
石灰岩山地不同生境淡竹的养分特征及其生理适应
2022-03-25 08:47:06吴姝瑾胡姝珍陈永镇俞社保施建敏
林业科学研究 2022年2期
吴姝瑾,梁 宽,樊 燕,苏 田,胡姝珍,陈永镇,俞社保,施建敏*
(1.江西农业大学林学院,江西 南昌 330045;2.江西省竹子种质资源与利用重点实验室,江西 南昌 330045)
氮(N)、磷(P)是植物体内维持细胞结构和功能的重要生命元素,是限制陆生植物生产力的两个关键因子[1]。植物的N、P 来源于土壤,在养分缺乏的生境中,植物对养分的吸收和合理分配是适应生境的重要方式[2-3]。石灰岩生境是典型的喀斯特系统,其土壤浅薄、异质性高,土壤养分供应不均衡[4]。石灰岩生境土壤有效氮含量在0.21 mg·g-1左右,有效磷含量为0.003~0.005 mg·g-1(低于0.01 mg·g-1缺磷标准)[5]。研究认为,石灰岩土壤缺磷,限制植物生长的养分通常是磷而不是氮[6-8]。
近年来,研究者对喀斯特生境的异质性、土壤养分供应、植物养分特征进行了研究。高华端[9]以裸岩率的高低将石灰岩生境分为三类:连续土(CS),裸露率<30%;半连续土(SCS),裸露率30%~50%;零星土(SS),裸露率>50%。吴海勇等[10]在桂西北峰丛洼地区研究发现,在高裸岩率区域内,土壤N 含量增加;张忠华等[11]、李恩香等[12]研究岩溶植被均发现,随着裸岩率的增高,土壤中的全P 含量随之降低。还有研究对喀斯特植物养分进行了分析,张亚冰等[13]发现,同一种植物在喀斯特地区生境下叶片的N 含量比在非喀斯特生境下高,随着裸岩率上升植物叶片的N 含量呈上升趋势;周俊妞等[14]在桂林喀斯特岩溶石山区域对50 种植物叶片养分特征进行探究,发现在喀斯特地区植物叶片中的N 含量和P 含量呈显著的正相关;原雅楠等[15]发现,榧树(Torreya grandisFort.et Lindl.)不同器官N、P 变化趋势一致且叶片中N、P 含量最高;庞丽等[16]发现,随裸岩率上升蕨类植物对土壤中的P 利用率提高。然而,在土壤养分供应迥异(N 充裕而P 缺乏)的喀斯特生境,植物如何进行体内养分分配调节尚不明确,含量丰富和缺乏的元素适应特征是否一致亦不清楚。
在赣西北石灰岩山地有大面积淡竹(Phyllostachys glaucaMcClure)纯林成片分布,甚至在裸岩率高达60%的生境下仍能生长[17]。由此可见,淡竹对石灰岩生境具有很强的适应能力,是研究植物对石灰岩生境养分适应的理想材料。为此,本文依据高华端[9]的生境分类,研究不同石灰岩生境N、P 养分供应特征,比较分析石灰岩优势种淡竹在低P 高N 生境中的体内养分分配特征与差异,以期探析淡竹在石灰岩生境养分适应机制,为喀斯特植被管理和植物资源利用提供理论依据。
1 研究区概况
瑞昌市位于江西省西北部(29°23'~29°51' N,115°06'~115°44' E),北临长江,幕阜山余脉自西向东绵延全境,地形多为低山、丘陵。气候属中亚热带北缘湿润性季风气候,年均气温16.6 ℃,极端最高气温41.2 ℃,极端最低气温-13.4 ℃,年均降水量1 394 mm,年均日照时数1 890 h,无霜期260 d[18]。研究区有5 600 hm2天然淡竹成片分布于石灰岩山地,其它树种仅零星点缀其中[19-20]。
2 研究方法
2.1 试验设计及取样方法
于2014 年7 月连续晴天,选择海拔、坡向一致的零星土、半连续土、连续土3 种生境,每生境3 次重复。每类生境按年龄分层法取9 株淡竹,同时在每株取样淡竹附近用土钻随机取3 份土壤样品作为混合样,取样深度为0~30 cm。淡竹采用全株收获法并分为地上部叶(Le)、枝(Br)、秆(Cu)和地下部蔸(Su)、鞭(Rh)、根(Ro)6个构件样品进行烘干处理,测定N、P 含量。地下部构件取样标准为:蔸为从竹秆颈部至与竹鞭相连的部分;鞭以待测分株为中心,通过挖掘确认与竹鞭相连的左右分株位置后,截取来鞭和去鞭各一半;根包括蔸根和鞭根两部分。
2.2 指标测定
称取烘干样品(土样250 mg,植物样品150 mg),加1.5 g 催化剂(硫酸钾:硫酸铜=10:1)后再加5 mL 浓硫酸,消煮完成后移入100 mL容量瓶定容,摇匀静置。
2.2.1 氮测定 取1 mL 消煮液于50 mL 容量瓶,加1 mL EDTA-甲基红,加入适量水,用酸碱调节pH,使溶液由红色变为淡黄色,再加5 mL 酚和5 mL 次氯酸钠摇匀,定容1 h 后在625 nm 比色。
全株N 含量=(叶N 含量 × 叶生物量+枝N含量 × 枝生物量+秆N 含量 × 秆生物量+蔸N 含量 × 蔸生物量+鞭N 含量 × 鞭生物量+根N 含量 ×根生物量)/全株生物量
叶片N 含量占比=(叶生物量 × 叶N 含量)/(全株N 含量 × 全株生物量)× 100%
其他构件(枝、秆、蔸、鞭、根)N 含量占比计算方式与叶相同。
2)重视施工材料的质量,保证各项材料的质量检验证明材料的齐全性。对于施工中所涉及的材料,在使用之前对施工材料的质量进行保证,就可以通过对材料的相关证明进行准备。
2.2.2 磷测定 取10 mL 消煮液于50 mL 容量瓶,加1 滴2,4-二硝基酚,加入适量水,用酸碱调节pH,使溶液由无色变成淡黄色,再加5 mL钼锑抗试剂,摇匀,定容30 min 后在880 nm 比色。
全株P 含量和构件P 含量占比计算与上述N 的计算方式一致。
2.3 生物吸收系数计算
生物吸收系数(A)表示植物从环境中吸收和积累化学元素的能力,淡竹N、P 生物吸收系数计算公式为[21]:

式中:A为生物吸收系数,Cp为植物体内某元素含量,Cn为土层元素含量。
3 研究结果
3.1 3 种生境土壤氮、磷含量
在3 种不同生境下,随着裸岩率的增加,土壤的全N 含量逐渐增加(图1)。零星土生境全N 含量与连续土、半连续土生境间差异显著(p<0.05),呈现零星土(3.08±0.29 mg·g-1)>半连续土(2.07±0.18 mg·g-1)>连续土(1.73±0.06 mg·g-1)的趋势;P 含量在连续土、零星土生境与半连续土生境间差异显著(p< 0.05),呈现连续土(0.382±0.03 mg·g-1)>零星土(0.381±0.02 mg·g-1)>半连续土(0.29±0.01 mg·g-1)的趋势(图1)。
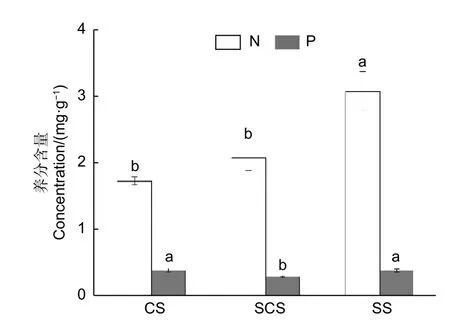
图1 石灰岩山地淡竹林3 种生境土壤的全氮、全磷含量Fig.1 Concentrations of soil nitrogen and phosphorus in three habitats in limestone mountains
3.2 不同生境植株水平氮、磷含量及氮磷比
在3 种不同生境,淡竹N、P 含量及N∶P 呈现出不同的规律(图2a),零星土生境下植株全N 含量显著(p<0.05)高于连续土和半连续土,连续土生境下植株P 含量显著(p<0.05)高于半连续土和零星土,由此N:P 呈现出随裸岩率增大而增大的趋势,3 种生境下N:P 均差异显著(p<0.05)。
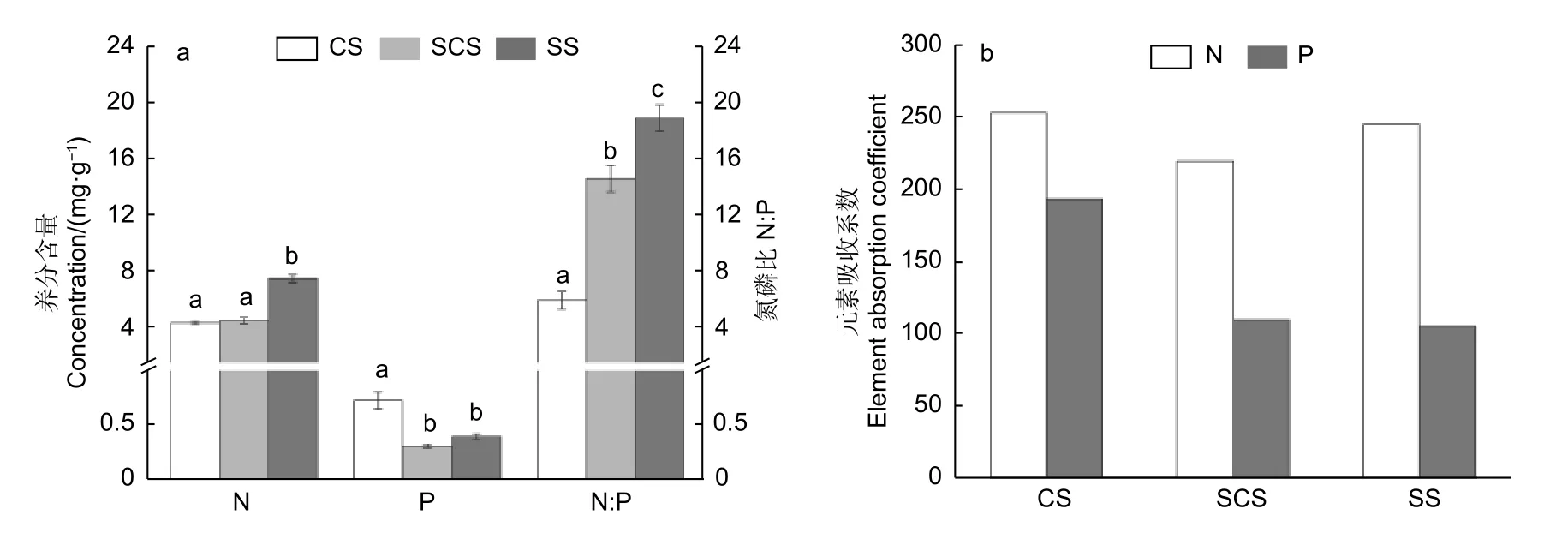
图2 不同生境淡竹全株氮、磷含量及N:P(a)和氮磷吸收系数(b)Fig.2 Concentrations of nitrogen, phosphorus and N: P (a), and N and P absorption coefficients (b) of Phyllostachys glauca in three habitats
淡竹对N 元素的吸收系数在连续土、半连续土、零星土分别为251.67、217.97、243.70;对P 元素的吸收系数随着裸岩率的增加不断减小,连续土、半连续土、零星土的数值分别为191.83、108.85、104.38(图2b)。在3 种生境下,N 和P 的吸收系数均大于100,即2 种元素都是在淡竹体内累积,但随着裸岩率的增加,淡竹对P 元素的吸收能力逐渐减弱。
3.3 不同生境淡竹构件水平氮、磷含量及氮磷比
3.3.1 N 含量变化 图3a 表明:在3 种生境中,淡竹不同构件N 含量的变化趋势为:除鞭外,叶、枝、秆、蔸、根的N 含量变化趋势均服从随裸岩率升高而升高,其中,叶的N 含量最高,在连续土、半连续土、零星土生境分别为17.36±0.60、19.49±0.51、20.41±0.37 mg·g-1;秆的N 含量在3 种生境下均最低,连续土、半连续土、零星土分别为2.36±0.12、2.02±0.20、3.11±0.36 mg·g-1。叶片和鞭中的N 含量在连续土与半连续土间差异显著(p<0.05)。
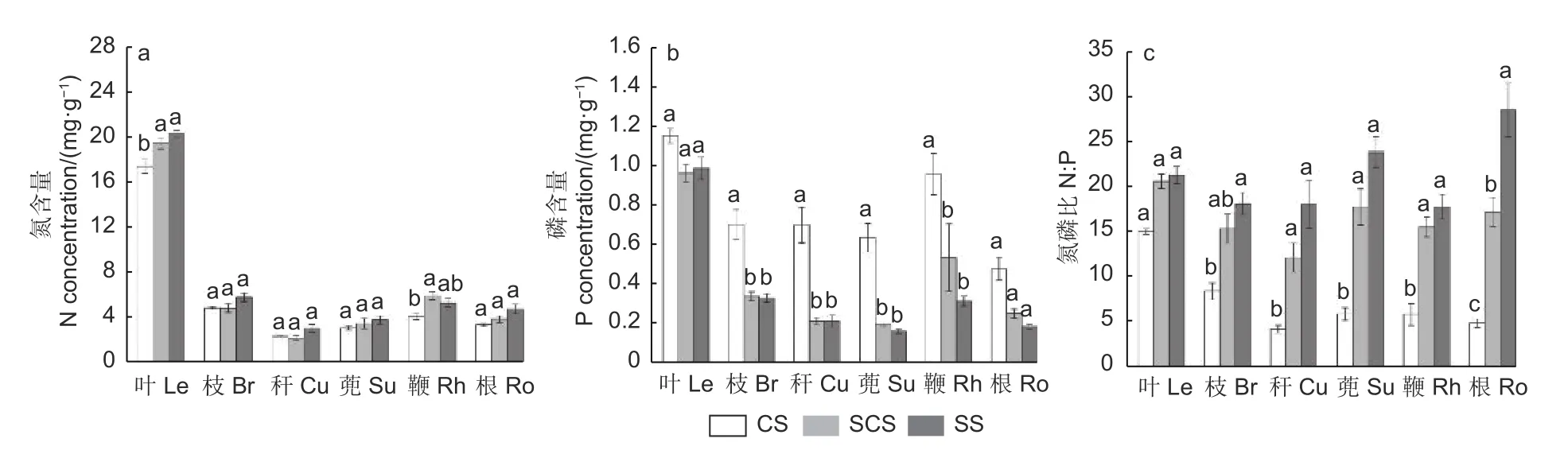
图3 不同生境下淡竹各构件N、P 含量及其N:PFig.3 N and P concentrations and N:P of Phyllostachys glauca in three habitats
3.3.3 N:P 变化 图3c 表明:随裸岩率升高,淡竹各构件的N:P 逐步升高。在连续土生境下的秆、蔸、鞭的N:P 与零星土生境之间显著差异(p<0.05);根的N:P 在3 种生境之间均存在显著差异(p<0.05)。所有构件中只有叶片在3 种生境下N:P 保持相对稳定,且3 种生境下N:P 差异不显著(p>0.05)。
3.4 不同生境淡竹构件N、P 分配格局
N、P 含量在植物体内的不同构件中分配占比不同。N 在叶中占比最大,其次为秆、枝、根、鞭、蔸(图4a);P 在叶片和秆中的占比较大,其次为枝、鞭、根、蔸(图4b)。N、P 随生境变化在各构件中的分配策略也不同。随裸岩率的升高,N含量在叶中的占比分别为49.55%、51.71%、48.28%,变化幅度较小,而在秆、鞭、根中的占比呈上升趋势。随裸岩率的升高,P 含量在叶片中的占比呈上升趋势,从连续土到零星土分配比例从25.81%上升至44.26%,差异显著(p<0.05);同时,秆中的P 占比从连续土到半连续土由40.48%降至26.74%,差异显著(p<0.05);枝、蔸、鞭、根中的P 占比亦有不同程度降低。

图4 不同生境淡竹构件N(a)、P(b)分配格局Fig.4 Distribution patterns of nitrogen(a) and phosphorus(b) of Phyllostachys glauca in three habitats
4 讨论
4.1 淡竹主要养分含量与分配特征
在赣西北地区的石灰岩山地上,淡竹林大面积广泛分布[19,22]。从连续土到零星土,随着裸岩率上升,土壤N 含量升高,淡竹全株的N 含量增加。在N 供应充足的情况下,植株保证叶片中的N 含量,然后将多余的N 分配给秆等其他构件。相比之下,土壤P 含量与N 截然相反,3 种生境的P 含量均较低。植物P 主要来源于土壤[9,23],土壤缺磷使淡竹植株P 含量下降,且各构件的P 含量均呈不同程度的下降(图3),其中,叶和根的P 含量下降较平缓,保持在一个相对稳定的值。
研究表明,在植物的各构件中,叶片的N、P 含量均高于其他构件[24-25]。李祥栋等[26]在薏苡抽穗期探究其养分分配,发现整株中叶片的N 占比最高,所占比例为46.59%~48.75%,且地上部分N 的积累量占整株的91.36%~91.12%。于钦民等[27]对杉木的养分分配开展研究,发现地上部分N 积累量最高可达80%,其中,叶的N 积累量为全株40%~60%,P 在叶片中积累量同样最高,占35%~45%。与其他植物的养分分配类似,淡竹的N、P 主要分配在地上部分的叶、枝、秆中,而地下部分的比例较小,不超过16.35%;随着生境裸岩率升高,N 供应充足而P 供应缺乏,淡竹叶片的N分配比例变化较小,叶片P 的分配比例大幅增加,枝、秆等其余构件中P 的分配比例均下降(图4)。这表明植物在P 缺乏生境下,有限的P 在植物体内进行重新分配,枝、秆中的P 被转移出来分配到更关键的构件上。王雪梅等[28]研究表明,在养分匮乏的情况下,车桑子(Dodonaea viscosa(L.)Jacq.)把更多的养分集中分配给叶片,保证叶片光合需要以维持植物生长,提高逆境下的适合度。该观点在本研究中得以印证,零星土生境淡竹降低秆中的P 分配比例而提高叶片中的分配量,以适应土壤的低P 供应。
陆地生态系统中,N、P 的有效性通常限制了植物生长,N:P 是表征群落养分限制的重要指标[29]。研究表明,N:P<14,植物受N 限制;N:P>16,植物受P 限制;介于二者之间时,则植物受N 或P 或N:P 影响[30]。通过植物各构件的化学计量特征,可以探究不同生境下植物内稳态特征[31]、植物特性[32]、物种组成以及多样性[33]。本研究中,随生境裸岩率的增加,淡竹植株N:P 不断上升,表现为连续土(5.93±0.16)<半连续土(14.52±0.40)<零星土(18.83 ±0.48),表明受P 限制程度越来越高;同时,叶片N∶P 却没有因为土壤缺磷程度的上升而发生急剧变化,从连续土到零星土的N:P 保持相对稳定,表现为:连续土(15.12±0.32)<半连续土(20.70±0.80)<零星土(21.41±0.97)。可见,淡竹在受P 限制的情况下,通过调节构件的分配满足叶片的N、P 含量稳定以保证正常生长需求。
4.2 淡竹N、P 分配格局比较
在3 种生境下,淡竹N、P 在植物体内的分配格局存在差异。从连续土到零星土,土壤中的N 含量随裸岩率上升而上升,但土壤可利用P 始终保持在较低水平。吴海勇等[10]研究发现,喀斯特高裸岩率区域土壤面积有限,凋落物高度聚集,造成了土壤中的N 含量增高,这与本文的研究结果一致。姜沛沛等[34]研究发现,地表土壤对P 具有吸附作用,伴随着水土流失造成土壤P 含量下降。另外,喀斯特地区土壤中P 多以钙磷(Ca-P)的形式被固定,能被植物直接利用的P 含量约1~8 mg·kg-1[35-36]。对于石灰岩生境土壤N 充足而P 缺乏的特点[37],淡竹采用了不同的分配格局策略以适应环境:从连续土到零星土,各构件的N 分配比例相对稳定,不同生境间差异不显著(图4a);P 的分配格局则不同,其在秆中的分配占比从连续土生境的40.48%下降到半连续土、零星土的不及30%,而在叶片中的分配占比从25.81%上升40%以上(图4b)。这表明,淡竹将更多的P 从秆中转移分配至叶片中,以保证叶片对P 的需求,以缓解P 缺乏对植物的影响。
叶是植物新陈代谢最旺盛的部位,光合作用需要多种光合酶的参与,而叶片光合速率与叶氮含量有密切的关系[38]。在光合途径中更高效的利用N,是抵御缺P 有害影响的一种机制[39]。郭延平等[40]研究发现,叶片在缺P 条件下光合作用受到光抑制。本研究中,随着裸岩率的增加,淡竹增加其叶片中的N 含量,旺盛的光合作用促进叶片中C 的积累使其快速生长繁殖[41];而当土壤P 缺乏时,淡竹通过合理调节分配更多P 至叶片来满足生理需求,以适应生境的养分变化。张天霖等[42]在粤北地区开展植物器官养分研究后认为,在养分缺乏的生境下,优势物种通过复杂的内在协调机制来维持其在群落中的优势地位。课题组前期对不同生境淡竹的生长研究发现,从连续土到零星土生境,淡竹平均胸径分别为2.47、2.63、3.01 cm,株高分别为5.64、5.95、6.67 m,胸径和株高均在零星土生境下达到最大,即零星土生境下淡竹长势好,且生物量高[43]。本研究发现,淡竹在零星土生境会调节N、P 养分分配以保证叶片生理需求,这一行为可能是支撑淡竹在P 缺乏生境应对胁迫保持良好长势的一个重要生理机制。
5 结论
养分的吸收和合理分配是植物应对养分匮乏生境的重要适应方式。本研究对赣西北石灰岩山地连续土、半连续土和零星土3 种土被生境优势种淡竹进行调查,研究发现植株N、P 含量与土壤N、P 供应水平正相关,在土壤N 供应充足而P 供应匮乏的情况下,淡竹通过减少秆中的P 分配而提高叶片的分配量,以保证叶的养分需求维持整株的正常生长,提高逆境下的适合度。研究结果揭示了淡竹在缺磷生境下的生存策略,为喀斯特植被管理和植物资源利用提供了理论参考。
猜你喜欢
青年文学家(2022年1期)2022-03-11 12:27:39
江苏安全生产(2021年11期)2022-01-19 02:23:22
落叶果树(2021年6期)2021-02-12 01:29:10
落叶果树(2021年6期)2021-02-12 01:28:54
文苑(2020年12期)2020-04-13 00:54:10
海峡姐妹(2019年1期)2019-03-23 02:42:46
江西建材(2018年2期)2018-04-14 08:01:03
体育科技(2016年2期)2016-02-28 17:06:07
外语教学理论与实践(2015年1期)2015-06-11 02:51:12
西华师范大学学报(自然科学版)(2015年3期)2015-02-27 15:31:17
