
AMF与根瘤菌对间作大豆光合与呼吸代谢的影响
2022-03-25 04:28:32吴艳芬刘秋鸣刘卫欢蒙爱萍陈振翔
广西师范大学学报(自然科学版) 2022年2期
吴艳芬, 刘秋鸣, 刘卫欢, 蒙爱萍, 陈振翔, 刘 灵*
(1.珍稀濒危动植物生态与环境保护教育部重点实验室(广西师范大学),广西 桂林 541006; 2.广西师范大学 生命科学学院,广西 桂林 541006)
当今世界,农业生产面临着土地资源紧缺的问题。间作是在同一时期同一田地按照一定的宽窄比例分行或分带种植2种或2种以上作物的种植方式[1],具有改善植物根际土壤质量[2]、促进植物养分吸收、增加产量[3]等优势,是提高土地资源利用率的有效手段[4],并已得到广泛应用。豆科与禾本科植物(以下简称豆/禾)的间作模式是近年来的研究热点之一[5]。李淑敏等[6]发现,在豆/禾间作系统中,豆科作物根系与固N微生物形成的共生体系可固定大气中的N素,不仅供自身吸收利用,还可转移到间作作物中供其生长。但也有研究表明,在豆/禾间作系统中处于低生态位的大豆因受高生态位作物的遮荫作用,其光合速率和光能利用率降低,最终产量降低[7-8]。
丛枝菌根真菌(arbuscular mycorrhizal fungi, AMF)能与80 % 以上的陆生植物形成共生关系[9]。菌根共生体系中,AMF的生长繁殖是通过内生菌丝吸收植物光合作用产生的糖类,将其合成脂类,再传递给根外菌丝[10],根外菌丝还可在不同植物间形成庞大的网络结构[11]。植物生长可通过根系和AMF的根外菌丝吸收土壤中的养分,尤其是移动性差的P、Zn、Cu等[12]。研究表明,AMF可改善植物对矿质营养的吸收[13],增强植物抗逆性[14],调节土壤团聚体结构[15],增加土壤酶活性[16]等。
根瘤菌(rhizobium, Rh)可与豆科植物共生形成根瘤,固定大气中N素供植物利用,减少N肥施用量[17]。在豆科/非豆科植物(以下简称豆/非豆)间作系统中,双接种AMF和Rh(以下简称双接种)形成AMF-寄主植物-Rh共生体系[11,18],可提高AMF侵染率,增加P素利用率,增强结瘤率和固N能力,促进宿主植物养分吸收,提高生物量和产量[19-20]。
目前,将不同AMF和Rh双接种于豆/禾间作系统已成为菌根真菌的研究热点之一,但鲜见田间条件下,双接种于豆科与木本经济作物间作系统的研究。金橘Fortunellamargarita(Lour.) Swingle是广西优势特色水果和地理标志产品之一,具较高食药用价值和经济价值。本文拟探究接种幼套球囊霉和费氏中华根瘤菌对金橘/大豆(以下简称橘/豆)间作系统中大豆Glycinemax光合与呼吸代谢和植株生长的影响,旨在为化肥减施及金橘大豆增产提供理论依据。
1 材料与方法
1.1 试验材料
金橘:一年生金橘嫁接苗(品种:阳朔金橘),购自广西阳朔白沙镇农贸市场。大豆品种为桂春豆108号,购自广西农科院。接种的大豆根瘤菌(rhizobium, Rh)为费氏中华根瘤菌Sinorhizobiumfredii(简写为R),菌种编号:1.227,购自广东省微生物菌种保藏中心;AMF菌种为幼套球囊霉Glomusetunicatum(简写为A),编号:BGC GZ03C,购自北京市农林科学院。
1.2 试验设计
试验地位于广西师范大学雁山校区生物园,总面积为1 050 m2(35 m×30 m)。
设以下2个栽培系统8个处理。其中,间作系统——1)金橘大豆间作(对照1):F+S;2)金橘大豆间作接种幼套球囊霉:F+S+A;3)金橘大豆间作接种费氏中华根瘤菌:F+S+R;4)金橘大豆间作双接种幼套球囊霉和费氏中华根瘤菌:F+S+A+R。单作系统——5)大豆单作(对照2):S;6)大豆单作接种幼套球囊霉:S+A;7)大豆单作接种费氏中华根瘤菌:S+R;8)大豆单作双接种幼套球囊霉和费氏中华根瘤菌:S+A+R。
每处理设3个重复,每重复的8个处理(每处理对应1个小区)综合为1个大区,每小区面积为30 m2(5 m×6 m),大区面积为270 m2。各大区之间留宽1 m,深0.6 m的大沟;各小区间距为0.5 m,试验地周围种植大豆作保护行。金橘按行距×株距为1.5 m×1.5 m种植,每行金橘与第1行间作大豆间距为60 cm,然后2行大豆间按株距×行距为20 cm×30 cm,播种沟深5 cm,每穴3粒种子,每行金橘1侧共种植2行大豆,依此类推。
基肥用湖北中农中加贸易有限公司的状元峰牌复合肥(mN∶mP∶mK=17∶17∶17,简称“复合肥”),施肥沟各距种植行两侧0.15 m,按1/2的常规推荐施肥量(即大豆3 g/株,金橘300 g/株)[21-22]。每月追肥1次,按2/3的常规推荐施肥量(即大豆4 g/株,金橘400 g/株),水肥一体化喷灌浇水。其余常规田间管理。
1.3 接种方法
金橘接种A:在40 cm×40 cm×40 cm的种植穴内填入土壤,深度约13 cm时,撒入含孢子、根段和沙子的A混合菌剂1 000 g/株,与表土混匀后,将金橘幼苗根系展开,填入土壤至距嫁接口5 cm左右,浇透定根水。未接种处理为加入等质量经灭菌处理的同种混合菌剂。
大豆接种A:大豆长到三叶期时,在距大豆种植行7.5 cm两侧各挖接种沟2条,每行均匀撒入含孢子、根段和沙子的A混合菌剂500 g(约20 g/株),与表土混匀后填土并镇压。未接种处理为加入等质量经灭菌处理的同种混合菌剂。
大豆接种R:采用拌种法。将约500 mL R菌悬液(活菌数约2×109cfu/mL)浸泡大豆种子20 min,捞出阴干后种植。每穴浇灌经离心浓缩并用无菌水重悬配制好的同等浓度R菌悬液10 mL。未接种处理为加等量无菌液体培养基。
1.4 采样分析
叶片采样时,每处理每重复各取生长一致的大豆植株5株,每株分别取上、中、下均为同一叶位的叶子各2片,装入干净的自封袋中,标记后立即置于冰盒,尽快带回实验室。大豆叶绿素含量按照李合生[23]方法测定,线粒体膜H+-ATP酶活性按照李玲[24]方法测定,琥珀酸脱氢酶(succinate dehydrogenase, SDH)活性按照刘家尧等[25]方法提取,并按南京建成生物工程研究所试剂盒说明书的方法测定。用LI-6400便携式光合仪测定不同处理大豆的同一叶位、叶龄基本一致叶片的光合速率。作物生长期间充分供水以满足作物对水分的需求,生长80 d后收获大豆,对大豆根瘤称鲜质量和计数。将地上部与根部分开,植物样品放入牛皮纸袋,置于烘箱中105 ℃杀青30 min,然后再调至70 ℃烘干至恒质量,最后称干质量。生物量为单位面积(m2)内所有大豆植株总干质量。大豆产量按照汤复跃等[26]方法(稍作修改)测定。
根系采样按照朱国政等[27]方法稍作修改:每处理的每重复随机选择5株大豆植株挖出,切勿伤及根部,取含根尖的幼嫩细根装入含FAA固定液的离心管后,按照Phillips等[28]方法进行根样染色,按照Trouvelot[29]方法计算根系菌根侵染强度及根系泡囊丰度。
1.5 数据分析和绘图
数据用SPSS 19.0软件进行ANOVA方差分析,并用Duncan新复极差法进行多重比较,不同小写字母表示各处理间差异显著(P<0.05)。用Sigmaplot 13.0软件进行绘图。
2 结果分析
2.1 不同处理对大豆根系菌根及根瘤发育的影响
如表1所示:1)间作系统中各处理的大豆根系菌根侵染强度(mycorrhizal infection intensity in root system)ηm在单接种A或R后均比F+S处理增加,3个间作接种处理的ηm值由大到小依次为间作双接种F+S+A+R处理、间作单接种F+S+A处理、间作单接种F+S+R处理,处理间差异显著(P<0.05);单作系统中各处理的ηm值也具有与间作系统中各处理相似的变化趋势。与单作系统各处理相比,间作系统中相对应各处理的ηm均显著(P<0.05)降低,其中,单作双接种S+A+R处理ηm值最高,间作双接种F+S+A+R处理ηm值第2,两者间差异显著(P<0.05)。2)间作系统的大豆根系泡囊强度(mycorrhizal vesicle intensity in root system)ηV,不论在单作还是间作系统中,各处理ηV的变化趋势与ηm的变化趋势相似。且除间作双接种F+S+A+R处理的ηV值比单作双接种S+A+R处理显著(P<0.05)增加外,其余间作系统各处理ηV与其相对应的单作系统各处理相比均降低,但差异不显著(P>0.05)。3)4个不接种A处理(F+S、S、F+S+R、S+R)的ηm和ηV值均较低,说明土壤中有极少量土著AMF存在,但不占优势,接种R对土著AMF影响不显著,但对外源AMF的发育有显著促进作用。

表1 不同处理对大豆根系菌根及根瘤发育的影响
大豆根瘤数量和干质量(the number and dry weight of nodules,简写为NN&DWN)见表1: 1)单作系统中各处理根瘤NN&DWN的变化趋势与ηm的变化趋势相似。而在间作系统中,3个间作接种处理的变化趋势由大到小依次为双接种F+S+A+R处理、单接种F+S+R处理、单接种F+S+A处理,各处理间差异显著(P<0.05)。与单作系统各接种处理相比,间作系统相对应各处理的NN&DWN均显著(P<0.05)降低,说明间作对根瘤的生长发育有一定的抑制作用。2)无论间作还是单作系统,外源R与外源A相互促进对方的发育,双接种处理均比相应的单接种处理显著促进大豆根瘤的发育。此外,4个不接种R处理(F+S、S、F+S+A和S+A)均有根瘤形成,说明土壤中也有土著Rh存在。单作单接种S+A处理的NN&DWN甚至显著高于单作单接种S+R处理,说明S+A处理显著促进土著Rh的发育,推测接种的外源R与土著Rh间有竞争关系。3)间作系统中,单接种外源A的F+S+A处理对土著Rh的NN&DWN比F+S处理也有显著的促进作用,但因间作处理的抑制效应,其促进作用减弱。单接种外源R的F+S+R处理则比F+S+A处理的NN&DWN升高,但仅根瘤数量差异显著(P<0.05),根瘤干质量差异不显著(P>0.05);间作双接种F+S+A+R处理的NN&DWN最高,单作双接种S+A+R处理的次之。综上,不论在单作还是间作系统中,接种A处理均可显著促进R的发育,双接种处理的R发育状况最好。
2.2 不同处理对大豆叶绿素含量和净光合速率的影响
如图1a所示,间作系统中各接种处理的叶绿素含量均高于不接种F+S处理,且叶绿素含量由大到小依次为双接种A+R处理、单接种A处理、单接种R处理,处理间差异显著(P<0.05)。单作系统中,各处理的叶绿素含量也具有与间作系统各处理相似的变化趋势,且间作系统各处理与相对应的单作系统各处理相比叶绿素含量均显著(P<0.05)降低;大豆净光合速率(图1b)与叶绿素含量的变化趋势相似,但间作系统各处理与相对应的单作系统各处理相比均升高,且仅间作双接种F+S+A+R处理与单作双接种S+A+R处理间差异显著(P<0.05)。
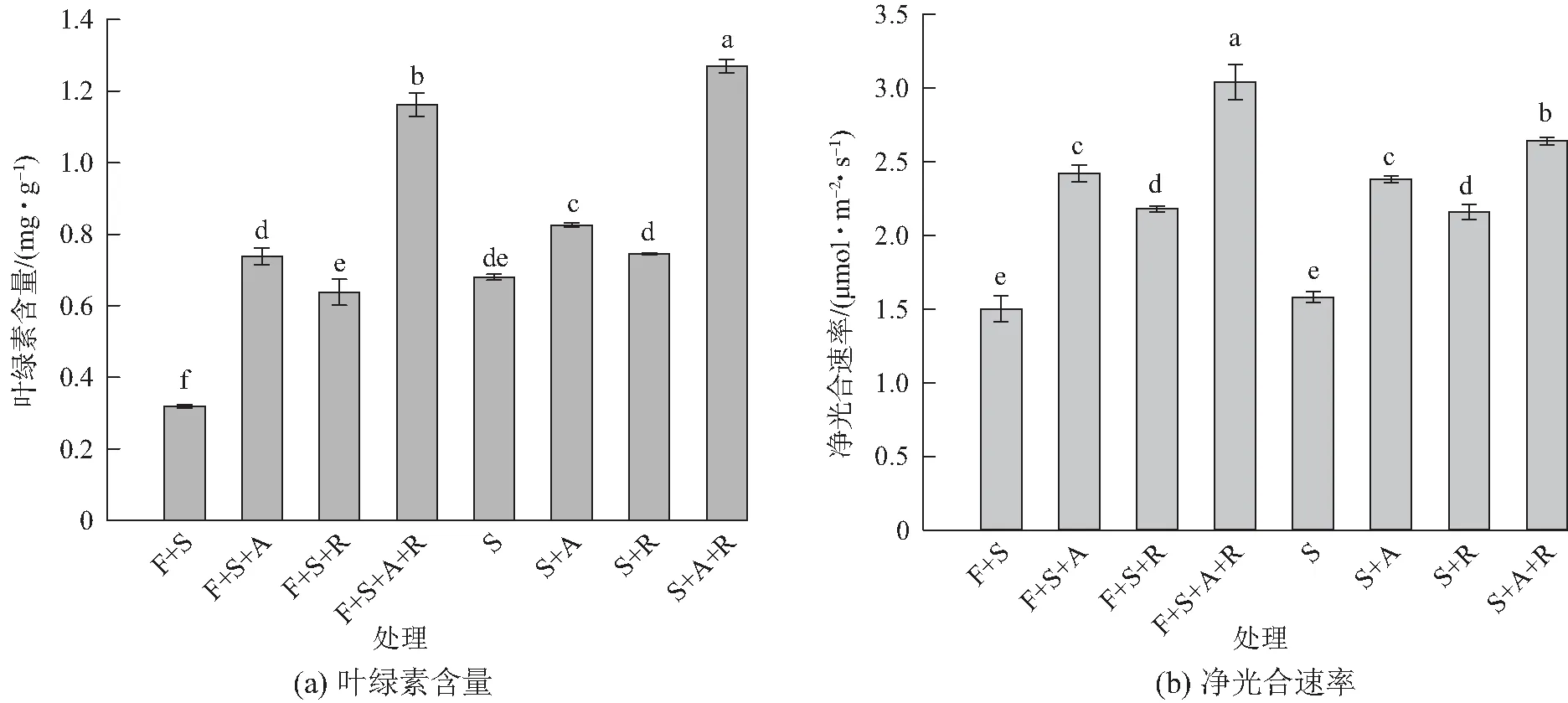
不同字母表示处理间差异显著(P<0.05)图1 不同处理对大豆叶绿素含量和净光合速率的影响Fig. 1 Effects of Glycine max with different treatments on chlorophyll content and net photosynthetic rate
2.3 不同处理对大豆叶片琥珀酸脱氢酶(SDH)和线粒体膜H+-ATP酶活性的影响
间作系统各接种处理大豆的叶片SDH活性均比相应的单作系统各接种处理高,其中,间作双接种F+S+A+R处理的SDH活性最高,间作单接种F+S+A处理次之,两处理间差异显著(P<0.05)(图2a)。间作系统各接种处理大豆叶片H+-ATP酶活性与叶绿素含量有相似的变化趋势,但处理间差异均不显著(P>0.05)(图2b)。单作系统各接种处理均比间作系统相对应的各接种处理高。在两大系统中,所有接种处理的H+-ATP酶活性,除双接种S+A+R与F+S+A+R处理、单接种S+R与F+S+R处理差异显著(P<0.05)外,其余相对应的各处理间差异均不显著(P>0.05)。
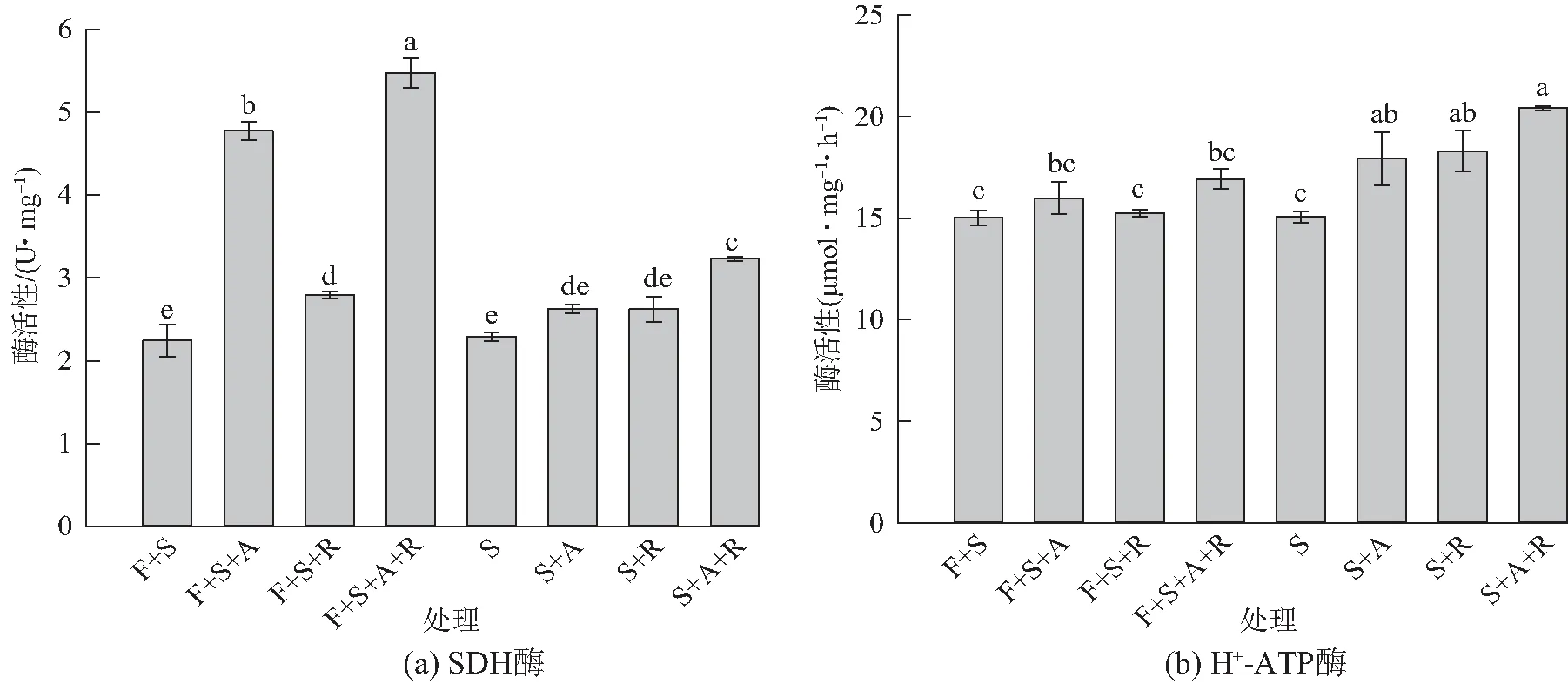
不同字母表示处理间差异显著(P<0.05)图2 不同处理对大豆叶片SDH、H+-ATP酶活性的影响Fig. 2 Effects of Glycine max with different treatments on succinate dehydrogenase and H+-ATPase activities
2.4 不同处理对大豆产量和植株生物量的影响
如表2所示,与大豆净光合速率的变化趋势相似,无论是单作还是间作系统,各接种处理的大豆生物量和产量均显著(P<0.05)高于相对应的不接种对照S和F+S处理,其接种效应由大到小均表现为双接种A+R、单接种A、单接种R,且处理间差异均显著(P<0.05)。间作系统各处理的生物量和产量与单作系统各处理相比,除F+S处理相对于S处理、F+S+R处理相对于S+R处理差异均不显著外(P>0.05),其余间作系统各处理均比相对应的单作系统各处理显著(P<0.05)增加。所有处理中,间作双接种F+S+A+R处理的生物量和产量最大。
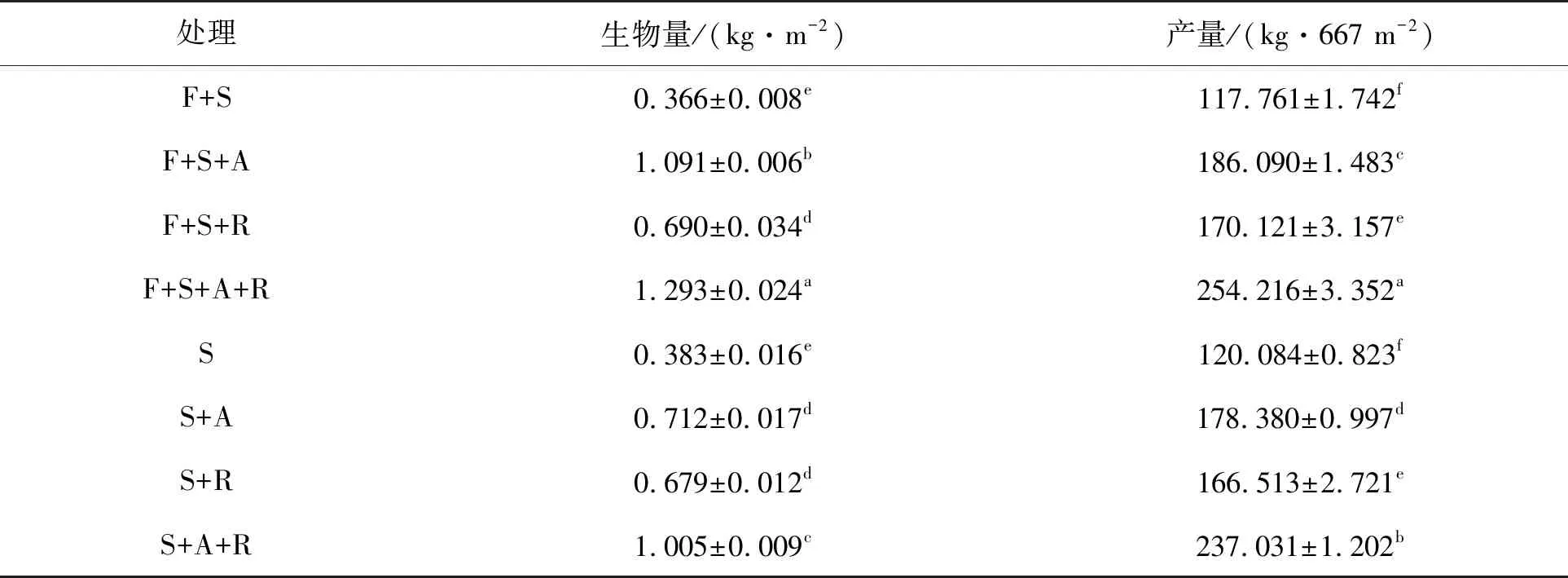
表2 不同处理对大豆生物量及产量的影响
3 讨论
3.1 AMF或Rh对间作大豆根系菌根及根瘤发育的影响
菌根共生体中,AMF的根外菌丝与寄主植物根系形成连接根与土壤的菌丝网络,其根外菌丝广泛伸入土壤,能扩大养分吸收范围,所分泌氢离子和有机酸(例如草酸、柠檬酸、苹果酸、琥珀酸等)可溶解土壤矿物,提高P、K、Ca、Mg的生物有效性等。这也可能是寄主植物(例如柑橘)和Rh提供自身所需的N、P、K、Mg、Cu、Zn等养分含量和吸收量增加的原因之一[22,25,33]。有研究表明,豆科植物和Rh的生长发育及共生固N作用受所居住土壤中营养状况的直接影响[35]。1)土壤N水平影响AMF与Rh的相互作用。施N可降低根瘤的形成,但丛枝菌根的形成可在一定程度上抵消因N增加对根瘤产生的抑制作用[40]。豆/禾作物,例如玉米/蚕豆间作系统双接种处理,能提高豆科植物(蚕豆)自身及与之间作的禾本科植物(玉米)的多种养分含量。其中,间作系统中禾本科作物生物量增加较大,对土壤N竞争较强[6],因此又可引起豆科作物固N能力增加[41],N转移增加[42],产生“N阻遏”消减作用,实现N高效利用[43]。单接种AMF或Rh后大豆向与其间作的玉米转移20%~43%的N,双接种处理则比单接种AMF时大豆向玉米转移的N提高45%[44-45],双接种具有协同增效作用[46]。Larimer等[47]认为适量施肥有利于大豆根瘤的形成和生长,但也有施N量低至30.38 kg/hm2(常规施N量为330 kg/hm2)时也显著抑制大豆根瘤形成和生长的报道[48]。大豆与玉米套作时大豆根瘤数及根瘤鲜质量显著下降[49],易受AMF侵染的植物对施N较为敏感,AMF只有在一定施N量的基础上才能促进作物生长,但施N量过高也会影响AMF侵染[43]。2)低P是根瘤形成的主要限制因子,AMF与Rh的相互作用取决于土壤P含量[40]。当向N、P缺乏的土壤中单独接种Rh,结瘤固N能力较差[50],施加足量的有效P后,AMF为Rh提供其所需要的P,Rh的结瘤固N能力增强[51-52],进而为植物提供更多的N,表明P对于Rh的结瘤固N具有重要的作用。此外,接种AMF促进宿主植物的生长发育,为Rh提供充足的能量,最终将促进植物固N。土壤中有效P含量增多,致使根瘤菌结瘤和固N能力增强[53]。因此,合理施肥十分重要。本文中,不论间作还是单作系统,外源A与R相互促进对方的发育,双接种处理对菌根和根瘤发育均有显著的促进效应。与R处理相比,A+R处理的根瘤数量多,但体积小,且根瘤的比表面积大,与刘忆等[39]的结果相似,意味着两者有协同增效作用,可能更有益于气体交换。
值得注意的是,本文的橘/豆间作系统中,间作处理大豆根系菌根和根瘤的生长发育均受不同程度抑制,尤对根瘤的抑制作用较为显著。但无论单作还是间作,双接种处理下的大豆结瘤数和菌根侵染率均显著高于单接种处理,双接种对大豆生长有协同促进作用,这与文献[20,49,54]的研究结果相似。这可能是间作环境中大豆受高位金橘的隐蔽,光合产物优先供给地上部生长,造成了对大豆根瘤数和菌根发育的调控[55];也可能是金橘施用的N肥在雨水和灌溉的作用下运移到大豆间作带,影响了大豆结瘤状况。本文认为,金橘和大豆2种作物间竞争生态位和水、肥等资源,使大豆结瘤能力和菌根侵染能力均低于单作处理,表现出抑制效应。此外,间作系统中大豆根系根瘤的侵染受抑制,可能与接种方式不同以及接种量不足有关。由于进行的是田间实验,需要R菌剂剂量较多,且对R菌剂进行了稀释。例如,李淑敏等[6]在盆栽蚕豆/玉米间作系统中,每盆接种20 mL Rh菌液(Rh菌数为16.4×109cfu·mL-1·盆-1);刘忆等[39]在紫花苜蓿根部浇灌75 mL Rh菌液(Rh菌数为50×109cfu·mL-1·穴-1)。本文中接种所用的R菌数不及文献[6,39]施用菌数的1/3,加上土著Rh的竞争,可能是R接种效果不理想,根瘤生长发育受抑制的原因。今后将参考文献[6,39]的方法增加R接种剂的浓度和体积来促进根瘤的侵染,同时适当提高土壤有效P和有效N含量来增强根瘤及菌根的生长和发育。
3.2 AMF或Rh对间作大豆光合、呼吸相关代谢的影响
叶绿素是光合色素,其含量和净光合速率可反映植物光合作用强弱,而光合作用影响植物的物质代谢和能量转化,对植物生长有重要意义[5]。本文间作系统中,大豆叶绿素含量均降低,可能与其合成受光、温度、矿质元素等因素影响,而橘/豆间作导致大豆在对光及矿质元素的竞争中处于劣势有关。但间作系统中各接种处理的大豆净光合速率均高于对应的单作接种处理,且间作双接种处理的值最大,从另一个角度说明A和R二者混合接种具有协同效应,更有益于大豆生长。这可能是因为净光合速率受多种因素(包括光、CO2浓度、水分、温度、作物的呼吸强度等)影响。对于AMF-Rh-植物共生系统,虽是以微生物消耗光合有机产物为前提,但AMF和Rh的侵染不仅刺激了其共生植物对养分的需求,继而提高共生植物的光合速率,还弥补了微生物对养分的额外消耗[56]。李智等[57]发现,间作状态下处于强竞争势力的谷子其叶绿素含量和净光合速率显著高于单作谷子,而处于弱竞争势力的绿豆叶绿素含量和净光合速率却低于单作。邢易梅等[58]发现,双接种G.m和Rh显著提高了紫花苜蓿叶绿素含量且效果优于单接种。
SDH是植物有氧呼吸过程中参与三羧酸循环的限速酶,而呼吸作用释放能量,是植物进行各种生理代谢的基础,对植物的生长发育有重要意义[59]。本文间作系统中,F+S处理的大豆SDH活性显著降低,可能是大豆受到间作金橘的遮阴作用,日照强度和时间减少、漫射光比例增大、大豆冠温降低、湿度增大,而导致呼吸作用降低。Zhao等[60]发现,间作遮阴处理较单作相比,茶树的呼吸强度显著降低,与本文得出的结论一致。此外,本文间作系统大豆接种A或R后,其SDH活性均显著增加,双接种效果显著优于单接种,且间作系统接种处理的大豆SDH活性大于对应单作系统接种处理。这可能与间作系统有更强大的菌根网络结构,菌根植物的矿质营养浓度较高,且豆科植物-Rh共生体系中,Rh可在根瘤体内发育成类菌体,并在呼吸作用中为固N体系提供还原力和ATP,增加共生植物的呼吸代谢[61]有关。刘智蕾[62]和Naseem等[63]分别发现,AMF可增加水稻在低温胁迫下的呼吸代谢,Rh可提高水分胁迫下植株的呼吸强度。可见,接种AMF或Rh可促进共生植物的呼吸代谢。
H+-ATP酶是广泛存在于线粒体内膜、质膜、类囊体膜上的一类ATP泵酶蛋白[64],在养分的吸收和运输、维持细胞稳态、增加植物抗性、物质跨膜运输提供能量等方面发挥重要作用,是植物生长发育过程中的动力来源[64-65]。本文发现,间作系统中大豆的H+-ATP酶活性均低于其对应的单作处理,或许是因为橘/豆间作时,大豆处于竞争劣势,导致该酶活性降低,接种A或R后则活性增加,双接种处理促进效果最好。但接种后促进效应的机制尚不清楚,因为H+-ATP酶受多条代谢途径调控。本文推测,接种A或R可能影响某些信号分子,例如生长素(IAA)、乙烯(ET)、活性氧(ROS)含量以及质膜 NADPH 氧化酶、抗氧化酶[64]活性等,促进了H+-ATP酶的表达,其详细机制有待于进一步研究。
3.3 AMF或Rh对间作大豆干物质积累量的影响
刘忆等[39]研究表明,单作紫花苜蓿接种AMF或Rh分别形成根瘤和菌根后,地上部生物量比对照显著增加,且以双接种处理最高,说明双接种AMF处理和Rh处理后,AMF和Rh消耗的光合产物低于增加的光合产物,使紫花苜蓿生物量和产量增加;Haro等[66]也发现双接种具有协同效应,且联合作用的优势远大于单独接种。间作系统中往往存在优势和劣势竞争力,处于竞争劣势的作物受到竞争优势作物的竞争作用而影响生长[67]。在保证作物各项生理活动正常的前提下,协调好光合与呼吸代谢的平衡,是作物获得高产的基础[60]。本文间作系统中,接种处理的大豆生物量和产量均高于相应的单作接种处理,且双接种处理的促进效果显著优于单接种,可能是金橘的竞争作用改变了间作大豆原有的生长环境,其光合和呼吸代谢受到不利影响,光合产物的合成与分配也受影响,降低了生物量和产量。但接种的A或R为土壤有益微生物,可改善间作大豆的养分吸收,促进光合和呼吸相关代谢,进而提高生物量和产量。李淑敏等[6]在蚕豆和玉米间作系统中接种AMF和Rh,发现根系不分隔间作体系中双接种处理显著增加了玉米和蚕豆的生物量,且促进效果优于单独接种。周小宇等[68]用AMF (G.m)与Rh对玉米和饭豆间作系统接种,发现G.m与Rh混合接种更能提高间作饭豆的产量。
4 结论
间作大豆接种幼套球囊霉或费氏中华根瘤菌后不同程度地促进了菌根和根瘤共生,促进其对土壤中养分的吸收以及光合和呼吸相关代谢和生长,最终提高产量,且间作双接种处理的促生效果最好。接种幼套球囊霉或费氏中华根瘤菌对作物生长的促进效应有助于改良金橘果园生态系统,提高经济收益。
猜你喜欢
广西林业科学(2022年2期)2022-05-09 07:49:54
中国南方果树(2022年1期)2022-01-28 07:39:16
农技服务(2021年7期)2021-09-24 04:13:02
河北果树(2020年1期)2020-02-09 12:31:36
山东农业科学(2019年11期)2019-12-24 01:11:27
植物营养与肥料学报(2019年11期)2019-12-13 05:55:04
饮食保健(2019年18期)2019-09-20 08:03:34
江苏农业科学(2019年11期)2019-07-22 01:32:45
家庭科学·新健康(2019年12期)2019-01-13 01:30:56
干旱地区农业研究(2017年5期)2017-12-18 06:18:18
