计各烧杯实生苗的存活数,计算各种群的实生苗存活率;测定实生苗的叶长、叶宽、叶鞘长、茎长及根长等生长指标;随后在每个种群的实生苗中随机选取5株用于实生苗呼吸速率及光合速率的测定(黑白瓶法),随机选取5株用于实生苗光合色素含量的测定(浸提法[23])。
1.3 数据分析
对不同种群鳗草种子和幼苗的所有指标进行单因素方差分析,若存在显著差异再使用Duncan’s分析进行多重比较,结果用平均值±标准误表示(Mean±SE),显著性水平设置为p=0.05。对不同种群鳗草种子的形态学、生理学及萌发指标进行聚类分析及nMDS分析;对各种群鳗草种子的萌发率、幼苗存活率与种子形态学、生理学指标进行主成分分析(Principal component analysis,PCA),并对关键指标进行线性拟合。数据统计分析采用SPSS 25.0软件进行,分析结果使用Origin 2018 C、R语言进行绘图。
2 结果
2.1 不同种群鳗草种子的形态学特征
三个种群鳗草种子的形态学特征存在明显差异。其中汇泉湾种群的种子规格最大,天鹅湖、俚岛湾种群的种子规格较小,如表1所示。单因素方差分析显示,汇泉湾鳗草种子的质量显著高于天鹅湖、俚岛湾鳗草种子的质量(p<0.05),是2个种群种子的1.8倍。同时,汇泉湾鳗草种子的长径、短径及最大横截面积也显著高于其他2个种群的鳗草种子(p<0.05),但3个种群种子的种形系数无显著差异(p>0.05)。

表1 不同种群鳗草种子的形态学特征
2.2 不同种群鳗草种子的贮存物质含量
不同种群鳗草种子的内部贮存物质含量也不同。其中天鹅湖种群种子的营养物质含量较高,汇泉湾种群种子较低,如图2所示。三个种群鳗草种子的水分含量均高于40%。各种群种子内部营养贮存物质中均以总脂肪含量最少(≤1%),总糖含量最高,且不同种群间鳗草种子的总糖含量具有显著差异(p<0.05),其中天鹅湖种群种子的总糖含量最高,平均达到62%,分别是俚岛湾和汇泉湾种群种子的1.3和1.1倍。

(不同字母表示不同种群间差异显著(p<0.05)。Different letters indicate significant differences among the different populations(p<0.05).)
2.3 不同种群鳗草种子的萌发率与实生苗存活率
不同种群鳗草种子的萌发率、发芽指数及实生苗存活率均存在明显差异,其中天鹅湖种群最高,如表2所示。天鹅湖种群鳗草的种子萌发率、发芽指数(GI)和实生苗存活率均显著高于汇泉湾种群(p<0.05),分别是其1.5、2.2和2.9倍。各种群鳗草种子的平均萌发历期(MTG)均在12 d左右,无显著差异(p>0.05)。
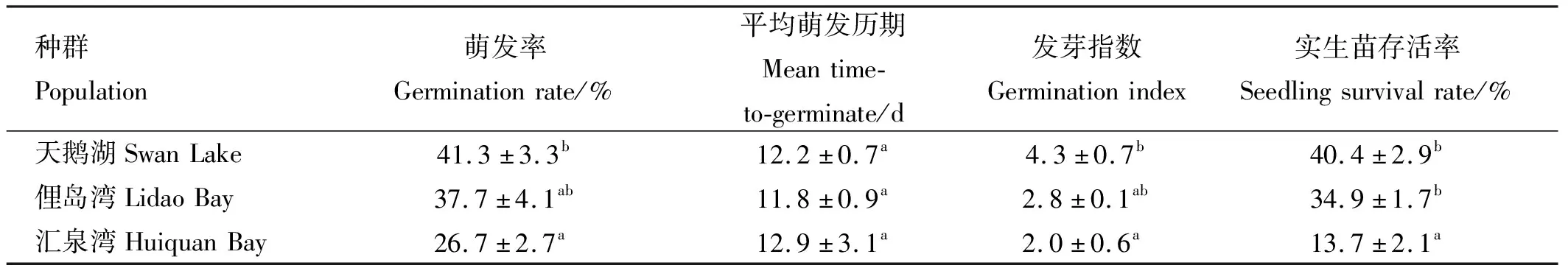
表2 不同种群鳗草种子萌发指标与实生苗存活率
2.4 不同种群鳗草种子的聚类分析
对山东半岛不同种群鳗草种子的形态特征和内部贮存物质等指标进行聚类分析,发现不同种群间存在明显区别,其中天鹅湖、俚岛湾种群的鳗草种子较为接近,聚为一类;汇泉湾种群鳗草种子与其它2个种群存在较大差异,单独聚为一类(见图3(A))。nMDS分析进一步验证了聚类分析的结果,应力值为0.039(见图3(B))。
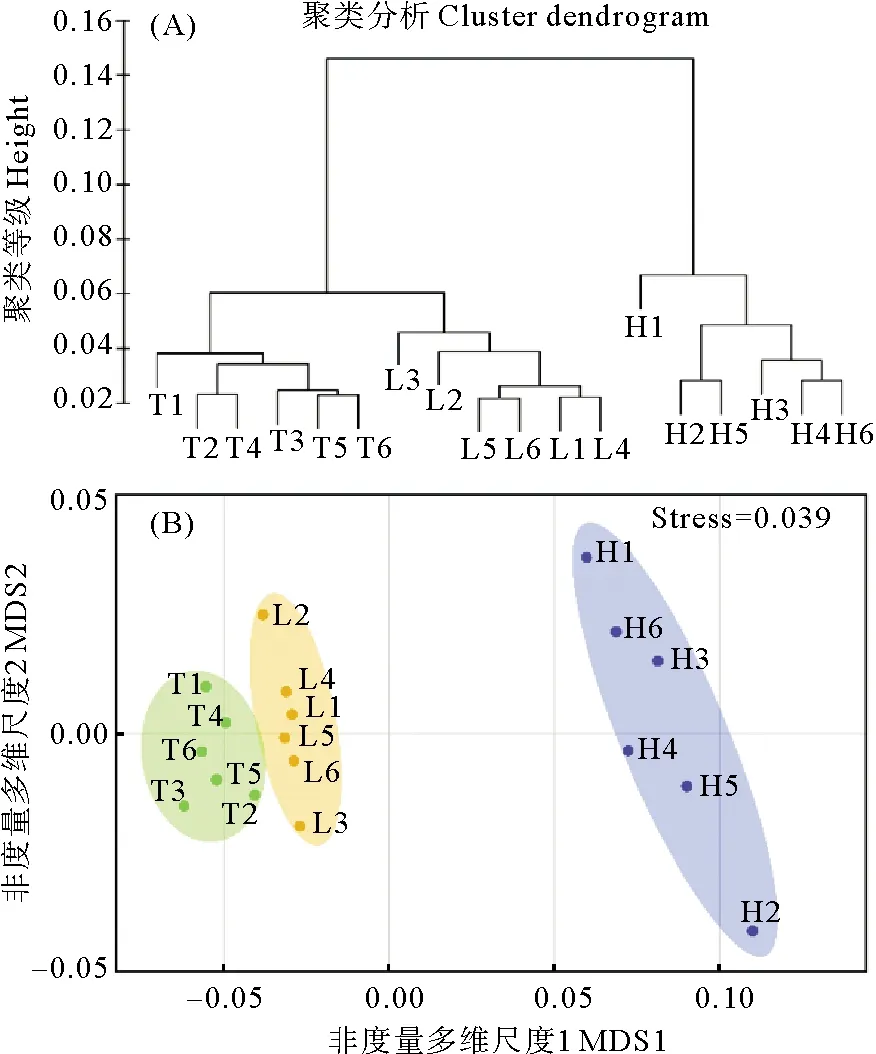
(图中T1~T6表示天鹅湖种群;L1~L6表示俚岛湾种群;H1~H6表示汇泉湾种群。T1~T6 in the figure represent the Swan Lake population; L1~L6 represent Lidao Bay population; H1~H6 represent Huiquan Bay population.)
2.5 不同种群鳗草种子萌发、实生苗存活与种子形态特征、内部贮存物质间的关系
主成分分析显示,种子萌发率、发芽指数、实生苗存活率与种子总糖含量、种子长短径、横截面积、种子质量、纵肋数、水分含量主要组成了第一排序轴,其中,以种子的萌发率、实生苗存活率、种子总糖含量和种子形态学这四个指标贡献较大。如图4所示,不同种群鳗草种子的萌发率、实生苗存活率与种子的总糖含量呈显著的正相关,而与种子水分含量、形态学指标呈显著的负相关。选取贡献较大的种子总糖含量、种子质量与种子萌发率、实生苗存活率进行曲线拟合显示,种子总糖含量与种子萌发率、实生苗存活率呈显著的线性正相关(p<0.05,见图5A),而种子质量与种子萌发率、实生苗存活率呈显著的线性负相关(p<0.05,见图 5B)。

(图中SL为天鹅湖种群,LB为俚岛湾种群,HB为汇泉湾种群;GR为萌发率,SSR为实生幼苗存活率,GI为发芽指数,TS为总糖,TF为总脂肪,MTG为平均萌发历期,TP为总蛋白,SW为质量,SD为短径,LD为长径,CSA为横截面积,LRN为纵肋数,WC为水分。SL in the figure is Swan Lake population, LB is Lidao Bay population, HB is Huiquan Bay population; GR is germination rate, SSR is seedling survival rate, GI is germination index, TS is total sugar, TF is total fat, MTG is mean time of germination, TP is total protein, SW is seed mass, SD is short diameter, LD is long diameter, CSA is cross-section area, LRN is number of longitudinal ribs, WC is water content.)
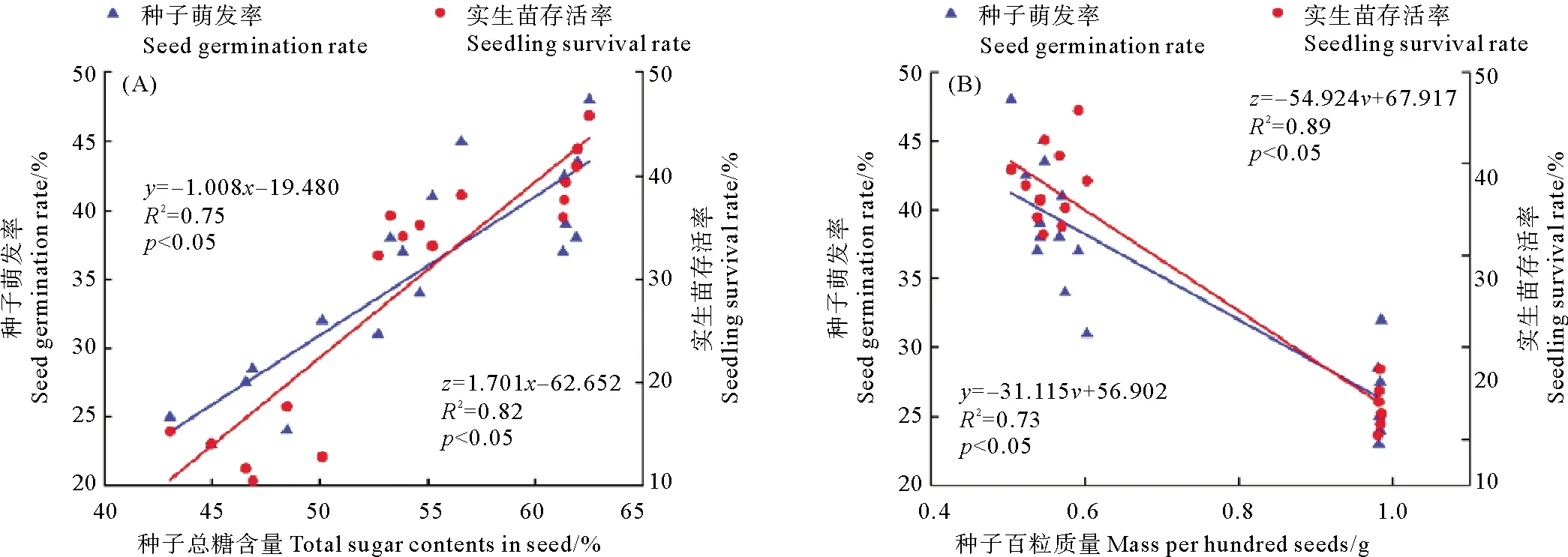
(图中y表示种子萌发率;z表示实生苗存活率;x表示种子总糖含量;v表示种子百粒质量。In the figure, y is seed germination rate; z is seedling survival rate; x is the total sugar content of seed; v is the mass per hundred seeds.)
2.6 不同种群鳗草实生苗的生长特性及光合生理
不同种群鳗草实生苗的生长存在差异。其中以天鹅湖种群的实生苗规格最大(见图6)。单因素方差分析显示,天鹅湖种群鳗草实生苗的叶宽、茎长和根长均显著大于俚岛湾鳗草实生苗(p<0.05),分别是其1.1、1.2和1.3倍;汇泉湾种群鳗草实生苗的茎长显著小于天鹅湖种群实生苗的茎长(p<0.05),仅为天鹅湖种群实生苗茎长的84%。
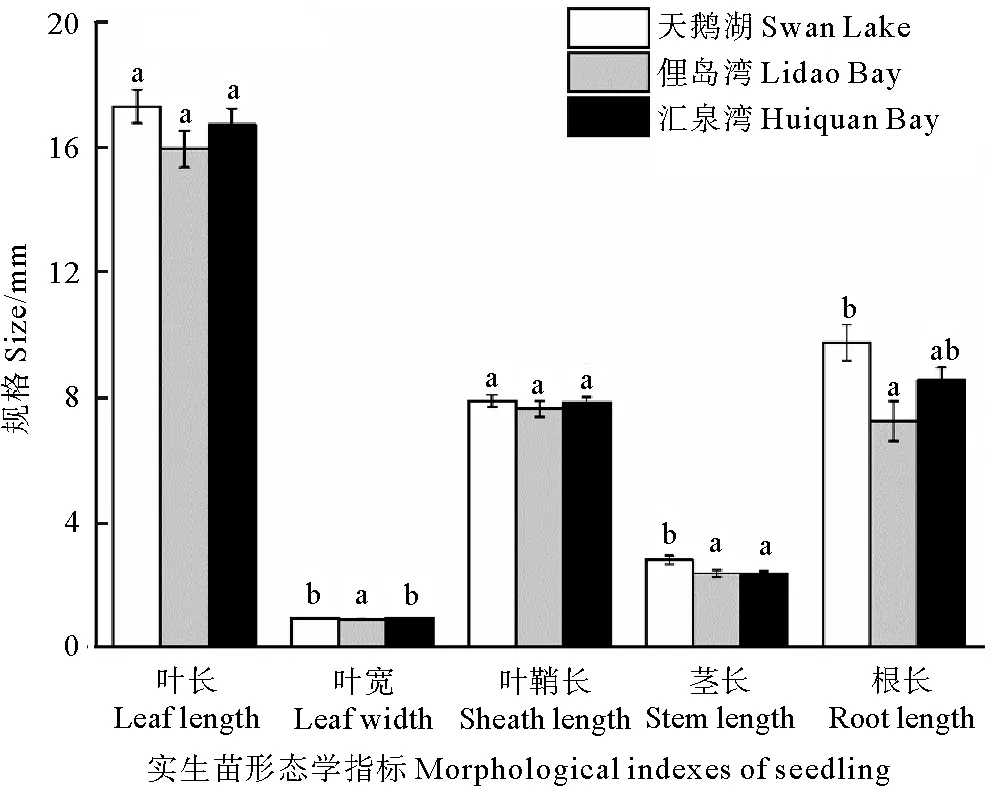
(样本数量为10~40。不同字母表示不同种群间差异显著(p<0.05)。The sample size is 10 to 40.Different letters indicate significant differences among the different populations(p<0.05).)
不同种群鳗草实生苗的光合色素含量和光合速率均存在差异。其中,天鹅湖种群实生苗的光合色素含量和光合速率最高,而俚岛湾种群实生苗最低(见图7和8)。单因素方差分析显示,天鹅湖种群鳗草实生苗的光合色素含量均显著高于俚岛湾种群和汇泉湾种群实生苗(p<0.05),其总叶绿素含量达到6.5 mg·g-1,是俚岛湾、汇泉湾实生苗叶绿素含量的1.5和1.2倍;天鹅湖鳗草实生苗的净光合速率和呼吸速率也显著高于俚岛湾种群实生苗(p<0.05),是其1.2倍。

(不同字母表示不同种群间差异显著(p<0.05)。Different letters indicate significant differences among the different populations(p<0.05).)
3 讨论
3.1 不同种群海草的种子形态学特征及其环境基础
同一植物种类的不同种群为更好地适应环境可能会发生遗传变异,并将稳定的性状表现在种子的品质和形态等方面,从而导致相同种类不同种群的种子形态学特征和内部贮存物质含量等存在显著差异[24]。本研究发现,汇泉湾鳗草种子的平均百粒质量达0.98 g,长径达3.69 mm,规格显著大于天鹅湖和俚岛湾种群的鳗草种子,与Xu等[25]研究结果相一致。Combs等[26]发现,采集于美国北卡罗莱纳州以北三个不同地点的鳗草种子大小显著不同,鲜质量在1.5~4.3 mg。对比其他研究发现,本研究中汇泉湾种子比丹麦利姆海峡的鳗草种子(长径3.4 mm)[27]和美国加利福尼亚湾北部的鳗草种子(长径3.0~3.5 mm)[28]更大,说明不同种群间鳗草种子的形态大小因受不同地区影响而存在较大差异。
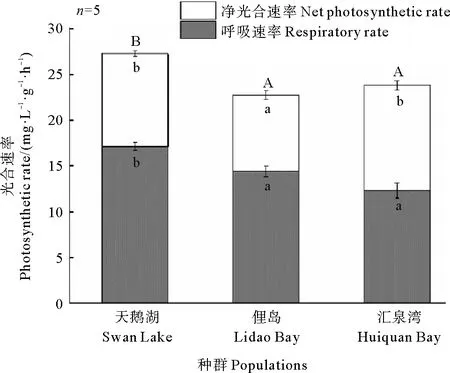
(不同字母表示不同种群间差异显著(p<0.05)。Different letters indicate significant differences among the different populations(p<0.05).)
个体间种子规格的变异一般是遗传型和环境因子共同作用的结果[29]。遗传型包括细胞核遗传、细胞质遗传和胚乳核遗传[30]。Smith等[31]发现,亚麻(Linumusitatissimum)植株母体的细胞质直接影响种子大小, 并且这种影响可能延续到子二代和子三代。然而李渊等[32]发现,爱尔兰、日本、韩国和中国青岛胶南、威海俚岛湾、威海天鹅湖等地的鳗草种群间不存在地理隔离形成的基因分化,因此鳗草种子种群间的形态学差异可能主要是由不同的环境条件所造成。
研究表明,不同的纬度、种子分布深度及水动力等环境条件对植物个体间种子规格变异有明显影响。张志翔等[33]发现,低纬度地区的小桐子(Jatrophacurcas)种子比高纬度地区的种子规格更大;Wyllie等[34]发现,美国部分海域鳗草种子的质量随纬度的降低而增大。本研究结果与这些研究相类似,三个种群鳗草种子的质量随天鹅湖、俚岛湾、汇泉湾纬度的降低而逐渐增大。种子分布深度也是影响种子规格的关键因素。彭鸿嘉等[35]发现,当种子处于较深底质时,较大的种子因储存更多的能量而形成更长的茎,从而顺利萌发并存活。汇泉湾底质为砂质[36],较天鹅湖的泥沙底质[22]和俚岛湾的砾沙底质[37]更松散,鳗草种子更易被深埋,因此,为适应环境并保证实生苗的顺利萌出,汇泉湾种子的个体更大。此外,Orth等[38]发现,海草植株在水流湍急的水域中会产生质量较大的种子,以便种子能快速沉降,减少被带至不宜生长区域的几率。汇泉湾水深7 m[36],该海域的鳗草植株通常生长于水流较急的潮下带,只有质量更大的鳗草种子才能保留下来;而天鹅湖、俚岛湾的鳗草植株生活在2 m以内水流较缓的潮下带[37],种子可以很快沉降,因而这两个地区的鳗草种子较小。
3.2 不同种群海草种子的萌发特性及其关键影响因素
种子萌发是实生苗建成的前提,进而影响整个群体的形成。本研究发现,天鹅湖种群鳗草种子萌发率和实生苗存活率均达40%以上,显著高于汇泉湾种群,分别是其1.6和3.0倍。此外,天鹅湖实生苗规格、光合色素含量和光合速率均为最高值,显著高于俚岛湾种群。Tanner等[39]发现,收集于美国马里兰州和弗吉尼亚州的鳗草种子的室内萌发率为33%,实生苗存活率为20%。Cabaco等[40]将大西洋西部四个不同种群鳗草种子进行室内萌发实验,发现仅有两个种群的鳗草种子萌发,且最高萌发率仅5%,在萌发的种子中,仅15%的种子能实现幼苗建成。综合对比发现,本研究地区及其他不同地区种群的鳗草种子萌发特性和实生苗存活存在显著差异,这可能与种子规格及营养贮存物质的含量有关[11-13, 41]。
本研究发现,鳗草种子萌发率、实生苗存活率与种子质量、长径、短径等形态学指标呈负相关,该结果与其他学者的研究存在不同。Jarvis等[42]认为,大规格的鳗草种子具有更高的活力、更短的萌发时间和更高的萌发率。Combs等[26]也发现,与小规格的种子相比,大规格的鳗草种子可以获得更多的能量资源,从而提高种子萌发率。而Jørgensen等[27]发现,鳗草种子的规格对出苗率和出苗时间无明显影响,但质量较大的种子寿命更长,产生的幼苗生物量显著提高。这些结果表明,在种子萌发期和幼苗初始发育过程中代谢贮存物的调动及其含量可能起到重要作用。
在本研究中,鳗草种子营养贮存物含量为总糖>总蛋白>总脂肪,其中各种群种子总糖和总蛋白含量存在显著差异,并以天鹅湖种群最高,分别达到62%和16%。对比发现,天鹅湖种子的总糖含量高于日本冈山Hinase湾的鳗草种子(粗糖含量约48%)[43]及美国加利福尼亚Hadiona泻湖的鳗草种子(糖含量约51%)[44]。此外,尽管俚岛湾的鳗草种子总蛋白含量最低,仅13%,但仍高于日本Otsuchi湾的鳗草种子(蛋白含量约9%)。Belzunce等[45]认为,大洋波喜荡草(Posidoniaoceanica)种子的内部营养贮存物质越充足,其幼苗存活及初始生长发育越好。本研究发现,鳗草种子的总糖含量越高,种子萌发率和实生苗存活率也越高,实生苗生长更好。
通过主成分分析和曲线拟合发现,种子总糖对鳗草种子的萌发与实生苗生长起主要作用,鳗草种子萌发率及实生苗存活率与种子总糖含量呈显著的线性正相关,糖含量较高的种子具备更高的萌发力。黄升谋等[46]发现,小麦(Triticumaestivum)种子萌发前期需要消耗大量的糖,以保证萌发时有足够能量及结构物质来合成新的细胞和细胞器。蒋敏明等[47]发现,水稻(Oryzasativa)种子总淀粉积累量与发芽率、发芽指数及活力指数显著相关。对于海洋植物种子的报道中也有类似发现。Kuo和den Hartog[48]指出,鳗草种子的下胚轴仅含有少量蛋白质,但淀粉含量却很高,淀粉在种子萌发和幼苗发育过程中起主要供能作用。Taylor[49]也发现,种子总糖是鳗草种子萌发早期主要的能量来源,在种皮破裂和下胚轴伸长过程中起关键作用,充足的糖含量才能保证种子萌发。鳗草种子在萌发过程中呼吸作用逐渐增强,作为鳗草种子的主要营养贮存物,充足的糖能为种子提供足够的呼吸作用底料,以支持萌发过程所需的大量能量,使种子萌发成功的几率大大提升。此外,糖也为幼苗形成初期提供必须的能量及养料。Venable[50]发现,种子的营养贮存物质所占比例越大,幼苗可获取的能量就越多、存活率越高。孙德兰等[51]发现,在莲(Arabidopsisthaliana)幼苗未具备自养能力之前,其生长主要依靠种子内部贮存的淀粉粒,淀粉粒不仅提供能量,其包含的DNA也为幼苗形态建成提供了核苷酸等原料。Sugiura等[52]发现,鳗草种子内部的淀粉降解产物不仅是胚芽突破种皮的主要能量来源,也是实生幼苗发育的必需代谢产物,对鳗草实生苗的建成与生长起决定性作用。鳗草种子糖含量越高,供给实生幼苗生长的能量及合成原料就越充足,幼苗就会产生更长的根系,具有更高的水分和养分吸收能力,从而利于植株的建成[53]。
本研究还发现,种子萌发率及实生苗存活率与种子质量存在显著的线性负相关关系。宗文杰等[54]发现,在青藏高原地区,菊科植物小规格种子具有更快的萌发速率和更高的萌发率,由此来适应高寒草甸环境。刘振恒等[55]发现,较小的种子可以通过迅速萌发的方式,优先占据资源及生存空间。本研究中天鹅湖、俚岛湾的年水温变化幅度较大,鳗草叶片通常于冬季脱落。在这种环境下,规格较小的种子更快萌发,从而获得竞争优势。
4 结语
探明不同种群鳗草种子的形态学特征及萌发特性,是实现优质种源筛选及成功开展海草种子种植的首要关键。本研究结果发现,山东半岛不同种群的鳗草种子在形态学特征和萌发特性上存在显著差异,种子萌发及幼苗建成的关键在于种子内部的糖含量。因此,在筛选种源时,可挑选糖含量高的鳗草种子作为海草床修复的材料。由于种子萌发与幼苗生长不仅受内在因素的影响,还受到外界多种环境因素的影响,关于不同种群所处的环境条件与种子内部营养物质含量对种子萌发及幼苗生长的影响是否存在交互作用等科学问题,还需进一步探讨。
猜你喜欢
幼儿画刊(2022年8期)2022-10-18
金桥(2021年8期)2021-08-23
小读者之友(2020年10期)2020-11-06
祝您健康(2020年5期)2020-05-14
科技与创新(2016年22期)2017-03-30
科技视界(2016年20期)2016-09-29
小朋友·快乐手工(2015年5期)2015-06-06
河北渔业(2015年1期)2015-01-19
河北渔业(2015年1期)2015-01-19
长江蔬菜·学术版(2014年12期)2015-01-08
