
松褐天牛的蛀道结构系统研究
2022-03-25 08:47:04高尚坤马深成杨化伟杨庆贺周成刚乔鲁芹
林业科学研究 2022年2期
高尚坤,马深成,杨化伟,杨庆贺,周成刚,乔鲁芹
(1.山东农业大学植物保护学院,山东省林业有害生物防控工程技术研究中心,山东 泰安 271018;2.泰安市泰山森林病虫害防治检疫站,山东 泰安 271000;3.山东省农业机械科学研究院,山东 济南 250100;4.山东农业大学后勤管理处,山东 泰安 271018)
松褐天牛(Monochamus alternatusHope),又名松墨天牛,属鞘翅目(Coleoptera)天牛科(Cerambycidae)墨天牛属(Monochamus),是松属植物的重要钻蛀性害虫[1]。松褐天牛还可通过成虫补充营养和刻槽产卵携带传播松材线虫,是松材线虫病自然扩散蔓延的重要媒介昆虫,现已造成大量林木死亡,给我国践行“绿水青山就是金山银山”理念和推进林业生态建设造成了极大的困难[2-3]。目前,松褐天牛的防控技术主要集中利用引诱剂监测和诱杀部分成虫[4],喷洒化学药剂防治成虫[5-6],预防区释放花绒寄甲(Dastarcus helophoroides)老熟幼虫、蛹或释放肿腿蜂(Sclerodermussp.)防治低龄幼虫[7-8]等。受限于森林生态系统的复杂性和松褐天牛生活的隐蔽性,寄主植物—松褐天牛的营养关系研究进展较为缓慢[9-10],且主要集中在裸露生活的成虫期[11]。松褐天牛整个世代超过2/3 的活动期都在寄主松树的木质部内度过,受外界环境干扰较少,使得其种群很稳定,从外部进行防治极其困难[12]。近年来,利用以天敌昆虫和病原微生物为主的生物防治技术防治天牛类等蛀干害虫得到广泛应用[2]。天敌昆虫和病原微生物施放后,能够主动爬行或飞行搜寻、定位寄主资源,但林间的控制效果仍不太稳定。另外,树干注射药物防治蛀干害虫也是重要的防治措施之一,该方式见效快、效果明显,但也因某些天牛种类利用蛀屑堵塞虫道,蛀道曲折、方位多样等原因导致效果不佳[13-14]。
蛀道是松褐天牛取食、排泄和躲避天敌的重要空间,也是与天敌互作的主要场所。因此,解析松褐天牛的蛀道结构,弄清其蛀道走向、长度,蛀屑堵塞规律及蛹室位置等钻蛀习性,能够为利用天敌昆虫、病原微生物和注射药物进行科学、高效和精准地防控提供依据。
1 材料与方法
1.1 调查地点
调查地点位于威海市环翠区羊亭镇北上夼村的松材线虫疫木处理场(37°15′ N,121°51′ E),属低山丘陵区,树种为赤松(Pinus densifloraSiebold &Zucc.),黑松(P.thunbergiiParl.)和栎类(Quercussp.),平均海拔为70~100 m,属温带季风气候,夏无酷暑,冬无严寒,年均气温12 ℃,年均降水量800 mm,年平均日照2 569 h。
1.2 调查方法
2018 年11 月威海地区松褐天牛老熟幼虫已进入木质部,且做好蛹室,不再取食,进入越冬状态。于疫木处理场随机选取松褐天牛危害的木段,记录树种,测量木段直径,然后将树皮剥掉,根据侵入孔的位置上下各保留20 cm 左右,将树干锯成段,用记号笔在横切面标记上下方位,用斧头和裁纸刀将其一一劈开,对完整的蛀道进行编号。用游标卡尺分别测量每个蛀道的侵入孔内径、蛹室内径、蛀道长度、蛀道内径和蛀屑堵塞长度,并将蛀道内松褐天牛幼虫和蛀屑取出带回实验室,用分析天平称质量。将蛀屑置于烘箱中于80℃下烘干24 h,再次称质量,计算蛀屑含水率。侵入孔内径(mm)为扁圆形侵入孔的最大宽度;羽化孔(蛹室)内径(mm)为羽化孔或蛹室的最大宽度;蛀道长度(mm)为侵入孔至羽化孔(蛹室)坑道外径长度;蛀道内径(mm)为蛀道最深处与树干平行部分的内径长度;蛀屑堵塞长度(mm)为蛀道内蛀屑的长度;蛀屑含水率%=(烘干前蛀屑质量-烘干后蛀屑质量)/烘干前蛀屑质量 × 100%。侵入孔与蛹室的相对方位是以侵入孔为基准,判断羽化孔在侵入孔的上方或下方。
1.3 数据分析
利用Microsoft Excel 97-2003 工作表记录相关数据,利用GraphPad Prism 5.0 分析2 种树种内松褐天牛蛀道长度与所处木段直径的相关性,松褐天牛幼虫的个体大小与蛀食特征的关系,松褐天牛侵入孔与蛹室的相对方位和2 种蛀屑含水率的差异性。
2 结果与分析
2.1 松褐天牛的钻蛀特点
大多数松褐天牛幼虫在3 龄后钻入木质部,其蛀道结构类型单一,大体呈“U”形。在侵入孔端有一小段弯曲,呈“S”形。坑道体积存在一定差异,蛀道间互不侵犯;松褐天牛幼虫钻蛀过程中具有边蛀食边将蛀屑连同粪便一起推出坑道的习性,蛀屑在树皮下的堆积压力造成蛀道侵入孔处一小段堵塞,且堵塞较为紧实;松褐天牛老熟幼虫在越冬前于蛀道末端咬食出蛹室,翌年化蛹。蛀道的侵入孔位置在蛹室上下方位均有分布;蛀道内部表面整体较粗糙,但无较大起伏(图1)。

图1 松褐天牛的钻蛀特点Fig.1 Boring characteristics of Monochamus alternatus
2.2 树种及直径对蛀道长度的影响
由图2 可知,赤松和黑松木质部内松褐天牛的蛀道长度存在一定差异,与所在木段直径的关系不大。其中黑松内松褐天牛蛀道长度随所在木段直径的增加而增加,而赤松内松褐天牛蛀道长度随所在木段直径的增加变化不大。黑松内松褐天牛的蛀道长度稍长于赤松内松褐天牛蛀道长度,表明松褐天牛幼虫对黑松的危害程度比赤松更高。

图2 树种差异对松褐天牛蛀道长度的影响Fig.2 Effects of tree species on the gallery length of Monochamus alternatus
2.3 松褐天牛个体大小与蛀食特征的关系
调查中发现蛀道内的松褐天牛幼虫体色呈乳黄色,已不再取食,虫体脂肪储量明显增加,包括蛹室在内的蛀道系统结构也已成形,表明越冬前的松褐天牛老熟幼虫已基本完成蛀食阶段,其能量储备能够满足越冬和翌年化蛹所需,此时,松褐天牛幼虫的个体大小和蛀食特征较为稳定。
由图3~图7 可知,越冬前松褐天牛幼虫的个体大小为0.2~0.8 g,差异明显,但与蛀屑堵塞长度、侵入孔内径、蛹室内径和蛀道长度等蛀食特征的相关性均较小,表明松褐天牛幼虫在木质部的蛀食规律比较一致。松褐天牛幼虫在黑松上的蛀食特征(如蛀道内径和蛀道长度)比在赤松上相对明显些(图4、图7),表明松褐天牛对黑松的危害程度比赤松严重。蛀屑堵塞长度随松褐天牛幼虫个体增大呈先下降再上升趋势,(图3),表明松褐天牛在蛀食过程中存在向蛀道外“推粪”习性,而后期不再向外“推粪。”赤松上侵入孔的内径与松褐天牛幼虫个体大小的相关性最大(R2=0.437,图5),表明钻入赤松木质部危害的松褐天牛幼虫个体大小存在一定差异。而黑松上蛹室的内径与松褐天牛幼虫个体大小的相关性最小(R2=0.036,图6),表明危害黑松的松褐天牛幼虫个体大小较为一致。

图3 松褐天牛幼虫个体大小与蛀屑堵塞长度的关系Fig.3 Relationship between body size of larvae and blockage length of frass

图4 松褐天牛幼虫个体大小与蛀道内径的关系Fig.4 Relationship between body size of larvae and inner diameter of galleries

图5 松褐天牛幼虫个体大小与侵入孔内径的关系Fig.5 Relationship between body size of larvae and inner diameter of entrance holes
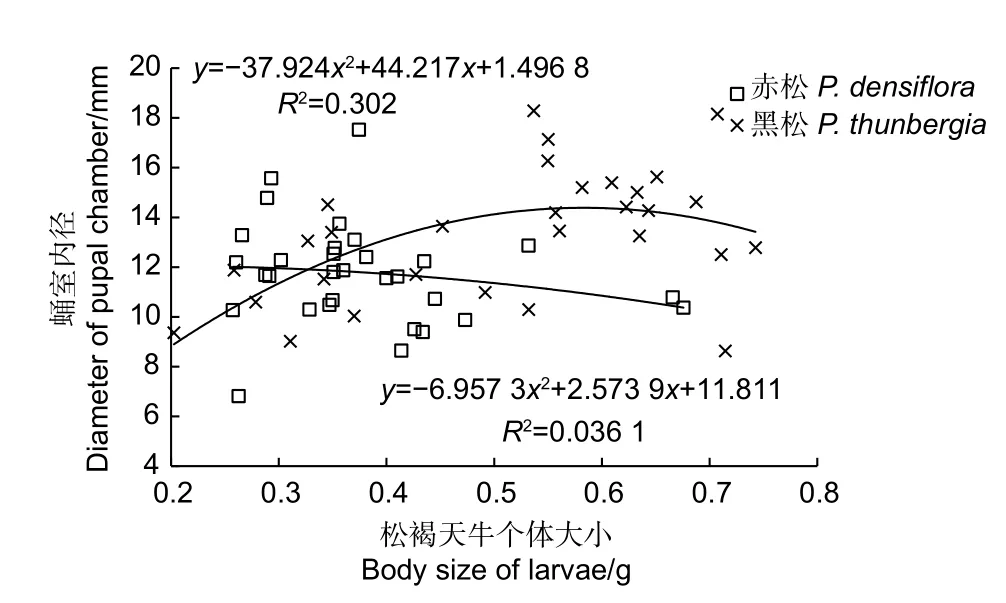
图6 松褐天牛幼虫个体大小与蛹室内径的关系Fig.6 Relationship between body size of larvae and diameter of pupalchambers

图7 松褐天牛幼虫个体大小与蛀道长度的关系Fig.7 Relationship between body size of larvae and gallery length
2.4 侵入孔与蛹室的相对方位
由图8 可知,无论赤松还是黑松,松褐天牛的蛹室在侵入孔上方位的数量均多于蛹室在侵入孔下方位的数量,表明松褐天牛幼虫钻入木质部后更偏好向上蛀食,树种差异对其蛀食方位选择影响不大。

图8 松褐天牛侵入孔与蛹室的相对方位Fig.8 Relative orientation of the entrance holes and pupal chamber
2.5 蛀屑含水率
由图9 可知,赤松和黑松内松褐天牛幼虫堵塞蛀道的蛀屑含水率无显著差异(t=1.626,df=56,P=0.109 6),均在12%~13%之间,表明树种差异对松褐天牛蛀屑含水率影响不大。

图9 松褐天牛幼虫蛀屑的含水率Fig.9 Frass moisture content of Monochamus alternatus larvae
3 讨论
钻蛀类害虫隐蔽的生活史阶段极大地限制了该时段内的生物生态学的研究,这也是钻蛀类害虫难以防治的重要原因之一。蛀道结构反映的是钻蛀类害虫的蛀食特征,弄清蛀道结构,可揭示其在寄主树木中的取食特性,补充完善害虫与寄主植物及其天敌的营养关系。刘奇志等[15]通过研究桃红颈天牛(Anomia bungii(Faldermann))的蛀道结构特点和推粪习性,为提高病原线虫对桃红颈天牛的防治效率提供了科学依据。云斑白条天牛(Batocera lineolateChevrolat)幼虫在核桃(Juglans regiaLinn.)木质部蛀食,蛀道方向朝上,排粪孔朝下,各蛀道前期不相通,后期相通;幼虫多时,各蛀道串连相通,一蛀道内多个幼虫,为施药防治方式提供了依据[16]。咖啡旋皮天牛(Acalolepta cervinus(Hope))韧皮部蛀道和木质部蛀道的空间特征,揭示了其幼虫钻蛀行为具有很强的位置选择性[13]。通过对比弗氏窄吉丁(Agrilus fleischeriObenberger)幼虫在钻天杨(Populus nigravar.italica(Moench)Koehne)和山杨(P.davidianaDode)的蛀道长度等钻蛀特征,表明弗氏窄吉丁对非本地种钻天杨的危害程度更高,更敏感[17]。叶片上豌豆植潜蝇(Phytomyza horticolaGoureau)蛀道的数量一定程度上可以用来反映寄主植物的生长状况和豌豆潜蝇姬小蜂(Diglyphus isaea(Walker))的寄生率及幼虫存活率[18]。Romo 等[19]通过测量Arhopalus ferus(Mulsant)和Prionoplus reticularisWhite 2 种天牛的钻蛀深度,为评估其检疫处理的效率提供了依据。松褐天牛为寡食性昆虫,主要危害松属植物,在山东地区多危害黑松和赤松,表明松褐天牛的生活史可以与2 种松树的物候期相配合,并在一个区域的多种寄主上完成发育和繁育。
同一种蛀干害虫在不同地区同一树种上的蛀食特征多有所差异。6 个不同地区樟子松(Pinus sylvestrisvar.mongolicaLitv.)上松树蜂蛀道特征均存在一定的差异性,蛀道形状为“Z”形、“U”形或“L”形等多种类型[20]。锈色粒肩天牛(Apriona swainsoni(Hope))在木质部的蛀道多呈扭曲“Z”形,但在韧皮部的蛀道受寄主植物的生长状况和钻蛀位置影响而呈多种类型[14]。本研究中调查发现松褐天牛在黑松和赤松上的蛀食特征无明显差异,且蛀道类型较统一,均为“U”形,表明蛀道特征与寄主树种的相关性也不大。大多数植食性的昆虫都受寄主植物生理状态和营养状况的影响,昆虫取食营养构成不同的寄主植物时,其体内的消化分解系统也会随之发生变化,影响昆虫对寄主植物的适应状况[21]。本研究中松褐天牛在黑松上的蛀食特征比赤松上的稍明显些,也间接表明树种间生理状况和营养水平的差异,即松褐天牛一定程度上更偏好蛀食黑松,这与郝德君等[11]研究中松褐天牛成虫在产卵选择中更偏好黑松的结果是一致的。但产卵偏好和蛀食偏好并不完全代表松褐天牛对黑松的危害程度高于赤松,因为松褐天牛幼虫在蛀食过程中并未将蛀道内的蛀屑全部取食,大部分蛀屑被推出坑道外,而且松褐天牛幼虫个体大小和蛀食特征相关性不大。这与张慧杰等[22]报道的美洲斑潜蝇(Liriomyza sativaeBlanchard)在不同寄主上的虫道面积和蛹质量之间相关程度极其微弱相一致。
另外,钻蛀类昆虫的取食经历和生境异质性可能对其成虫补充营养行为及种群遗传分化产生显著影响[23]。而松褐天牛幼虫在其它树种上的蛀食特征和成熟期阶段的行为等是否存在差异,仍需进一步研究。也有研究表明钻蛀性昆虫的蛀道内节肢动物种群极为丰富[24],因此,仍需期望对松褐天牛蛀道结构的功能及其相同生态位中的伴生生物进行深入研究,以期为探索松褐天牛的精准防控提供依据和新思路。
4 结论
松褐天牛在赤松和黑松内的蛀道均为“U”形,类型单一,蛀道间互不串通,在侵入孔端部存在蛀屑堵塞,多数羽化孔在侵入孔上方;松褐天牛的蛀食特征与幼虫个体大小和寄主树种的相关性均较小。
猜你喜欢
湖南林业科技(2023年5期)2023-10-31 08:29:24
种子(2023年5期)2023-08-01 06:54:36
——以蛀食类害虫为例
高教学刊(2022年30期)2022-10-30 02:02:48
南方林业科学(2021年3期)2021-07-28 07:51:34
汉语世界(2017年5期)2017-09-21 07:44:37
山东林业科技(2016年5期)2016-07-05 00:43:04
意林(2015年1期)2015-05-14 16:49:40
花木盆景·盆景赏石(2010年7期)2010-07-28 05:39:30
花木盆景·盆景赏石(2010年6期)2010-06-19 05:53:06
花木盆景·盆景赏石(2010年5期)2010-05-18 04:42:44
